


Abstract
The concept of a purinergic signaling system, using purine nucleotides and nucleosides as extracellular messengers, was first proposed over 30 years ago. After a brief introduction and update of purinoceptor subtypes, this article focuses on the diverse pathophysiological roles of purines and pyrimidines as signaling molecules. These molecules mediate short-term (acute) signaling functions in neurotransmission, mechanosensory transduction, secretion and vasodilatation, and long-term (chronic) signaling functions in cell proliferation, differentiation, and death involved in development and regeneration. Plasticity of purinoceptor expression in pathological conditions is frequently observed, including an increase in the purinergic component of autonomic cotransmission. Recent advances in therapies using purinergic-related drugs in a wide range of pathological conditions will be addressed with speculation on future developments in the field.
I. Introduction
A seminal article by Drury and Szent-Györgi in 1929 described the potent actions of purine nucleotides and nucleosides, ATP, and adenosine on the heart and blood vessels. Then, in 1970, evidence was presented for ATP as a neurotransmitter in nonadrenergic, noncholinergic (NANC1) nerves supplying the gut (Burnstock et al., 1970) and in 1972 the word “purinergic” was coined and the purinergic neurotransmission hypothesis was proposed by Burnstock (Burnstock, 1972). This concept met with considerable resistance for many years, because ATP had been established as an intracellular energy source involved in various metabolic cycles, and it was thought that such a ubiquitous molecule was unlikely to be involved in selective extracellular signaling. However, the concept is now widely accepted. Later, it was established that ATP was a cotransmitter with classic transmitters in both the peripheral and central nervous systems and that purines are also powerful extracellular messengers to non-neuronal cells, including exocrine and endocrine, secretory, endothelial, musculoskeletal, immune, and inflammatory cells (Burnstock and Knight, 2004).
Implicit in the purinergic hypothesis was the presence of purinoceptors (Ralevic and Burnstock, 1998). A basis for distinguishing P1 (adenosine) from P2 (ATP/ADP) receptors was proposed by Burnstock in 1978. This helped resolve some of the ambiguities in earlier reports, which were complicated by the breakdown of ATP to adenosine by ectoenzymes so that some of the actions of ATP were directly on P2 receptors, whereas others were due to indirect action via P1 receptors. Four subtypes of P1 receptors were cloned, namely, A1, A2A, A2B, and A3. In 1985, Burnstock and Kennedy proposed a basis for distinguishing two types of P2 purinoceptor, namely, P2X and P2Y, based largely on pharmacological criteria. In the early 1990s, studies of transduction mechanisms and cloning of both P2X and P2Y receptors were carried out, which led Abbracchio and Burnstock to put forward a new nomenclature system in 1994, which is now widely accepted (Fredholm et al., 1994; Schwiebert, 2003). They proposed that there were two families of P2 receptors, namely, P2X ionotropic ligand-gated ion channel receptors and P2Y metabotropic G protein-coupled receptors. This framework has allowed a logical expansion as new receptors were identified. Currently seven subtypes of P2X receptors and eight subtypes of P2Y receptors are clearly established (Table 1). It is recognized that the pyrimidine UTP is equipotent with ATP at P2Y2 and P2Y4 receptors and UDP is a potent agonist at P2Y6 receptors. The distribution and functional roles of P2 receptors in many cell types in the body have been described (Burnstock and Knight, 2004).
Characteristics of purine-mediated receptors Table modified from Burnstock (2003a), with permission from Elsevier (New York, NY).
Purinergic signaling is rapid in synaptic neurotransmission, in neuromuscular transmission leading to contraction or relaxation of smooth muscle, and in exocrine or endocrine secretion. However, there are now many examples of purinergic signaling regulating long-term events such as cell proliferation, differentiation, migration, and death in development, regeneration, and wound healing. Both P2X and P2Y receptors play prominent roles both directly and by modulation of other signaling systems in embryonic development, including the nervous system, cartilage in limb buds, the mesonephros, retina, myotubes, and neuromuscular junctions (Burnstock, 2001a).
There is increasing interest in the therapeutic potential of purinergic compounds (including receptor agonists and antagonists, ectoenzyme inhibitors and enhancers, and ATP transport inhibitors and enhancers) in a wide range of disease conditions in relation to both P1 and P2 receptors. A number of purine-related compounds have been patented. The autonomic nervous system shows marked plasticity. Dramatic changes occur in the expression of cotransmitters and receptors during development and aging, in nerves that remain after trauma or surgery and in disease conditions. There are several pathological conditions in which the purinergic component of cotransmission is increased (Burnstock, 2002b). The emphasis in this article is on the pathophysiology and therapeutic potential of P2 receptors (see also Burnstock and Williams, 2000; Boeynaems et al., 2001; Yerxa, 2001; Burnstock, 2002a; Jacobson et al., 2002; Ralevic and Burnstock, 2003) and readers are referred to recent reviews about the therapeutic potential of P1 receptors (Fredholm et al., 2002, 2005; Okusa, 2002; Pelleg et al., 2002; Dhalla et al., 2003; Ribeiro et al., 2003; Hutchinson and Scammells, 2004; McCallion et al., 2004).
II. Cardiovascular Diseases
ATP and adenosine are very much involved in the mechanisms underlying local control of vessel tone (Burnstock and Ralevic, 1994) (Fig. 1) as well as cell migration, proliferation, differentiation, and death during angiogenesis, atherosclerosis, and restenosis following angioplasty (Erlinge et al., 1998; Burnstock, 2002b). ATP, released as a cotransmitter from sympathetic nerves, constricts vascular smooth muscle via P2X receptors, whereas ATP released from sensory-motor nerves during “axon reflex” activity dilates or constricts some vessels. Furthermore, ATP released from endothelial cells during changes in flow (shear stress) or hypoxia acts on P2Y receptors in endothelial cells to release nitric oxide (NO), resulting in relaxation. Adenosine produced by the breakdown of extracellular ATP causes vasodilatation via smooth muscle P1 receptors. P2X receptors are also present on endothelial cells and appear to be associated with cell adhesion and permeability. ATP release from red blood cells is increased in pathological conditions such as subarachnoid hemorrhage, largely because there is widespread blood cell lysis (Sprague et al., 2003). This leads to transient constriction of arterioles via P2X receptors and sustained constriction of large cerebral vessels, largely through P2Y2 receptors. The differences in purinergic receptor distribution between macro- and microvessels in the cerebral circulation are likely to have important consequences in pathological conditions.
A. Thrombosis
There have been very promising developments concerning purinergic antithrombotic drugs. Platelets are known to express P2Y1, P2Y12, and P2X1 receptors (Hollopeter et al., 2001). “Mega” clinical trials CAPRIE (CAPRIE Steering Committee, 2001), CURE (Yusuf et al., 2001), and CREDO (Beinart et al., 2005) have provided clear evidence that the purinergic antithrombotic drugs clopidogrel and ticlopidine reduce the risks of recurrent strokes and heart attacks, especially when combined with aspirin (Kam and Nethery, 2003; Kunapuli et al., 2003; Boeynaems et al., 2005). These drugs are antagonists to the P2Y12 receptor that mediates platelet aggregation (Gachet, 2001). MRS2500, a highly potent and selective antagonist for the P2Y1 receptor, has also been shown recently to have antiaggregating activity on human platelets (Cattaneo et al., 2004). There are genetic variations in P2Y1 and P2Y12 receptor gene sequences in healthy subjects that explain variations in the platelet response to ADP; this may reflect individual variation in atherothrombotic risk and the efficacy of purinergic antithrombotic drugs (Fontana et al., 2003; Hetherington et al., 2004). A recent study has shown that there is a synergistic inhibition of ADP-induced platelet activation via P2Y1 and P2Y12 receptors and for thrombin and P2Y12 inhibition; it is suggested that there may be clinical benefit by combining these inhibitors, providing that bleeding problems do not outweigh this benefit (Nylander et al., 2004). Postoperative carotid thrombosis is a significant risk for stroke; it seems likely that clopidogrel or ticlopidine may provide an avenue for targeted antiplatelet therapy following vascular intervention (Hayes et al., 2003). Platelet aggregation in response to ADP is significantly inhibited in patients with peripheral vascular disease 2 to 4 h after a loading dose of clopidogrel, and ADP-induced platelet shape change is significantly inhibited following ingestion of clopidogrel (Matsagas et al., 2003). There is a synergistic interaction between ATP and noradrenaline in stimulating platelet aggregation, which suggests a prothrombotic role for ATP in stress (Birk et al., 2003). Platelet activation that occurs in human acute malaria infection is associated with elevated plasma ATP concentrations (Essien and White, 1998).
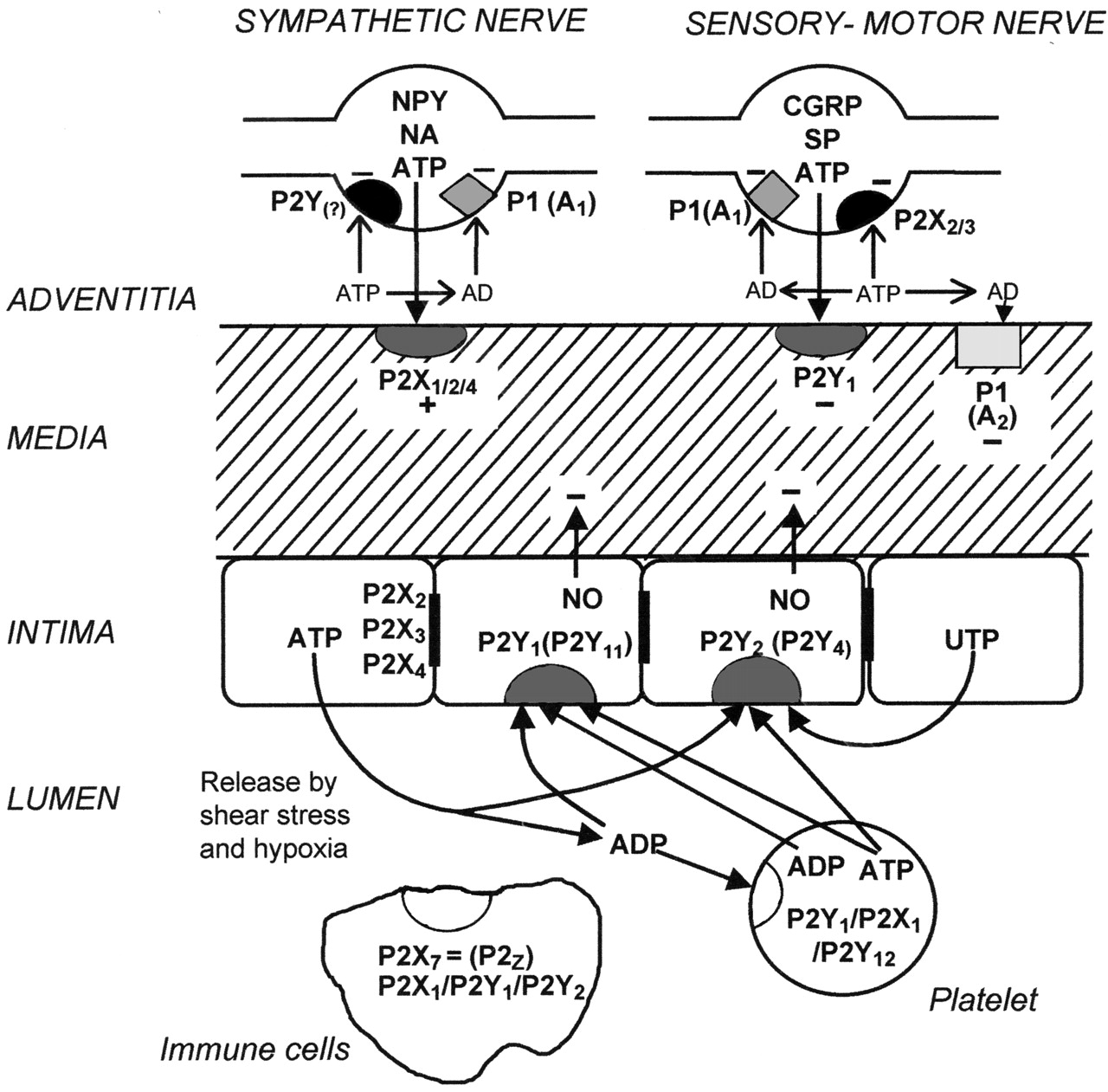
Short-term (acute) purinergic signaling controlling vascular tone. Schematic diagram illustrating the main receptor subtypes for purine and pyrimidines present in most blood vessels. Perivascular nerves in the adventitia release ATP as a cotransmitter: ATP is released with NA and neuropeptide Y (NPY) from sympathetic nerves to act on smooth muscle P2X1 and in some vessels P2X2, P2X4, and P2Y2 purinoceptors, resulting in vasoconstriction; it is released with calcitonin generelated peptide (CGRP) and substance P (SP) from sensory nerves during “axon reflex” activity to act on smooth muscle P2Y-purinoceptors in some regions of some vessels, resulting in vasodilatation. P1(A1)-purinoceptors on nerve terminals of sympathetic and sensory nerves mediate adenosine (arising from enzymatic breakdown of ATP) modulation of transmitter release. P2X3 purinoceptors are present on a subpopulation of sensory nerve terminals. P1(A2)-purinoceptors on vascular smooth muscle mediate vasodilatation. Endothelial cells release ATP and UTP during shear stress and hypoxia to act on P2Y1, P2Y2, and sometimes P2Y4 purinoceptors, leading to the production of NO and subsequent vasodilatation. ATP, following its release from aggregating platelets, also acts on these endothelial receptors. Blood-borne platelets possess P2Y1 and P2Y12 ADP-selective purinoceptors as well as P2X1 receptors, whereas immune cells of various kinds possess P2X7, as well as P2X1, P2Y1, and P2X2 purinoceptors. P2X2, P2X3, and P2X4 receptors have also been identified on endothelial cell membranes. Reproduced from Burnstock (2002a), with permission from Lippincott Williams & Wilkins (Philadelphia, PA).
Much is now known about the ectonucleotidases that break down ATP released from non-neural cells as well as neurons (Zimmermann, 2001; Vorhoff et al., 2005). Several enzyme families are involved: ectonucleoside triphosphate diphosphohydrolases (E-NTPDases), of which NTPDase1, 2, 3, and 8 are extracellular; ectonucleotide pyrophosphatase of three subtypes; alkaline phosphatases, ecto-5′-nucleotidase and ectonucleoside diphosphokinase. NTPDase1 hydrolyzes ATP directly to AMP and UTP to UDP, whereas NTPDase2 hydrolyzes ATP to ADP and 5′-nucleotidase AMP to adenosine. Ectonucleotidases are expressed by vascular endothelium, accessory vascular cells (e.g., monocytes, pericytes, and vascular smooth muscle cells), and dendritic cells; these are predominantly NTPDase1 and NTPDase2 (Sevigny et al., 2002). NTPDase1 hydrolyzes both tri- and diphosphonucleosides and blocks platelet aggregation responses to ADP. In contrast, NTPDase2, a preferential nucleoside triphosphatase, activates platelets by converting the competitive antagonist (ATP) of platelet ADP receptors to the specific agonist. Vascular NTPDase1 biochemical activity is rapidly lost from the endothelium of vascularized cardiac grafts subjected to oxidant stress. These changes are associated with thrombotic injury and platelet sequestration at sites of injury.
B. Heart Failure
Up-regulation of P2X1 and P2Y2 receptor mRNA in the hearts of rats with congestive heart failure has been reported (Hou et al., 1999) and an increase in expression of P2X1 receptors in the atria of patients suffering from dilated cardiomyopathy (Berry et al., 1999). In congestive heat failure, the muscle mechanoreflex is accentuated and appears to be associated with enhanced P2X receptor-mediated responses of sensory nerves to α,β-methylene ATP (Sinoway and Li, 2005). The positive inotropic response to ATP and ATP-induced increases in [Ca2+]i in cardiomyocytes are impaired in heart failure due to myocardial infarction; imidapril partially reverses this impairment (Saini et al., 2005). ATP and adenosine are widely used for the treatment of paroxysmal supraventricular tachycardia in both infants and adults (Belhassen and Pelleg, 1985; Vassort, 2001). The substantial enhancement of mechanical performance with 2-deoxy-ATP in cardiac muscle suggests that it may be a better substrate for contractility than ATP and suggests that ribonucleotide reductase may be a target for therapy in heart failure (Regnier et al., 2000).
Enhanced sympathetic nerve activity causes cardiac dysfunction, arrhythmias, and sudden cardiac death in myocardial ischemia. ATP is coreleased with noradrenaline (NA) and enhances NA release from sympathetic nerve terminals. A role for the ectonucleotidase, E-NTPDase1 at sympathetic nerve terminals may offer a novel therapeutic approach to hyperadrenergic states such as myocardial ischemia (Sesti et al., 2002). Because NTPDase1 also inhibits platelet aggregation, it is proposed for the treatment of thrombotic diatheses (Marcus et al., 2003). To examine the role of NTPDases after cardiac transplantation and grafts, several techniques to manipulate NTPDase1 expression have been used, for example, deletion of this enzyme in mice. These mutant mice exhibit major perturbations of P2 receptor-mediated signaling in the vasculature and immune systems and show hemostatic defects, thromboregulatory disturbances, and heightened acute inflammatory responses with a failure to generate cellular responses (Enjyoji et al., 1999); NTPDase1-null hearts have been shown to develop diffuse thrombotic infarcts and undergo rejection more rapidly than control grafts when transplanted into wild-type mice or into rats. In contrast, transgenic organs or cardiac grafts in which NTPDase1 has been up-regulated by gene therapy vectors enjoy a survival advantage post-transplantation (Imai et al., 2000).
ATP is routinely used to terminate spontaneous ventricular tachycardia, but it is not always successful, although no difference in efficacy was found between ischemic and nonischemic cardiomyopathy (Kouakam et al., 2003). By using transgenic overexpression of human P2X4 receptors, cardiac P2X4 receptors have been shown to have a beneficial life-prolonging role in heart failure; increased expression or activation of these ATP-stimulated receptor channels may represent a new therapeutic approach to the treatment of heart failure (Yang et al., 2004). Administration of ATP during sinus rhythm has been suggested as a useful bedside test for identifying patients with a concealed accessory pathway who are prone to develop atrioventricular reentrant tachycardia (Belhassen et al., 2000). The negative chronotropic effect of ATP on sinus node is much more pronounced in patients with sick sinus syndrome (Tan et al., 2004).
Magnesium-ATP has been recommended for the treatment of ischemia, radiation injury, shock, and sepsis for many years (Harkema and Chaudry, 1992; Nalos et al., 2003). A recent study suggests that the protective effect of ATP on renal ischemic injury is, in part, related to inhibition of the inducible transcription factor, nuclear factor-κβ, via P2Y receptors in proximal tubule cells (Lee and Han, 2005). ATP was shown to inhibit atrioventricular conduction rather than the firing rate of sinoatrial nodes in patients with ischemic heart disease; injection of ATP is used only when a transient cardiac standstill is needed, such as for endovascular grafting surgery (Watanabe et al., 2002). Treatment of myocardial ischemia and reperfusion by ATP-MgCl2 is still recommended (Katircioglu et al., 2000), but many reports suggest that adenosine (a breakdown product of ATP) also mediates this effect (Liang and Jacobson, 1999; Broadley, 2000; Sommerschild and Kirkeboen, 2000; Thompson et al., 2002).
C. Hypertension
ATP plays a significant cotransmitter role in sympathetic nerves supplying hypertensive blood vessels. The purinergic component is increased in spontaneously hypertensive rats (Ralevic and Burnstock, 1998). The increase in sympathetic nerve activity in hypertension is well established, and there is an associated hyperplasia and hypertrophy of arterial walls (Julius and Nesbitt, 1996). ATP is a rapidly acting hypotensive agent that compares favorably with sodium nitroprusside (Kien et al., 1987). ATP-MgCl2 is a safe, effective, and preferential pulmonary vasodilator in children with pulmonary hypertension secondary to congenital heart defects; it has also been used for treating pulmonary hypertension after cardiac surgery (Brook et al., 1994). Red blood cells of humans with primary pulmonary hypertension fail to release ATP in response to mechanical deformation (Sprague et al., 2001). This is likely to result in the loss of local control of total pulmonary vascular resistance in the lung. Low-dose ATP exerts mitogenic actions on human pulmonary artery smooth muscle cells, which may be relevant to the pathophysiological basis of severe pulmonary hypertension (Zhang et al., 2004b). Pulmonary hypertension can be a problem in patients with chronic obstructive pulmonary disease, which also has other causes; it is a life-threatening condition, and intravenous ATP infusion produces a significant decrease in mean pulmonary arterial pressure and pulmonary vascular resistance without changing the mean systemic arterial pressure (Brook et al., 1994). ATP has been used to treat meconium aspiration-induced pulmonary hypertension in pigs (Kääpä et al., 1997). Eicosapentaenoic acid, one of the active components in fish oil that has antihypertensive effects, increases the release of ATP from vascular endothelial cells, leading to reduction of the blood pressure rise characteristic of aging (Hashimoto et al., 1998). It has been proposed that there is prognostic value of stress myocardial perfusion imaging using ATP at the beginning of hemodialysis treatment in patients with end-stage renal disease (Hase et al., 2004). P2X4 receptors are localized in the syncytiotrophoblast, stroma, and fetal capillary endothelial cells of human placenta. Placental P2X4 receptors are significantly up-regulated in mild preeclampsia (Roberts et al., 2005). The authors hypothesized that with preeclampsia, in which there is an increased potential for ATP release, P2X4 receptors may mediate a change in cellular homeostasis to limit cell damage.
D. Atherosclerosis and Restenosis
An early study reported that adenosine produces changes in cAMP and DNA synthesis in cultured arterial smooth muscle cells and suggested that this might result in the regulation of cell proliferation (Jonzon et al., 1985). The authors speculated that adenosine could be one of several regulatory factors in the development of atherosclerosis and might also regulate the release of a smooth muscle mitogen, platelet-derived growth factor. There is now good evidence that adenosine does regulate smooth muscle cell proliferation, but its properties differ from those for ATP and ADP (Burnstock, 2002a; Di Virgilio and Solini, 2002). Vascular injury represents a critical initiating event in the pathogenesis of various vascular diseases. Large amounts of ATP are released from injured cells and ATP and adenosine have potent actions in smooth muscle and endothelial cell growth, migration, proliferation, and death (Burnstock, 2002a). Apoptotic cell death is recognized to occur in a number of vascular diseases, including atherosclerosis and restenosis. Vascular endothelial cells are continuously exposed to variations in blood flow, which plays an important role in vessel growth or regression and in the local development of atherosclerosis. The shear stress that occurs during changes in blood flow leads to substantial release of ATP (and UTP) from endothelial cells (Burnstock, 1999), and these purines might mediate alterations in the balance between proliferation and apoptosis. Atherosclerotic damage results in the disappearance of endothelium-dependent responses to ATP. The release of ATP from endothelial cells has also been claimed to be impaired in atherosclerotic arteries and long-term supplementation with a high cholesterol diet decreases the release of ATP. In restenosis following balloon angioplasty, there is a peak in the proliferation and apoptosis of vascular smooth muscle cells at approximately 14 days. Clinical trials with clopidogrel and ticlopidine (P2Y12 receptor antagonists) in patients with atherosclerotic disease have shown significant benefit compared with aspirin.
Saphenous vein, internal mammary, and radial arteries have been used as grafts for coronary bypass surgery; the level of endothelial P2Y2 receptors is comparable in all three vessels, but endothelial P2X4 receptors vary from high in saphenous vein to significantly lower in the other two vessels. It has been suggested that P2X4 receptors play a more significant role in intense proliferation in arteriosclerosis and restenosis than P2Y2 receptors, as reflected by the susceptibility of saphenous vein grafts to atherosclerosis compared with internal mammary arteries (Ray et al., 2002). In another study, P2X1 and P2Y6 receptors mediated more prominent contractions in the saphenous vein compared with the internal mammary artery; it has been suggested that selective antagonists to these receptors may prevent vasospasm and restenosis in the saphenous vein during and after revascularization surgery (Borna et al., 2003). A novel role for P2Y2 receptors in the development of atherosclerosis has been suggested, whereby UTP induces vascular cell adhesion molecule-1 expression in coronary artery endothelial cells that mediate the recruitment of monocytes (Seye et al., 2003). The long-term (trophic) roles of purinergic signaling in vascular smooth muscle and endothelial cell proliferation and death have been implicated in atherosclerosis and restenosis and suggest the exploration of therapeutic strategies in relation to these events (Erlinge et al., 1998; Burnstock, 2002b; Hou et al., 2002; Wang et al., 2003b).
Newly developing vascular endothelia express very high levels of NTPDase1, also seen under hypoxic conditions (Eltzschig et al., 2003). Angiogenesis requires the dynamic interaction of endothelial cell proliferation and differentiation with orchestrated interactions between extracellular matrix and surrounding cells (such as vascular smooth muscle and/or pericytes). Such interactions could be coordinated by interplay between nucleotide release, P2 receptor modulation, and altered NTPDase expression (Goepfert et al., 2001). The role for adenosine in growth regulation in angiogenesis has been reviewed recently (Adair, 2005).
Varicose veins are characterized by hypoxia, inflammation, and cell death, all likely to lead to increases in extracellular ATP. P2X1 and P2X7 receptors are expressed by saphenous vein smooth muscle, and it has been suggested that the resultant P2X7 receptor activation contributes to the decrease in contractile myocytes and resulting disorganization of the vessel wall (Cario-Toumaniantz et al., 1998).
III. Neurology
Functional implications for purinergic signaling in the nervous system include neuron-glia interactions, differentiation of stem cells, diseases of the nervous system and responses to injury, neuroimmune and neurovascular interactions, sympathetic transmission and plasticity, glial cell differentiation, intercellular communication between glia, and myelination (Fields and Stevens, 2000; Burnstock, 2003b; Newman, 2003; Pascual and Haydon, 2003; Bowser and Khakh, 2004; Mulligan and MacVicar, 2004; Davalos et al., 2005; Köles et al., 2005).
ATP is a cotransmitter in many nerve types, probably reflecting the early evolutionary presence of purinergic signaling (Burnstock, 1996a, 2004). There is evidence for the action of ATP as a cotransmitter with NA and neuropeptide Y in sympathetic nerves, with acetylcholine and vasoactive intestinal peptide in some parasympathetic nerves, with NO and vasoactive intestinal peptide in enteric NANC inhibitory nerves, and with calcitonin gene-related peptide (CGRP) and substance P in sensory-motor nerves. There is also evidence for the cotransmission of ATP with GABA in retinal nerves, and with glutamate, 5-hydroxytryptamine (serotonin), NA, or dopamine in nerves in the brain (Burnstock, 2003b). In sympathetically innervated tissues, such as the vas deferens or blood vessels, ATP produces fast responses mediated by P2X receptors, followed by a slower component mediated by G protein-coupled α-adrenoceptors. Similarly, in the parasympathetic nerves supplying the urinary bladder, ATP provokes a fast transient response via P2X receptors, whereas the slower component is mediated by G protein-coupled muscarinic receptors. There are differences in the proportion of cotransmitters between species in development and aging and in different pathophysiological conditions. P2X3 and P2X2/3 receptors are selectively localized on sensory neurons in trigeminal, nodose, and dorsal root ganglia (DRG), and the terminals of these nociceptive neurons in the skin and visceral organs represent unique targets for novel analgesic agents that function as P2X3 and P2X2/3 receptor antagonists. Nonspecific P2 receptor antagonists, e.g., suramin and pyridoxal-5′-phosphate-6-azophenyl-2′,4′ disulfonic acid (PPADS), are antinociceptive, and P2X3 receptor-knockout mice reduce nociceptive inflammatory responses (Cockayne et al., 2000; Burnstock, 2001b).
The first clear evidence for nerve-nerve purinergic synaptic transmission was published in 1992 (Edwards et al., 1992; Evans et al., 1992; Silinsky et al., 1992). Synaptic potentials in the celiac ganglion and in the medial habenula in the brain were reversibly antagonized by suramin. Since then, many articles have described either the distribution of various P2 receptor subtypes in the brain and spinal cord or electrophysiological studies of the effects of purines in brain slices, isolated nerves, and glial cells (Burnstock, 2003b). Synaptic transmission has also been demonstrated in the myenteric plexus and in various sensory, sympathetic, and pelvic ganglia (Dunn et al., 2001). Adenosine, produced following ectoenzymatic breakdown of ATP, acts through presynaptic P1 receptors to inhibit the release of excitatory neurotransmitters in both the peripheral and the central nervous systems. P2Y receptors are expressed on both nonmyelinating and myelinating Schwann cells.
A. Neuroprotection
In the brain, P2 purinergic signaling is involved in the regulation of a variety of physiological and pathophysiological processes, including development and nervous tissue remodeling following trauma, stroke, ischemia, or neurodegenerative disorders (Burnstock, 2003a; Neary et al., 2004). Agonists and antagonists of adenosine and inhibitors of adenosine kinase are also being explored as therapeutic neuroprotective agents as well as for a number of other neurological conditions including epilepsy, sleep apnoea, pain, inflammation, aggression, anxiety, and memory loss (Kowaluk and Jarvis, 2000; Ribeiro et al., 2003; Fredholm et al., 2005; Köles et al., 2005). Microinjection of ATP analogs into the prepiriform cortex induces generalized motor seizures (Knutsen and Murray, 1997). P2X2, P2X4, and P2X6 receptors are expressed in the prepiriform cortex, suggesting that P2X receptor antagonists may have potential as neuroleptic agents. The hippocampus of chronic epileptic rats shows abnormal responses to ATP associated with increased expression of P2X7 receptors; it has been suggested that P2X7 receptors (perhaps on microglia) may participate in the pathophysiology of temporal lobe epilepsy (Vianna et al., 2002).
In nervous tissue, trophic factors ensure neuronal viability and regeneration. Neuronal injury releases fibroblast growth factor, epidermal growth factor, and platelet-derived growth factor (Neary et al., 1999). In combination with these growth factors, ATP can stimulate astrocyte proliferation, contributing to the process of reactive astrogliosis, and to hypertrophic/hyperplastic responses. P2Y receptor antagonists have been proposed as potential neuroprotective agents in the cortex, hippocampus, and cerebellum by modulation of kainate and α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid-induced currents, excessive activation of glutamate receptor systems being implicated in neuronal cell death associated with stroke, epileptic seizures, and neurodegenerative diseases such as Alzheimer's, Parkinson's, Huntington's, and amyotrophic lateral scoliosis (Zona et al., 2000). Release of ATP from disrupted cells may cause cell death in neighboring cells expressing P2X7 receptors, leading to a necrotic volume increase, which has been proposed as a cellular mechanism in the pathogenesis of Parkinson's disease (Jun and Kim, 2004). Guanine nucleotides inhibit NMDA and kainate-induced neurotoxicity in cultured rat hippocampal and neocortical neurons and may be candidates for antagonizing glutamate receptor-mediated neurotoxicity (Morciano et al., 2004). P2 receptors have been claimed to mediate neuroprotective effects in the cerebellum and the possible therapeutic use of P2 receptor agonists as neuroprotective agents has been raised (Volonte et al., 1999, 2003). It has been suggested that adenine is involved in the control of Purkinje cell survival (Watanabe et al., 2003). Up-regulation of P2X1 and P2X2 receptors after cerebellar lesions has been reported (Florenzano et al., 2002). Purine derivatives are in clinical trials as memory-enhancing agents in Alzheimer's disease; two of these, propentofylline and AIT-082, appear to act as trophic effectors, increasing the production of neurotrophic factors in brain and spinal cord (Rathbone et al., 1999). It has been reported that aluminum can produce Alzheimer-like symptoms, and a mechanism has been proposed whereby the aluminum binds to ATP to act on P2 purinoceptors leading to formation of amyloid fibrils (Exley and Korchazhkina, 2001). The therapeutic potential of adenosine A2A receptor antagonists in Parkinson's disease is being explored (Simola et al., 2004; Xu et al., 2005).
ATP inhibits the release of the excitatory transmitter, glutamate, and stimulates release of the inhibitory transmitter, GABA, from hippocampal nerves, thus serving a protective role (Inoue, 1998). In addition, ATP coreleased with glutamate induces long-term potentiation (LTP) in CA1 neurons associated with learning and memory (Fujii et al., 1999; Mori et al., 2001). Nanomolar concentrations of ATP induce long-lasting enhancement of LTP in hippocampal neurons; the P2 antagonist, suramin, inhibited activity of the ectoenzyme, apyrase, which has been shown to participate in the mechanisms of memory acquisition (Bonan et al., 1999). It has been suggested that ATP coreleased with glutamate activates CA1 pyramidal hippocampal neurons, allowing calcium to enter postsynaptic cells and thereby inhibiting the effectiveness of NMDA receptors in inducing LTP (Pankratov et al., 2002). Because P2X receptors contribute to synaptic transmission, mainly at low frequencies of stimulation, they may act as a dynamic low-frequency filter, preventing weak stimuli from inducing long-lasting changes in synaptic efficacy. It has also been claimed that ATP analogs can facilitate LTP through P2 receptor activation that triggers adenosine release, leading to activation of P1 (A2A) receptors (Almeida et al., 2003). Large rises in [Ca2+]i in CA1 neurons induce LTP, but small rises induce long-term depression (Yamazaki et al., 2002). ATP and activation of glutamate NMDA receptors leads to potentiation of LTP in CA1 neurons (Fujii et al., 2002) in keeping with the synergism that often occurs between cotransmitters (Burnstock, 2004). There is expression of functional P2X receptor channels in the axons of CA3 neurons branching to their postsynaptic targets and predominantly in nerve terminals forming synapses with interneurons (Khakh et al., 2003).
ATP, released from astrocytes, acts as an activity-dependent signaling molecule in neuron-glia communication, resulting in astrocyte Ca2+ waves and synaptic modulation; neuron-glia cross talk may represent an integral part of activity-dependent plasticity of neural networks (Zhang et al., 2003). Clearly there are multiple roles for P2 and P1 receptors in relation to learning and memory, but the way that therapeutic manipulation of purinergic mechanisms can be used to improve these functions is still unresolved. Higher order cognitive functions, including learning and memory in the prefrontal cortex appear to involve P2Y receptor signaling (Wirkner et al., 2003).
ATP injected into the supraoptic nucleus of the hypothalamus has antidiuretic effects (Mori et al., 1994). Purinergic and adrenergic synergism for vasopressin and oxytocin release is consistent with ATP cotransmission in the hypothalamus (Kapoor and Sladek, 2000). Purinergic signaling appears to play a significant role in the regulation of body temperature during fever by central hypothalamic and brainstem nuclei (Gourine et al., 2002, 2004). Functional interactions seem likely to occur between purinergic and nitrergic neurotransmitter systems; they may be important for regulation of hormone secretion and body temperature at the hypothalamic level and for cardiovascular and respiratory control at the level of the brainstem (Yao et al., 2003). ATP is coreleased with GABA or NA to act on P2 receptors, which are strongly expressed in most nuclei in the hypothalamus, including arcuate, paraventricular, retrochiasmatic, supraoptic, ventromedial, and dorsomedial (Matsumoto et al., 2004).
P2X and GABAA receptors play an important role in CO2 chemoreception and are involved in mediation of the ventilatory response to hypercapnia (Gourine and Spyer, 2003). P2X receptors expressed in neurons in the trigeminal mesencephalic nucleus may be involved in the processing of proprioceptive information (Khakh et al., 1997). The nucleus tractus solitarius (NTS) is a major integrative center in the brainstem that is involved in reflex control of the cardiovascular system; stimulation of P2X receptors in the NTS evokes hypotension with decreases in both cardiac output and total peripheral resistance (Kitchen et al., 2001). Different NTS purinoceptor subtypes may contribute to patterned autonomic responses observed in specific physiological or pathological situations (Scislo et al., 2001). Evaluation of the roles of purinergic signaling in processing of the sympathoexcitatory component of the chemoreflex at the NTS level may illuminate the mechanisms underlying the sympathetic overactivity observed in pathophysiological conditions such as hypertension, obstructive sleep apnea, and heart failure (De Paula et al., 2004).
In the striatum, extracellular ATP and adenosine are involved in the regulation of the feeding-associated mesolimbic neuronal activity in an antagonistic manner (Kittner et al., 2003). PPADS suppresses the feeding-evoked dopamine release in the nucleus accumbens, a brain region regarded as important for the regulation of appetite behavior and reinforcement (Kittner et al., 2000). Adenosine-dopamine interactions in the ventral striatum have been implicated in schizophrenia (Tsai, 2005). A hypothesis in which dysfunction of purinergic signaling (for example, decreased ATPase activity in erythrocytes, leading to increased levels of ATP and decreased adenosine) may lead to schizophrenia has been put forward (Lara and Souza, 2000).
It has been claimed that purinergic signaling dysfunction (perhaps largely reduced adenosinergic activity) is involved in mania and aggressive behavior (Machado-Vieira et al., 2002). Endogenous ATP has been claimed to be involved in the regulation of anxiety via stimulation of P2Y1 receptors in the dorsomedial hypothalamus in rats (Kittner et al., 2003). Chronically administered guanosine has anticonvulsant, amnesic, and anxiolytic effects in mice, perhaps associated with modulation of glutamatergic excitation (Vinade et al., 2003).
Multiple P2X and P2Y receptors have been identified on single cerebellar granule cells (Hervas et al., 2003). It has been reported that ATP continuously modulates the cerebellar circuit by increasing the inhibitory input to Purkinje neurons, probably via P2X5 and P2Y2 and/or P2Y4 receptor subtypes, thus decreasing the main cerebellar output activity, which contributes to locomotor coordination (Brockhaus et al., 2004).
Spinal cord traumas are a major health problem. ATP-MgCl2 has been shown to decrease lipid peroxidation in spinal cord injury and protect the spinal cord from secondary injury after trauma; it was concluded that ATP-MgCl2 should be explored for the treatment of spinal cord injuries in conjunction with other treatment modulators (Cakir et al., 2003). In a recent study, topical application of ATP after spinal cord injury significantly improved locomotor function (Shields et al., 2004).
Phenylketonuria is an innervated deficiency of phenylalanine hydroxylase activity in the liver, which causes increased brain levels of phenylalanine and its metabolites, leading to permanent brain damage in the early period of postnatal brain development. Phenylalanine has been shown to inhibit ATP diphosphohydrolase, resulting in increases in ATP levels, perhaps the neurotoxic mechanism underlying brain damage in this disease (Berti et al., 2001).
Acanthamoeba is a protozoan parasite that can cause fatal granulomatous amebic encephalitis. It has been shown recently to hydrolyze ATP, and it was suggested that this ecto-ATPase activity may play a role in the pathogenesis of this disease (Sissons et al., 2004).
Diadenosine tetraphosphate (Ap4A) protects against injuries induced by ischemia and 6-hydroxytryptamine in rat brain and has been suggested as a potentially useful target molecule in the therapy of stroke (Wang et al., 2003b). Up-regulation of P2X2 and P2X4 receptors in organotypic cultures of hippocampus, cortex, and striatum is associated with ischemic cell death and was prevented by P2 receptor antagonists (Cavaliere et al., 2003).
ATP is an extracellular signaling molecule between neurons and glial cells. ATP released by cellular damage and from astrocytes may be important in triggering cellular responses to trauma and ischemia by initiating and maintaining reactive astrogliosis, which involves striking changes in proliferation and morphology of astrocytes and microglia. Some of the responses to ATP released during brain injury are neuroprotective, but in some cases ATP contributes to the pathophysiology initiated after trauma (Fields and Stevens-Graham, 2002; Murakami et al., 2003; Neary et al., 2003; Meme et al., 2004). It has been claimed that P2Y2 receptors activate neuroprotective mechanisms in astrocytic cells (Chorna et al., 2004). In contrast, experimental infusion of ATP into nucleus accumbens or cerebral hemisphere of rats suggests that purines might be a signal for induction of malignant brain tumors. It has been claimed recently that ischemic brain injury is regulated by extracellular ATP-mediated interleukin (IL)-10 expression in microglia (Han et al., 2004). Multiple P2X and P2Y receptor subtypes are expressed by astrocytes, oligodendrocytes, and microglia (James and Butt, 2002). P2Y receptors mediate reactive astrogliosis via induction of COX-2, and P2Y receptor antagonists might counteract excessive COX-2 activation in both acute and chronic neurological disease (Brambilla et al., 1999). P2 receptors also mediate regulation of COX-2 in microglia (Choi et al., 2003). IL-1β-induced astrocyte activation is regulated by purinergic signaling; this is compatible with the hypothesis that nucleotides released under inflammatory conditions activate autocrine or paracrine signaling pathways that modulate inflammation (John et al., 2001).
Recent experiments suggest that astrocytes can sense the severity of damage in the central nervous system (CNS) via ATP release from damaged cells and can modulate the tumor necrosis factor-α-mediated inflammatory response, depending on the extracellular ATP concentration and corresponding type of astrocyte P2 receptor activated (Kucher and Neary, 2005). Thus, micromolar ATP/P2Y receptor activation may act to boost a moderate inflammatory response, whereas millimolar ATP/P2X receptor activation may prevent the perpetuation of a comparatively large inflammatory response perhaps by induction of apoptosis. Protein kinase B/Akt is a key signaling molecule that regulates cell survival, growth and metabolism and inhibits apoptosis. Traumatic brain injury activates Akt. When cortical astrocytes were subjected to mechanical strain, ATP was released, leading to Akt activation; PPADS attenuated the Akt activation (Neary et al., 2005). ATP can activate P2X7 receptors in astrocytes to release glutamate, GABA, and ATP, which regulate the excitability of neurons (Wang et al., 2002). ATP release during neuronal excitation or injury can enhance the inflammatory effects of cytokines and prostaglandin E2 in astrocytes and may contribute to the chronic inflammation seen in Alzheimer's disease (Xu et al., 2003). Astrocytic gap junctions are involved in the neuroprotective process, in particular, to protect neurons from oxidative stress and glutamate toxicity (Naus et al., 2001). ATP released from astrocytes is essential in mediating the injury-induced responses of microglial processes (Davalos et al., 2005).
Microglial cells are the major cellular elements with immune functions inside the CNS and play important roles in orchestrating inflammatory brain responses to trauma and hypoxia. They express multiple P2 receptors (Bianco et al., 2005) and are activated by purines and pyrimidines to release inflammatory cytokines such as IL-1β and IL-6 and tumor necrosis factor-α. Activated microglia can also act as scavenger cells that induce apoptosis in damaged neurons by releasing toxic factors, including NO (Sanz and Di Virgilio, 2000; Inoue, 2002). Thus, whereas microglia may play an important role against infection in the CNS, overstimulation of this immune reaction may accelerate the neuronal damage caused by ischemia, trauma, or neurodegenerative diseases such as Alzheimer's and Parkinson's disease, human immunodeficiency virus encephalopathy, multiple sclerosis, and amyotrophic lateral scoliosis, which exhibit microglial proliferation and activation (Ogata et al., 2003). These authors showed that ATP inhibits cytokine release from lipopolysaccharide-activated microglia via P2Y receptors and suggested that P2Y agonists may be a potential treatment for toxic immunoreactions. P2X4 receptors, induced in spinal microglia, gate tactile allodynia after nerve injury (Tsuda et al., 2003) (see section II.B.). P2X7 receptors mediate superoxide production in primary microglia and are up-regulated in a transgenic model of Alzheimer's' disease, particularly around β-amyloid plaques (Parvathenani et al., 2003). Stimulation of microglial P2X7 receptors also leads to enhancement of interferon-γ-induced type II nitric oxide synthase activity (Gendron et al., 2003). P2X7 receptors may therefore provide a therapeutic target for inflammatory responses seen in neurodegenerative disorders.
P2 receptors are expressed by oligodendrocytes. However, P2 receptors on oligodendrocytic progenitor cells mediate an increase in [Ca2+]i and may mediate the formation of myelin, raising the possibility that activation of P2 receptors may offer new approaches to the treatment of demyelinating diseases in the CNS, such as multiple sclerosis (Stevens et al., 2002; Agresti et al., 2005). P2X receptors expressed by Schwann cells may regulate the synthesis and release of cytokines during pathophysiological events (Colomar and Amedee, 2001; Irnich et al., 2001).
B. Migraine
Classical migraine is associated with two distinct cerebrovascular phases: an initial vasoconstriction (not associated with pain) followed by vasodilatation (reactive hyperemia) associated with pain. The “purinergic” hypothesis for migraine was originally put forward in 1981 as a basis for the reactive hyperaemia and pain during the headache phase (Burnstock, 1981). It was suggested that ATP and its breakdown products adenosine 5′-monophosphate and adenosine were strong contenders for mediating the vasodilatation following the initial vasospasm and subsequent hypoxia. ATP was also implicated in the pathogenesis of pain during migraine via stimulation of primary afferent nerve terminals located in the cerebral microvasculature. Later studies have shown that the ATP-induced cerebral vasodilatation is endothelium-dependent via activation of P2Y receptors on the endothelial cell surface and subsequent release of endothelium-derived relaxing factor and that the endothelial cells are the main local source of ATP involved, although adenosine 5′-diphosphate and ATP released from aggregating platelets may also contribute to this vasodilatation. These findings have extended the purinergic hypothesis for migraine in two ways. First, they have clarified the mechanism of purinergic vasodilatation during the headache phase of migraine. Second, they suggest that a purinergic mechanism may also be involved in the initial local vasospasm, via P2X receptors on smooth muscle cells occupied by ATP released either as a cotransmitter with NA from perivascular sympathetic nerves or from damaged endothelial cells (Burnstock, 1989). The hypothesis has gained further support by the identification of P2X3 receptors on primary afferent nerve terminals arising from trigeminal, nodose, and spinal ganglia (Chen et al., 1995; Burnstock, 2001b). Thus, P2X3 receptor antagonists may be candidates for antimigraine drug development (Waeber and Moskowitz, 2003). There is also recent evidence that migraine is a chronic sympathetic nervous system disorder, with which there is an increase in release of sympathetic cotransmitters, including ATP (Peroutka, 2004), which may contribute to the initial vasospasm (Macdonald, 2000).
C. Pain
The involvement of ATP in the initiation of pain was recognized early (Collier et al., 1966; Bleehen and Keele, 1977; Burnstock, 1981; Jahr and Jessell, 1983). A major advance was made when the P2X3 ionotropic receptor was cloned in 1995 (Chen et al., 1995; Lewis et al., 1995) and shown later to be predominantly localized in the subpopulation of small nociceptive sensory nerves that label with isolectin B4 (IB4) in DRG (Bradbury et al., 1998). Burnstock (1996b) put forward a unifying purinergic hypothesis for the initiation of pain by ATP acting via P2X3 and P2X2/3 receptors associated with causalgia, reflex sympathetic dystrophy, angina, migraine, and pelvic and cancer pain. This has been followed by an increasing number of papers expanding on this concept for acute, inflammatory, neuropathic, and visceral pain (see reviews by Burnstock and Wood, 1996; Chizh and Illes, 2000; Burnstock, 2001c; Dunn et al., 2001; Cooke et al., 2003; Gu, 2003; Inoue et al., 2003; Jarvis, 2003; Sawynok and Liu, 2003; Ueda and Rashid, 2003; North, 2004). Sensory terminals are sensitive to ATP released from local cells during mechanical stress and to α,β-meATP in the tongue (Rong et al., 2000), tooth pulp (Alavi et al., 2001; Renton et al., 2003), bladder (Cockayne et al., 2000; Vlaskovska et al., 2001; O'Reilly et al., 2002; Rong et al., 2002), ureter (Knight et al., 2002; Rong and Burnstock, 2004), gut (Wynn et al., 2003, 2004), and uterine cervix (Papka et al., 2005). Both P2X3 (homomultimer) and P2X2/3 (heteromultimer) receptors mediate nociceptive afferent responses, but the proportions vary in different organs. P2Y1 receptors have also been demonstrated in a subpopulation of sensory neurons that colocalise with P2X3 receptors (Ruan and Burnstock, 2003; Gerevich et al., 2004).
The search is on for selective P2X3 and P2X2/3 receptor antagonists that do not degrade in vivo. PPADS is a nonselective P2 antagonist, but has the advantage that it associates and dissociates approximately 100 to 10,000 times more slowly than other known antagonists (Spelta et al., 2002). The trinitrophenyl-substituted nucleotide TNP-ATP is a very potent antagonist at both P2X3 and P2X2/3 receptors (Mockett et al., 1994; King et al., 1997; Virginio et al., 1998; Burgard et al., 2000; Honore et al., 2002). A-317491 is a potent and selective non-nucleotide antagonist of P2X3 and P2X2/3 receptors, and it reduces chronic inflammatory and neuropathic pain in the rat (Jarvis et al., 2002, 2004; McGaraughty et al., 2003). Antisense oligonucleotides have been used to down-regulate the P2X3 receptor, and in models of neuropathic (partial sciatic nerve ligation) and inflammatory (complete Freund's adjuvant) pain, inhibition of the development of mechanical hyperalgesia as well as significant reversal of established hyperalgesia, were observed within 2 days of treatment (Barclay et al., 2002; Honore et al., 2002; Stone and Vulchanova, 2003). P2X3 antisense oligonucleotides or antagonists appear to be less effective for treating discogenic (lumbar intervertebral disc) than cutaneous tissue pain (Aoki et al., 2003). Combined antisense and RNA interference-mediated treatment for specific inhibition of the recombinant rat P2X3 receptor appears to be promising for pain therapy (Hemmings-Mieszczak et al., 2003). P2X3 double-stranded short interfering RNA relieves chronic neuropathic pain and opens up new avenues for therapeutic pain strategies in man (Dorn et al., 2004). Cytotoxic targeting using the IB4-saporin conjugate of isolectin IB4-binding nociceptive sensory neurons decreases the severity to noxious stimuli (Vulchanova et al., 2001; Nishiguchi et al., 2004). Antagonism of P2X1 and P2X3 receptors by phenol red has recently been reported (King et al., 2005) and shown to cause significant increases in the pressure and volume threshold required to initiate the micturition reflex in female urethra-anesthetized rats (King et al., 2004). Modulation of neurotransmission through P2X3 receptors in central and peripheral nervous systems may contribute to the anesthesia and analgesia produced by barbiturates (Kitahara et al., 2003). Tetramethylpyrazine, a traditional Chinese medicine used as an analgesic for dysmenorrhea, was shown to block P2X3 receptor signaling, inhibit significantly the first phase of nociceptive behavior induced by 5% formalin, and attenuate slightly the second phase in the rat hindpaw pain model (Liang et al., 2004, 2005). Interactions between vanilloid and metabotropic P2Y receptors are also being explored in terms of treatments for chronic pain (Premkumar, 2001) and thermal hypersensitivity (Moriyama et al., 2003). It has been reported recently that antagonism of spinal P2X3/P2X2/3 receptors results in an indirect activation of the opioid system to alleviate inflammatory hyperalgesia and chemogenic nociception (McGaraughty et al., 2005).
For neuropathic pain, the tactile allodynia that follows peripheral nerve injury is reduced by A-134974, a novel adenosine kinase inhibitor acting at spinal sites (Zhu et al., 2001). Endogenous ATP acting on P2X receptors appears to be necessary for the induction of the postoperative pain characterized by mechanical allodynia (Tsuda et al., 2000, 2001). Up-regulation of P2Y1 receptor expression in the DRG occurs after transection of sciatic nerves (Xiao et al., 2002). P2X4 receptors are induced in spinal microglia that appear to gate tactile allodynia after nerve injury (Tsuda et al., 2003, 2005; Inoue et al., 2004, 2005). Intraspinal administration of p38 inhibitor suppressed allodynia, which suggests that neuropathic pain hypersensitivity depends on the activation of the p38 signaling pattern in microglia in the dorsal horn following peripheral nerve injury (Inoue et al., 2003). Suramin inhibits spinal cord microglia activation and long-term hyperalgesia induced by inflammation produced by formalin injection (Wu et al., 2004; Guo et al., 2005). Analgesic effects with intrathecal administration of P2Y receptor agonists UTP and UDP in normal and the neuropathic pain rat model have been reported, suggesting that P2Y2 (and/or P2Y4) and P2Y6 receptors produce inhibitory effects in spinal pain transmission (Okada et al., 2002). Disruption of the P2X7 receptor gene abolishes chronic inflammatory and neuropathic pain (Chessell et al., 2005). Purinergic mechanisms are also beginning to be explored in relation to cancer pain (Burnstock, 1996; Cain et al., 2001; Mantyh et al., 2002; Gilchrist et al., 2005).
D. Diseases of Special Senses
1. Eye.
Purinergic signaling is widespread in the eye (Pintor, 2000) and novel therapeutic strategies are being developed for glaucoma, dry eye, and retinal detachment (Pintor et al., 2003a). P2Y receptors on human corneal epithelial cells appear to play a critical role in the injury-repair process (Klepeis et al., 2004).
ATP, acting via both P2X and P2Y receptors, modulates retinal neurotransmission, affecting retinal blood flow and intraocular pressure. The ATP analog β,γ-methylene ATP is more effective in reducing intraocular pressure (40%) than muscarinic agonists such as pilocarpine (25%) and β-adrenoceptor blockers (30%), raising the potential for the use of purinergic agents in glaucoma (Pintor and Peral, 2001). Dinucleoside polyphosphates acting via P2Y1 receptors on trabecular network cells increase aqueous humor outflow and may be another target for antiglaucomatous drugs (Soto et al., 2005). Suramin, a P2 receptor antagonist, has been shown to inhibit the fibrotic wound healing reactions that sometimes follow trabeculectomies for surgically treating eyes with glaucoma (Mietz et al., 1998). Topical application of Ap4A has been proposed for the lowering of intraocular pressure in glaucoma (Pintor et al., 2003b). The formation of P2X7 receptor pores and apoptosis is enhanced in retinal microvessels early in the course of experimental diabetes, suggesting that purinergic vasotoxicity may play a role in microvascular cell death, a feature of diabetic retinopathy (Sugiyama et al., 2004).
P2Y2 receptor activation increases salt, water, and mucus excretion and thus represents a potential treatment for dry eye conditions (Murakami et al., 2000; Yerxa, 2001). In the pigmented layer of the retina, P2Y2 receptor activation promotes fluid absorption and may be involved in retinal detachment. INS37217, a long-lasting synthetic P2Y2 receptor agonist, stimulates the retinal pigment epithelium by activating P2Y2 receptors at the apical membrane, and in vivo treatment enhances the rate of subretinal fluid reabsorption in experimentally induced retinal detachments and may be useful for treating a variety of retinal diseases that result in fluid accumulation in the subretinal space (Maminishkis et al., 2002). Reactive responses of Müller cells occur within 24 h of retinal detachment. Suramin inhibits some of these responses and may provide a therapeutic candidate to limit the detrimental effects of immune cell activation and Müller cell gliosis during retinal detachment (Uhlmann et al., 2003).
ATP and UTP restore the rates of both net Cl- and fluid secretion in adenovirus type 5-infected conjunctival tissues and are considered as potential therapeutic modulators for the treatment of various transport defects encountered in ocular tissues in diseased and/or inflamed states (Kulkarni et al., 2003). UTP and Ap4A accelerate wound healing in the rabbit cornea, by regulating the rate of epithelial cell migration (Pintor et al., 2004).
The UPL rat is a dominant hereditary cataract model derived from Sprague-Dawley rats and has been used to show that Ca2+-ATPase expression increases, whereas ATP control decreases in lenses during the development of the cataract and opacification; disulfiram and aminoguanidine, which inhibit inducible NO and scavenge reactive oxygen species, attenuate the decrease in ATP, resulting in a delay in cataract development (Nabekura et al., 2004).
A recent article has raised the possibility that alterations in sympathetic nerves may underlie some of the complications observed in diabetic retinopathy (Wiley et al., 2005); ATP is well established as a cotransmitter in sympathetic nerves.
2. Ear.
In the auditory system ATP, acting via P2Y receptors, depresses sound-evoked gross compound action potentials in the auditory nerve and the distortion produces otoacoustic emission, the latter being a measure of the active process of the outer hair cells (Housley, 2000; Sueta et al., 2003). P2X splice variants are found on the endolymphatic surface of the cochlear endothelium, an area associated with sound transduction. Both P2X and P2Y receptors have been identified in the vestibular system (Xiang et al., 1999). ATP may regulate fluid homeostasis, cochlear blood flow, hearing sensitivity and development, and thus may be useful in the treatment of Ménière's disease, tinnitus, and sensorineural deafness.
Sustained loud noise produces an up-regulation of P2X2 receptors in the cochlea, particularly at the site of outer hair cell sound transduction. P2X2 expression is also increased in spiral ganglion neurons, indicating that extracellular ATP acts as a modulator of auditory neurotransmission that is adaptive and dependent on the noise level (Wang et al., 2003a). Excessive noise can irreversibly damage hair cell stereocilia leading to deafness. Data have been presented showing that release of ATP from damaged hair cells is required for Ca2+ wave propagation through the support cells of the organ of Corti, involving P2Y receptors, and this may constitute the fundamental mechanism to signal the occurrence of hair cell damage (Munoz et al., 2001; Gale et al., 2004). ATP is claimed to mitigate the effects of noise trauma (Jakobi et al., 1977; Sugahara et al., 2004), although the mechanisms involved are not clear.
3. Nasal Organs.
Purinergic receptors have been described in the nasal mucosa, including the expression of P2X3 receptors on olfactory neurones (Gayle and Burnstock, 2005). Enhanced sensitivity to odors in the presence of P2 purinergic antagonists suggests that low-level endogenous ATP normally reduces odor responsiveness (Hegg et al., 2003a). It appears that the induction of heat-shock proteins by noxious odor damage can be prevented by the in vivo administration of P2 receptor antagonists (Hegg et al., 2003b). The predominantly suppressive effect of ATP in odor responses could play a role in the reduced odor sensitivity that occurs during acute exposure to noxious fumes and may be a novel neuroprotective mechanism.
IV. Immune System and Inflammation
ATP and adenosine are released at sites of inflammation. ATP is involved in the development of inflammation through a combination of actions: release of histamine from mast cells, provoking production of prostaglandins; and the production and release of cyokines from immune cells (Di Virgilio et al., 1998). In contrast, adenosine exerts anti-inflammatory actions. P2X7 and P2Y1 and P2Y2 receptors located on inflammatory and immune cells play a pivotal role in inflammation and immunomodulation (Di Virgilio et al., 1998; Luttikhuizen et al., 2004). In addition, NTPDases have effects on immune reactions (Robson et al., 2001). NTPDase1 deficiency appears to have two opposing impacts on nucleotide-mediated signaling, that is, inhibition via desensitization of some, but not all, P2 receptors and augmentation of certain other responses via impaired hydrolysis of their ligands. Consequently, there are defects in dendritic cell function, antigen presentation, T-cell responses to haptens (type IV hypersensitivity reactions), and delayed cellular rejection responses under partial costimulation blockade in NTPDase1-null mice.
In addition to the roles of purines in inflammation, they have a broad range of functions carried out through purinergic receptors on immune cells, including killing intracellular pathogens by inducing apoptosis of host macrophages, chemoattraction, and cell adhesion (Burnstock, 2001d; Di Virgilio et al., 2001). Purinergic compounds may turn out to be useful for the treatment of neurogenic inflammation, rheumatoid arthritis, and periodontitis (Dubyak and El Moatassim, 1993).
ATP-induced apoptosis in macrophages via P2X7 receptors, also results in killing of the mycobacteria contained within them, in contrast, to the macrophage apoptosis produced by other agents. Elucidation of the bacterial killing mechanism initiated by the P2X7 receptor may help to devise new strategies to combat the most potent and enduring of human pathogens (Lammas et al., 1997; Saunders et al., 2003). Infection with Mycobacterium tuberculosis causes macrophages to release ATP, which leads to oxygen radical production, providing antibacterial effects at sites of infection (Sikora et al., 1999). In later studies, it was shown that the ATP-induced bactericidal activity toward virulent M. tuberculosis requires an increase in cytosolic Ca2+ in infected macrophages, and it was hypothesized that the Ca2+ dependence was linked to promotion of phagosome-lysosome fusion (Kusner and Barton, 2001; Fairbairn et al., 2001). The P2X7 receptor plays a fundamental role in lipopolysaccharide signal transduction and activation of macrophages and may therefore represent a therapeutic target for Gram-negative bacterial septicemia (Sommer et al., 1999). Vibrio cholerae, the causative organism of the intestinal disease cholera, secretes enzymes that lead to ATP degradation; this may allow the pathogen to evade the immune system by reducing the apoptotic actions of the P2X7 receptor (Punj et al., 2000). Evidence has been presented to support the view that, whereas the cytotoxic actions of ATP on macrophages were via P2X7 receptors, the bacteriocidal effects of ATP (and UTP) were probably via P2Y2 receptors (Stober et al., 2001). It is conceivable that various pathogenic bacteria might secrete adenine nucleotide-interconverting enzymes during host cell infection to modulate host cell purinergic receptors to their own advantage (Yamada and Chakrabarty, 2004). Lesional accumulation of macrophages expressing P2X4 receptors in rat CNS during experimental autoimmune encephalomyelitis has been described recently, and it was suggested that P2X4 receptors might be valuable markers to dissect the local monocyte heterogeneity in autoimmune disease (Guo and Schluesener, 2005). ATP moderates anti-IgE-induced release of histamine from lung mast cells and may therefore be mechanistically involved in human allergic/asthmatic reactions (Schulman et al., 1999). Alveolar macrophages express P2X7 receptors, which upon stimulation trigger proinflammatory responses, including activation of IL-1 to IL-6 cytokines and granulomatous reactions (Lemaire and Leduc, 2004). In contrast, ATP and ADP inhibit cytokine generation by human mast cells through P2Y receptors and could present a novel therapeutic target for asthma, arthritis and other disorders with a prominent contribution from mast cells and their products (Feng et al., 2004).
Extracellular ATP inhibits the activation of CD4+ T lymphocytes via P2Y receptors, which suggests a novel therapeutic target for topical immunosuppression in eye, skin, or airway inflammatory disease (Duhant et al., 2002). In addition to the apoptosis mediated by P2X7 receptors, a lower level of activation sometimes results in cell proliferation; it has been suggested that the expression and function of P2X7 receptors on B lymphocytes may correlate with the severity of B-cell chronic lymphocytic leukemia (Adinolfi et al., 2002).
During the acute phase of Trypanosoma cruzi infection, the etiologic agent of Chagas' disease, thymic atrophy occurs; ATP also induces cell death in CD4+/CD8+ double-positive thymocytes and may play a central role in thymus atrophy during T. cruzi infection (Mantuano-Barradas et al., 2003). Infection by the parasitic blood fluke Schistosoma mansoni also leads to thymic atrophy. The cloning and characterization of a P2X receptor (schP2X) from S. mansoni provide the first example of a nonvertebrate ATP-gated ion channel and may provide an alternative drug target for the treatment of schistosomiasis (Agboh et al., 2004).
A novel mechanism by which ATP, probably via P2Y11 receptors, can regulate the trafficking of specific dendritic cell populations has been described (Schnurr et al., 2003). The migration of dendritic cells from the site of antigen capture to lymphoid tissue is a prerequisite for the induction and regulation of immune responses. Therefore, ATP-mediated inhibition of migration could play an important role in inflammatory disease and cancer. Targeting of P2Y11 receptors may provide a new therapeutic strategy to improve the migration of dendritic cells to induce the trafficking of antigen from the vaccine site to the draining lymph nodes.
Allopurinal and captopril have a therapeutic effect in granulomatous disorders, such as sarcoidosis, by a direct action on monocyte/macrophage lineage cells partly by down-regulation of intracellular adhesion molecular-1 and P2X7 receptors (Mizuno et al., 2004). ATP and UTP have been shown to be potent stimulators of human hematopoietic stem cells both in vitro and in vivo (Lemoli et al., 2004). Thus, these extracellular nucleotides may provide a novel and powerful tool to modulate hematopoietic stem cell function to increase the number of transplantable cells in vivo in the event of bone marrow failure. P2X7 receptors control endocannabinoid production by microglia cells and might constitute promising therapeutics to temper exacerbated microinflammatory responses and allied cell damage (Witting et al., 2004).
V. Endocrinology, Diabetes, and Obesity
Purinoceptors are widely expressed in endocrine glands (Burnstock and Knight, 2004). For example, ATP and UTP increase cytosolic free calcium in human thyrocytes (Schofl et al., 1995), ATP modulates aldosterone production by adrenal cortex (Szalay et al., 1998), ATP regulates prolactin release from the anterior pituitary (Stojilkovic et al., 2000) and vasopressin and oxytocin secretion from the posterior pituitary (Sperlágh et al., 1999); ATP stimulates insulin release from the endocrine pancreas (Coutinho-Silva et al., 2001); ATP and UTP inhibit estradiol and progesterone secretion from the ovary (Tai et al., 2001) and mediate increases in intracellular calcium in Sertoli cells from testis (Ko et al., 2003). Ovariectomy significantly increased the hydrolysis of ATP, ADP, and AMP in rat blood serum, whereas estradiol replacement therapy significantly decreased the hydrolysis of adenine nucleotides, suggesting a relationship between ecto-ATPases and the hormonal system (Pochmann et al., 2004).
A diversity of actions of purines in the pituitary gland have been described, including trophic effects and cytokine production as well as actions in hormone release, with implications for pathological as well as physiological states (Rees et al., 2003). 5′-Nucleotidase activity is increased in synaptosomes from hippocampus and cerebral cortex of hypothyroid rats and may be related to the cognitive disorders found in hypothyroidism (Bruno et al., 2005a). 5′-Nucleotidase activity is also altered in platelets by hypo- and hyperthyroidism and may represent an additional mechanism by which disturbances related to thyroid hormones are associated with some vascular diseases (Bruno et al., 2005b).
A. Diabetes
Early studies from the laboratory of Loubatière-Mariani et al. (1997) showed that P2Y receptors are present on pancreatic β-cells and are involved in insulin secretion. ATP stimulates pancreatic insulin release through a glucose-dependent P2Y receptor-mediated mechanism and also modulates insulin secretion through interactions with ATP-sensitive potassium channels in islet β-cells. Biotin enhances ATP synthesis in pancreatic islets, resulting in reinforcement of glucose-induced insulin secretion (Sone et al., 2004).
P2Y purinoceptors appear to be impaired in fibroblasts from type 2 diabetic patients, which results in reduced glucose uptake via the glucose transporter GLUT 1, suggesting that P2Y receptors may be candidate targets for the design of innovative antidiabetic drugs (Solini et al., 2003). Hormone-specific defects in insulin regulation of ATPase that may contribute to their insulin resistance are seen in non-insulin-dependent diabetic rats (Levy et al., 1994).
A feature of diabetic retinopathy is the apoptotic death of microvascular pericytes and endothelial cells; there appears to be an enhancement of P2X7 receptor-induced pore formation and apoptosis on the retinal microvasculature in early diabetes (Sugiyama et al., 2004). In streptozotocin-induced diabetic animals, P2X7 receptor expression, located in glucagon-containing α-cells in pancreatic islets, increases, and the expression migrates centrally to take the place of the insulin-containing β-cells, although the functional significance of this is not known (Coutinho-Silva et al., 2003). The potential role of purinergic compounds as novel treatments for diabetes has yet to be explored. Stimulation of insulin secretion and improvement of glucose tolerance in rats and dogs by the P2Y receptor agonist adenosine-5′-(2-thiodiphosphate) has been claimed (Hillaire-Buys et al., 1993).
B. Obesity
ATP, released as a cotransmitter from sympathetic nerves, has been shown to stimulate brown adipocytes (Lee and Pappone, 1997; Omatsu-Kanbe et al., 2002). Deficits in receptor regulation, transporter mobilization, and adipocyte hormone secretion are all thought to contribute to the pathology of obesity (Flier, 1995; Schmidt and Loffler, 1998). Stimulation of lipogenesis in rat adipocytes by ATP, which regulates fat stores independently from established hormones has been reported (Schodel et al., 2004). A recent article presents evidence that adipocytes express two different P2Y receptor subtypes and that activation of P2Y11 receptors might be involved in inhibition of insulin-stimulated lepton production and stimulation of lipolysis (Lee et al., 2005). Some of the effects of ATP are caused by its breakdown product, adenosine, which has been known for a long time to be involved in the activities of adipocytes (see Fredholm, 1981).
VI. Gastroenterology
Purinergic signaling plays a major role in different activities of the gut (Burnstock, 2001b; Galligan, 2002; Giaroni et al., 2002; Lecci et al., 2002; Nassauw et al., 2002; Poole et al., 2002; Bertrand, 2003; Kadowaki et al., 2003; Xiang and Burnstock, 2004a,b). ATP is a cotransmitter in NANC nerves responsible for the inhibitory phase in peristalsis, it participates in synaptic transmission in the myenteric and submucosal ganglia, and it is involved in vascular control of the gastrointestinal tract and in the control of mucosal secretion. Both glial cells (Kimball and Mulholland, 1996; Vanderwinden et al., 2003) and the interstitial cells of Cajal (Burnstock and Lavin, 2002) express P2 receptors, although their roles have yet to be clarified.
A limited number of studies have been conducted to date on changes in purinergic signaling in the diseased gut. ATP and adenosine have been implicated in the development of gastric ulcers, Hirschsprung's and Chagas' diseases, ischemia and colonic tumors (Burnstock, 2001c). Extracellular nucleotides and their receptors have been implicated in the pathogenesis of inflammatory bowel disease (IBD) (Somers et al., 1998). T cells are thought to play a primary role in the induction of epithelial cell damage in IBD, and the P2Y6 receptor was found to be highly expressed on the T cells infiltrating IBD, but absent in T cells of unaffected bowel. This suggests that the P2Y6 receptor and its selective agonist, UDP, may play a role in the pathogenesis of IBD. Later articles have shown that P2Y6 receptors are involved in monocytic release of IL-8 and stimulation of NaCl secretion (Kottgen et al., 2003). During inflammation of the gastrointestinal tract, glial cells proliferate and produce cytokines; thus, P2X7 receptors may play a role in the response of enteric glia to inflammation (Vanderwinden et al., 2003). P2X3 purinergic signaling enhancement in an animal model of colitis has been described (Wynn et al., 2004). P2X3 receptor expression is increased in the enteric plexuses in human irritable bowel syndrome, suggesting a potential role in dysmotility and pain (Yiangou et al., 2001), and the possibility that P2X receptors are potential targets for the drug treatment of irritable bowel syndrome has been raised (Galligan, 2004). Bile induces ATP depletion and contributes to the early mucosal permeability alteration and barrier lesions that occur during experimental esophageal reflux (Szentpali et al., 2001). P2X3 immunohistochemistry has been demonstrated in aganglionic bowel in Hirschsprung's disease, suggesting that the sensory nerves may form a significant proportion of its hypertrophic innervation (Facer et al., 2001). P2Y receptors on smooth muscle and ATP production in myenteric neurons increase in postoperative ileus, probably contributing to delayed colonic transit (Wang et al., 2004a). Recent reviews have highlighted the potential of purinergic drugs for the treatment of functional bowel disorders and visceral pain (Holzer, 2001; Kirkup et al., 2001; Galligan, 2004). Intestinal epithelial cells from patients with cystic fibrosis fail to consistently conduct Cl- in response to ATP and UTP that elevate intracellular Ca2+ and this may be of value in the design of treatments to ameliorate gastrointestinal symptoms of cystic fibrosis (Smitham and Barrett, 2001).
Intrinsic sensory neurons in the submucous plexus of the gut, as well as extrinsic sensory nerves, show positive immunoreactivity for P2X3 receptors (Xiang and Burnstock, 2004b). It has been proposed (Burnstock, 2001b) that during moderate distension, low threshold intrinsic enteric sensory fibers may be activated via P2X3 receptors by ATP released from mucosal epithelial cells, leading to reflexes concerned with propulsion of material down the gut. Studies showing that peristalsis is impaired in the small intestine of mice lacking the P2X3 receptor subunit support this view (Bian et al., 2003). In contrast, during substantial (colic) distension associated with pain, higher threshold extrinsic sensory fibers may be activated by ATP released from the mucosal epithelia; these fibers pass messages through the DRG to pain centers in the CNS (Wynn et al., 2003, 2004). A recent study suggests that peripheral sensitization of P2X3 receptors on vagal and spinal afferents in the stomach may contribute to dyspeptic symptoms and the development of visceral hyperalgesia (Dang et al., 2005).
In the liver, purinergic receptors have been identified in the plasma membrane of the two principal epithelial cell types that form the bile-secreting unit, namely, hepatocytes, which constitute the liver parenchymal cells, and cholangiocytes, which line the lumen of intrahepatic bile ducts (Nathanson et al., 2001; Feranchac and Fitz, 2003). Activation of the receptors has been linked to several fundamental responses important to cellular metabolism, ion channel activation, cell volume regulation, and bile formation. It is suggested that pharmacological modulation of ATP release and purinergic signaling might provide novel strategies for the management of cholestasis and other disorders characterized by impaired bile flow. Purinergic receptors are present on both quiescent and activated hepatic stellate cells; quiescent cells express P2Y2 and P2Y4 receptors activated by UTP and ATP, whereas activated cells express P2Y6 receptors activated by UDP and ATP (Dranoff et al., 2004). It was speculated by these authors that the P2Y receptors on satellite cells might be an attractive target to prevent or treat liver fibrosis, via regulation of procollagen-1 transcription. ATP has been shown recently to rapidly activate multiple components of the c-Jun NH2-terminal kinase cascade, a central player in hepatocyte proliferation and liver regeneration (Thevananther et al., 2004). This study identifies extracellular ATP as a hepatic mitogen with implications about the regulation of liver growth and repair. Sympathetic nerves using NA and ATP as cotransmitters alleviate immune-mediated experimental hepatitis in the mouse; it is speculated that nerve-immune cell interactions may offer novel therapeutic strategies in immune and inflammatory liver diseases (Neuhuber and Tiegs, 2004).
The effect of ATP on salivary glands has been recognized since 1982. Both P2X and P2Y subtypes are expressed and opportunities for utilization of these receptors as pharmaceutical targets for diseases involving salivary gland dysfunction appear promising (Turner et al., 1999; Arreola and Melvin, 2003).
VII. Urogenital Diseases
A. Kidney and Ureter
There is a substantial presence of purinoceptors in different regions of the nephron, the glomerulus, and renal vascular system in the kidney, including subtypes involved in the regulation of renin secretion, glomerular filtration, and transport of water, ions, nutrients, and toxins (Chen and Chen, 1998; Unwin et al., 2003; Bailey et al., 2004; Shirley et al., 2005) (Fig. 2). The distribution of NTPDase1 and NTPDase2 parallels the distribution of P2 receptors in the kidney and is likely to influence physiological as well as pathophysiological events in the kidney (Kishore et al., 2005). ATP and adenosine have been used to protect kidneys from renal ischemic-reperfusion injury, and are being explored for the treatment of chronic renal failure and transplantation-induced erythrocytosis (Jackson, 2001). It has been proposed that luminal P2 receptors in the nephron are part of an epithelial “secretory” defense mechanism against bacteria or harmful particles involved in the regulation of cell volume when transcellular solute transport is out of balance. P2Y receptors mediate renin secretion (Churchill and Ellis, 1993). ATP released from macula densa cells serves as a major paracrine agent, mediating tubuloglomerular feedback signals to regulate afferent arteriolar resistance (Nishiyama and Navar, 2002). ATP exerts a dual effect on mesangial extracellular matrix production, stimulatory probably via P2X7 receptors and inhibitory via a P2Y receptor (Solini et al., 2005). In the presence of elevated ATP levels in inflammatory or ischemic conditions, extracellular matrix proteins accumulate due to functional predominance of P2X receptors. This mechanism might participate in the pathogenesis of mesangial expression that occurs in diabetes.
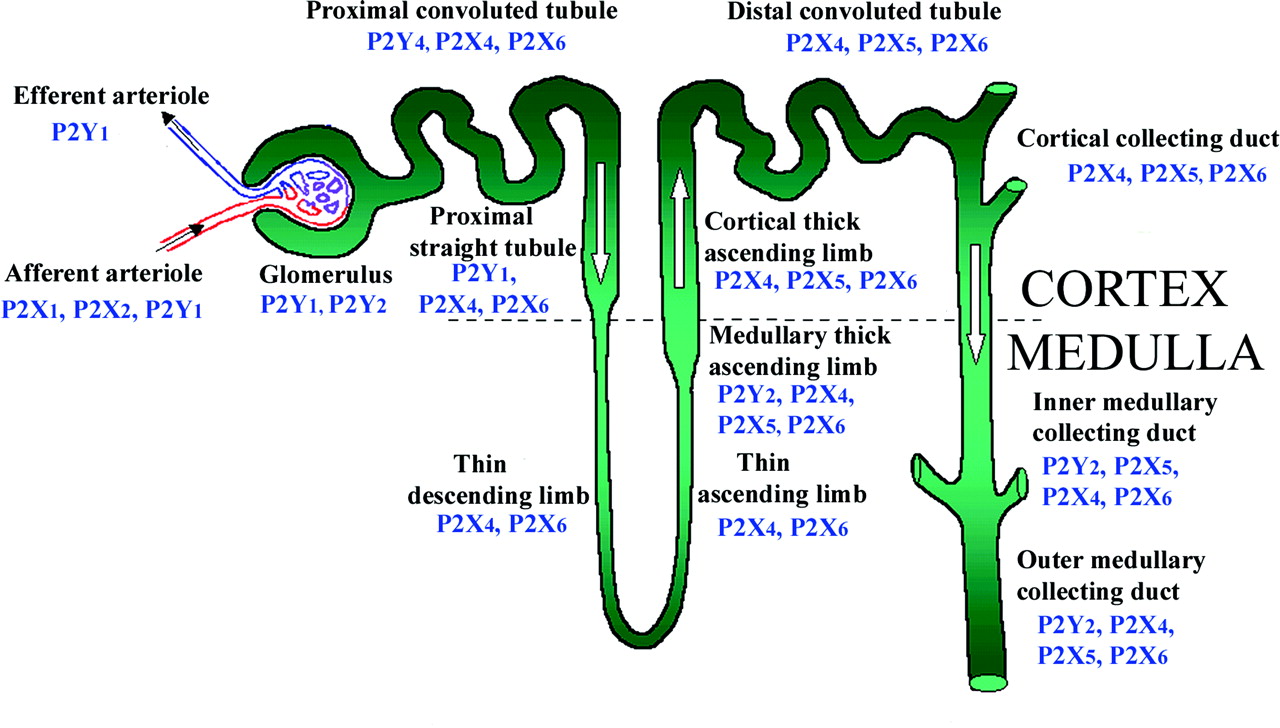
Summary of the nephron segments and the distribution of P2 receptor subtypes. Based on Turner et al. (2003), with permission from S. Karger AG (Basel, Switzerland).
Cyclosporine is a potent immunosuppressive agent, but its use has been limited by the side effect of nephrotoxicity; however, ATP treatment after verapamil pretreatment greatly reduces the nephrotoxic potential (Sumpio et al., 1987). The P2 receptor antagonist, PPADS, has been shown to be an effective inhibitor of mesangial cell proliferation in an experimental rat model of mesangial proliferative glomerulonephritis (Rost et al., 2002).
In polycystic kidney disease, tubules are altered, leading to dilated tubules or cysts encapsulated by a single monolayer of renal epithelium. It has been postulated that autocrine purinergic signaling enhances cyst expansion and accelerates disease progression (Schwiebert et al., 2002). An increase in expression of P2Y2, P2Y6, and P2X7 receptors has been reported in cystic tissue from the Han:SPRD cy/+ rat model of autosomal dominant polycystic kidney disease (Turner et al., 2004a). P2 antagonists and inhibitors of ATP release are being explored as therapeutic agents to treat this disease. A recent article suggests that ATP may inhibit pathological renal cyst growth through P2X7 signaling (Hillman et al., 2004). There is increased glomerular expression of P2X7 receptors in two rat models of glomerular injury due to diabetes and hypertension (Vonend et al., 2004). A further study of human and experimental glomerulonephritis also shows increase in P2X7 receptor expression (Turner et al., 2004b).
Administration of ATP complexed with MgCl2 has been used for many years to improve postischemic and drug-induced glomerular and tubular function (Sumpio et al., 1984, 1985). There is convincing evidence that there is increased sympathetic activity in renal disease, especially ischemia (Joles and Koomans, 2004). Because ATP is established as a cotransmitter with NA in sympathetic nerves, this may be a source of enhanced ATP in these conditions.
The potential role of P2X3 receptors in mechanosensory transduction has already been mentioned in relation to the bladder (Burnstock, 2001e). However, there is increasing evidence that this is not an isolated phenomenon and that ATP released from the epithelial lining of other organs including the ureter following distension may act on P2X3 receptors on afferent nerves in the subepithelial plexuses to provide sensory feedback and, in the case of the ureter, renal colic pain (see Fig. 2 and section II.B.). P2X3 receptors have been found on the suburothelial nerve plexus, and both the human and guinea pig ureter and urothelial cells have been shown to release ATP in a pressure-dependent fashion when the ureter is distended (Knight et al., 2002). This ATP release is abolished when the urothelium is removed, and sensory nerve-recording studies during ureteral distension demonstrate purinergic involvement, suggesting that specific P2X3 antagonists may have efficacy in alleviating renal colic (Rong and Burnstock, 2004).
B. Lower Urinary Tract
In the normal human bladder, atropine will block at least 95% of parasympathetic nerve-mediated contraction, indicating that its innervation is predominantly cholinergic; purinergic signaling is responsible for the atropine-resistant component of contraction (Burnstock, 2001e). There are a number of examples of the purinergic component of cotransmission increasing in pathological conditions (Abbracchio and Burnstock, 1998; Burnstock, 2001e). One is that purinergic nerve-mediated contraction of the human bladder is increased to 40% in pathophysiological conditions such as interstitial cystitis, outflow obstruction, idiopathic detrusor instability, and probably also neurogenic bladder (Wammack et al., 1995; Andersson, 1997; Andersson and Hedlund, 2002). ATP release from bladder epithelial cells from patients with interstitial cystitis is significantly greater than that from healthy cells (Sun and Chai, 2002), and there is a change in expression of both P2X and P2Y receptors in urothelial cells (Birder et al., 2004; Tempest et al., 2004). P2X1 receptor subtype expression markedly increased in obstructed bladder (Boselli et al., 2001) and in the absence of P2X3 receptors in mouse knockouts, the bladder is hyperactive (Cockayne et al., 2000; Vlaskovska et al., 2001). Botulinum toxin A, which has antinociceptive effects in treating interstitial cystitis, inhibits distension-mediated urothelial release of ATP in conditions of bladder inflammation (Smith et al., 2004).
Purinergic signaling also plays a role in afferent sensation from the bladder (Wyndaele and De Wachter, 2003). ATP is released from urothelial cells when the bladder is distended (Ferguson et al., 1997; Vlaskovska et al., 2001). Sensory nerve recording has indicated that P2X3 receptors are involved in mediating the nerve responses to bladder distension, providing mechanosensory feedback involving both the micturition reflex and pain (Cockayne et al., 2000; Burnstock, 2001c; Vlaskovska et al., 2001; Rong et al., 2002). Purinergic agonists acting on P2X3 receptors in the bladder can sensitize bladder afferent nerves, and these effects mimic the sensitizing effect of cystitis induced by cyclophosphamide (Yu and de Groat, 2004). Thus, P2X3 receptors are a potential target for pharmacological manipulation in the treatment of both pain and detrusor instability.
In aging rats there is increased sensitivity of the bladder to ATP, but no change in response to acetylcholine or potassium chloride (Ferguson and Christopher, 1996). Comparable results showing increased purinergic, but not cholinergic, neurotransmission to aging human bladder smooth muscle have been found (Yoshida et al., 2001). Activation of P2 receptors in the brain stem (both periaqueductal gray matter and Barrington's nucleus/locus ceruleus) generates patterns of activity in the parasympathetic innervation of the bladder (Rocha et al., 2001). In patients with idiopathic detrusor instability, there is abnormal purinergic transmission in the bladder; this may account for some of the symptoms and provide a novel therapeutic target for treatment of overactive bladder (Andersson, 2002; O'Reilly et al., 2002). Voiding dysfunction involves P2X3 receptors in conscious chronic spinal cord-injured rats, which raises the possibility that P2X3 antagonists might be useful for the treatment of neurogenic bladder dysfunction (Lu et al., 2002). Drugs that alter ATP release or breakdown might also be therapeutic targets (Harvey et al., 2002; Chess-Williams, 2004). Copper inhibits purinergic transmission in the bladder and the copper(I) chelator, neocuproine, enhances bladder activity by facilitating purinergic excitatory responses (Göçmen et al., 2005). Expression of P2 receptors in bladder urothelium, as well as in nerve and smooth muscle, changes in a cat model of interstitial cystitis and may be linked with painful bladder symptoms (Birder et al., 2004). Recent reviews of management of detrusor dysfunction highlight the potential of therapeutic strategies related to purinergic signaling (Andersson, 2002; Kumar et al., 2003; Fry et al., 2004; Moreland et al., 2004; Nishiguchi et al., 2004; Ouslander, 2004; Yoshida et al., 2004; Birder, 2005; Rapp et al., 2005).
C. Reproductive System
Normal penile erectile function is dependent upon a delicate balance between contracting and relaxing factors in the corpus cavernosum smooth muscle, which are modulated by signaling from both nerves and endothelial cells. Evidence has accumulated to support a pivotal role for NANC neurotransmitters. NO plays a central role in mediating cavernosal smooth muscle relaxation, but other neurotransmitters can modulate this action and may play a role in erectile dysfunction. ATP potently relaxes cavernosal smooth muscle strips in vitro, an action pharmacologically consistent with P2Y receptors. Indeed, P2Y receptors are present on both cavernosal smooth muscle cells and endothelial cells, and ATP is released from a subpopulation of the cavernosal nerves. It appears that smooth muscle relaxation is caused by ATP acting directly on the cavernosal smooth muscle cells and indirectly is mediated by NO released from the endothelial cells. ATP-mediated cavernosal relaxation is impaired in diabetes mellitus (independent of NO), implying that purinergic signaling may be involved in the pathophysiology of erectile dysfunction (Gür and Öztürk, 2000).
In humans, ATP induces a significant increase in sperm fertilizing potential and this provides a rationale for the use of ATP for treatment of spermatozoa during in vitro fertilization (Rossato et al., 1999). Knockout mice lacking P2X1 receptors appear normal, but fail to breed, and this is associated with loss of the purinergic component of sympathetic cotransmission in the vas deferens; these findings raise the possibility of developing nonhormonal ways of regulating male fertility (Dunn, 2000). Differential, stage-dependent immunostaining for P2X receptors during spermatogenesis in the adult rat testes has been described (Glass et al., 2001) and opens up the possibility of purinergic targets for both fertility and contraception. Evidence has been presented that glycolysis has an unexpectedly important role in providing the ATP required for sperm motility throughout the length of the sperm flagellum (Mukai and Okuno, 2004).
Synergistic effects of ATP and oxytocin in increasing [Ca2+]i in mouse mammary myoepithelial cells suggest that activation of purinergic receptors may facilitate myoepithelial cell contraction in the milk ejection response (Nakano et al., 2001). P2Y2 receptors on apical and basolateral membranes appear to be involved (Blaug et al., 2003).
Micromolar concentrations of ATP stimulate a biphasic change in transepithelial conductance in the human uterine cervix, with phase 1 mediated by the P2Y2 receptor and phase II by the P2X4 receptor (Gorodeski, 2002). Given the potential role of ATP regulation of cervical paracellular permeability for human fertility, contraception, and health, these findings may have clinical significance and may lead to the development of drugs that can target specific signaling pathways in the cervix.
VIII. Dermatology
Purinergic signaling is much involved in keratinocyte turnover in skin epidermis: P2Y1 and P2Y2 receptors in basal and parabasal layers mediate cell proliferation; P2X5 receptors in the granular layer mediate cell differentiation; and P2X7 receptors at the stratum granulosum/stratum corneum border mediate apoptosis (Greig et al., 2003c). There are changes in the expression of P2 receptor subtypes in proliferative disorders of the epidermis, including psoriasis and scleroderma (Greig and Burnstock, unpublished data), and P2Y2 receptors have been proposed as a novel target for therapy of these disorders (Dixon et al., 1999). P2Y1, P2Y2, and P2Y4 receptors have also been identified on cultured keratinocytes (Cook et al., 1995) and on the HaCaT cell line (Burrell et al., 2003).
There is an increase of P2X3 and P2X2/3 nociceptive receptors on sensory nerve endings in inflamed skin (Hamilton et al., 2001), and antagonists are being developed as analgesics (see section II.B.). Data have been presented to support a pathogenic role for keratinocyte-derived ATP in irritant dermatitis (Mizumoto et al., 2003).
Changes in expression of purinergic receptors in the regenerating epidermis in wound healing have been described (Greig et al., 2003a). The P2X receptor antagonist, PPADS, has been shown to accelerate skin barrier repair and prevent epidermal hyperplasia induced by skin barrier disruption (Denda et al., 2002).
P2 agonists have significant immunostimulating effects on Langerhans cells, a subclass of dendritic cells in the epidermis (Granstein et al., 2005). The authors suggest that ATP, when released after trauma and infection, may act as an endogenous adjuvant to enhance the immune response and that P2 agonists may augment the efficacy of vaccines. Purinergic signaling involved in skin tumors is discussed in section XI.
IX. Respiratory Diseases
P2 receptor purinergic compounds are being explored for the treatment of cystic fibrosis, to improve the clearance of secretions from the bronchi in chronic obstructive pulmonary disease, and for sputum expectoration in smokers (Gaba et al., 1986; Stutts and Boucher, 1999; Taylor et al., 1999; Mall et al., 2000; Yerxa, 2001). ATP and UTP stimulate P2Y2 receptor-mediated surfactant secretion and transepithelial chloride secretion in type II alveolar cells; there are abnormalities in this mechanism in cystic fibrosis (Yerxa, 2001; Bucheimer and Linden, 2004). Nucleotides also increase mucus secretion from goblet cells and increase the ciliary beat frequency of airway epithelial cells (Kemp et al., 2004). The long-lasting P2Y2 receptor analog, INS 37217, increases the duration of mucociliary clearance and therefore has significant advantages over other P2Y2 agonists for the treatment of cystic fibrosis (Kellerman et al., 2002; Yerxa et al., 2002). P2X4 receptors have also been identified on lung epithelial cells and appear to be involved in regulation of ciliary beat, manipulation of which may also be of therapeutic benefit for cystic fibrosis (Zsembery et al., 2003). Recent evidence supports the view that vagal afferent purinergic signaling may be involved in the hyperactivity associated with asthma and chronic obstructive pulmonary disease (Adriaensen and Timmermans, 2004; Undem et al., 2004).
The use of theophylline, an adenosine-receptor antagonist, as an antiasthmatic agent has focused attention on the development of novel P1 receptor antagonists as asthmatic medications (Feoktistov and Biaggioni, 1996; Meade et al., 2001; Fozard and McCarthy, 2002; Polosa et al., 2002; Blackburn, 2003; Lee et al., 2003; Livingston et al., 2004; Zablocki et al., 2005). It has also been suggested that adenosine cooperates with inflammatory cytokines to stimulate mucin production in the asthmatic airway (McNamara et al., 2004).
Erythromycin is a widely used antibiotic for the treatment of upper and lower respiratory tract infections. One of the most conspicuous effects of erythromycin is the suppression of fluid secretion from bronchial epithelial cells in the treatment of bronchitis. Erythromycin has been shown to block the P2X receptor-mediated Ca2+ influx and may represent one mechanism by which it exerts its antisecretory effects in the treatment of chronic respirator tract infections (Zhao et al., 2000).
The ventrolateral medulla contains a network of respiratory neurons that are responsible for the generation and shaping of respiratory rhythm; it also functions as a chemoreceptive area mediating the ventilating response to hypercapnia. Evidence has been presented that ATP acting on P2X2 receptors expressed in ventrolateral medulla neurons influences these functions (Gourine et al., 2003). A potentially important role for P2 receptor synaptic signaling in respiratory motor control is suggested by the multiple physiological effects of ATP in hypoglossal activity associated with the presence of P2X2, P2X4, and P2X6 receptor mRNA in nucleus ambiguous and the hypoglossal nucleus (Collo et al., 1996; Funk et al., 1997) and microinjection of ATP into the caudal NTS of awake rats produces respiratory responses (Antunes et al., 2005).
The pneumovirus respiratory syncytial virus is the most common cause of lower respiratory tract disease in infants and children. It has a detrimental inhibitory effect on alveolar clearance, an effect that appears to be mediated by UTP, perhaps released by the bronchoalveolar epithelium in response to infection (Davis et al., 2004). This suggests that P2Y2 receptor antagonists may be useful for the treatment of severe respiratory syncytial virus broncheolitis.
Alveolar macrophages play a pivotal role in the development of chronic lung inflammatory reactions such as idiopathic pulmonary fibrosis, silicosis, asbestosis, hypersensitivity pneumonitis, sarcoidosis, and mycobacteria tuberculosis. P2X7 receptors are expressed in alveolar macrophages, which upon stimulation activate the proinflammatory IL-1 to IL-5 cytokine cascade and the formation of multinucleated giant cells, a hallmark of granulomatous reactions (Lemaire and Leduc, 2004). P2X7 receptors may be a relevant target for therapeutic intervention in lung hypersensitivity reactions associated with chronic inflammatory responses.
The need to support the failing lung (acute respiratory distress syndrome) with mechanical ventilation is potentially lifesaving, but, unfortunately, alveolar overdistension and pulmonary shear stress may cause lung injury (ventilator-induced lung injury), increasing bronchoalveolar lavage and leading to lung edema. It has been suggested that ventilator-induced lung injury may involve stretch-associated release of ATP from neuroepithelial cell bodies (Brouns et al., 2000, 2003; Rich et al., 2003) and may therefore be a therapeutic target for this condition.
The protective effect of ATP-MgCl2 in ischemia-reperfusion lung injury appears to require the presence of leukocytes (Chen et al., 2003).
X. Musculoskeletal Diseases
Several reports implicate purinergic signaling in bone development and remodeling (Bowler et al., 1999; Elfervig et al., 2001; Hoebertz et al., 2003; Costessi et al., 2005). P2X and P2Y receptors are present on osteoclasts, osteoblasts, and chondrocytes (Fig. 3). ATP, but not adenosine, stimulates the formation of osteoclasts and their resorptive actions in vitro and can inhibit osteoblast-dependent bone formation. The bisphosphonate clodronate, which is used in the treatment of Paget's disease and tumor-induced osteolysis, may act through osteoclast P2 receptors. Very low (nanomolar) concentrations of ADP, acting through P2Y1 receptors, turn on osteoclast activity (Hoebertz et al., 2001).

Schematic diagram illustrating the potential roles played by extracellular nucleotides and P2 receptors in modulating bone cell function. ATP, released from osteoclasts (e.g., through shear stress or constitutively) or from other sources, can be degraded to ADP or converted into UTP via ecto-nucleotidases (1). All three nucleotides can act separately on specific P2 receptor subtypes, as indicated by the color coding. ATP is a universal agonist, whereas UTP is only active at the P2Y2 receptor and ADP is only active at the P2Y1 receptor. ADP via P2Y1 receptors appears to stimulate both the formation (i.e., fusion) of osteoclasts from hematopoietic precursors and the resorptive activity of mature osteoclasts. For the latter, a synergistic action of ATP and protons has been proposed via the P2X2 receptor. ADP could also stimulate resorption indirectly through actions on osteoclasts, which in turn release proresorptive factors [e.g., receptor activator of nuclear factor-κB ligand (RANKL)]. ATP at high concentrations might facilitate fusion of osteoclast progenitors through P2X7 receptor pore formation or induce cell death of mature osteoclasts via P2X7 receptors. In osteoblasts, ATP, via P2X5 receptors, might enhance proliferation and/or differentiation. By contrast, UTP, via P2Y2 receptors, is a strong inhibitor of bone formation by osteoblasts. For some receptors (e.g., P2X4 and P2Y2 receptors on osteoclasts or P2X2 receptors on osteoblasts), evidence for expression has been found, but their role is still unclear (question marks). Dashed lines indicate signaling events in the cell. Reprinted from Hoebertz et al. (2004), with permission from Elsevier.
In a recent study, deletion of the P2X7 receptor revealed its regulatory roles in bone formation and resorption (Ke et al., 2003). It reduces bone resorption by decreasing osteoclast survival (Korcok et al., 2004), and P2X7 receptors are expressed in a subpopulation of osteoblasts (Gartland et al., 2001). The multiple purinoceptors on bone and cartilage also represent potential targets for the development of novel therapeutics to inhibit bone resorption in diseases such as rheumatoid arthritis, osteoporosis, tumor-induced osteolysis, and periodontitis (Dixon and Sims, 2000; Komarova et al., 2001; Naemsch et al., 2001).
Nucleotide metabolism is tightly controlled in the cartilage extracellular matrix. Apart from modulating purinoceptor activation, ectoenzymes can regulate the levels of extracellular phosphate and pyrophosphate, the components involved in crystal deposition (Graff et al., 2003). Mechanical stimulation, which is critical for the maintenance of healthy articular cartilage, can influence the rate of nucleotide release and metabolism. The repair of fetal, but not adult, articular cartilage involves the intercellular transfer of ATP, an increase of [Ca2+]i, and expression of c-fos in cartilage (Kumahashi et al., 2004).
Tendinosis is a disorder characterized by acute or chronic pain and degenerative changes in the matrix. Mechanical loading induces ATP release from tendon cells and stimulates expression of IL-1β, COX-2, and metalloproteinases as a negative feedback mechanism to limit activation of the injurious pathway. Attenuation of the feedback mechanism may result in the progression of tendinosis (Tsuzaki et al., 2003).
Ultrasound is often used to accelerate fracture healing. It has been shown recently that osteoblasts respond to ultrasound stimulation by increasing ATP release, which appears to mediate stimulation of osteoblast gene expression and cell proliferation (Hayton et al., 2005).
A role for purinergic signaling in rheumatic diseases has been considered for some time. Quinacrine (Atabrine), a drug that binds strongly to ATP, has been used for the treatment of patients with rheumatoid arthritis for many years (Wallace, 1989). One of its mechanisms of action is to decrease levels of prostaglandin E2 and COX-2, which are known to be produced following occupation of P2Y receptors by ATP (Needleman et al., 1974; Brambilla et al., 1999). The articular fluid removed from arthritic joints contains high levels of ATP (Ryan et al., 1991). Purinergic regulation of bradykinin-induced plasma extravasation and adjuvant-induced arthritis has been reported (Green et al., 1991). ATP and UTP activate calcium-mobilizing P2Y2 or P2Y4 receptors and act synergistically with IL-1 to stimulate prostaglandin E2 release from human rheumatoid synovial cells (Loredo and Benton, 1998). When monoarthritis was induced by the injection of complete Freund's adjuvant into the unilateral temporomandibular joint of the rat, the pain produced was associated with an increase in P2X3 receptor-positive small neurons in the trigeminal ganglion (Shinoda et al., 2005). Relief of inflammatory pain by the P2X7 receptor antagonist, oxidized ATP, in arthritic rats has been reported (Dell'Antonio et al., 2002a,b). Spinal P1 receptor activation has been claimed to inhibit inflammation and joint destruction in rat adjuvant-induced arthritis, supporting the view that therapeutic strategies, which target the CNS, might be useful in arthritis (Boyle et al., 2002; Sorkin et al., 2003). Suppression of experimental zymosan-induced arthritis by intraperitoneal administration of adenosine has also been described (Baharav et al., 2002).
Lymphoblastoid cells isolated from patients with Duchenne muscular dystrophy are highly sensitive to stimulation by extracellular ATP (Ferrari et al., 1994). A recent study provides the first evidence for a role for purinergic signaling in muscle regeneration using the mdx mouse model of muscular dystrophy and raises the possibility of new therapeutic strategies for the treatment of muscle disease (Ryten et al., 2004). Pain related to the musculoskeletal system (myofascial pain) is very common, and ATP has been claimed to excite or sensitize myofascial nociceptors (Kennedy and Leff, 1995; Bland-Ward and Humphrey, 1997; Mork et al., 2003).
XI. Oncology
The anticancer activity of adenine nucleotides was first described by Rapaport in 1983. Intraperitoneal injection of ATP into tumor-bearing mice resulted in significant anticancer activity against several fast-growing aggressive carcinomas (Agteresch et al., 2003). ATP inhibits the growth of murine colonic adenocarcinoma and human pancreatic carcinoma in mice as well as inhibiting the associated weight loss. In a recent comprehensive review about the use of ATP for the treatment of advanced cancer (Abraham et al., 2003), evidence was presented that 1) extracellular ATP inhibits the growth of a variety of human tumors, including prostate, breast, colon, liver, ovarian, colorectal, esophageal, and melanoma cancer cells, partly by mediating apoptotic cancer cell death; 2) ATP administration induces resistance of nonmalignant tissue to chemo- and radiation therapy, and 3) ATP has pronounced anticachexia effects, particularly in older patients, reducing weight loss, anorexia, and hormonal aberrations, largely via its ability to expand blood plasma ATP pools. It was concluded that preclinical data support utilization of ATP in the treatment of advanced cancers confirmed by phase I and II human trials indicating that ATP has a future place as a useful anticancer agent. The combination of ATP administration with other anticancer modalities is beginning to be explored.
Growth of prostate cancer cells in vitro is inhibited by up to 90% by ATP via P2 receptors, although it is not yet clear which subtype mediates this effect and whether it is a directly antiproliferative effect or a proapoptotic effect (Janssens and Boeynaems, 2001; Vanoverberghe et al., 2003; Calvert et al., 2004; Chen et al., 2004). Phase 1 clinical trials have shown that ATP infusion in patients with advanced cancer is feasible, but it is limited by chest tightness and dyspnea, probably due to conversion to adenosine (Agteresch et al., 1999). P2Y2 receptors are expressed in human colon carcinoma cells (Höpfner et al., 1998; Nylund et al., 2004).
A phase II trial has been carried out in patients with non-small-cell lung cancer, showing that intravenous ATP administered for 96 h at 4-week intervals reduced weight loss and improved muscle strength and quality of life (Agteresch et al., 2000; Dagnelie and Agteresch, 2004), as well as inducing cancer cell death (Agteresch et al., 2003). The combined use of ATP and the antitumor agent, etoposide, appears to improve the therapeutic index in human lung carcinoma (Hatta et al., 2004). Exposure of human lung cancer cells to 8-chloroadenosine, a potent therapeutic agent, induces mitotic catastrophe (Zhang et al., 2004a).
A combination of interferon-γ and ATP is being explored for the treatment of acute myeloid leukemia. ATP induced irreversible damage of leukemic cells without injuring normal hemopoietic stem cells, and it was suggested that it could be useful for purging the residual leukemic cells in autologous bone marrow transplantation (Hatta et al., 1994). Extracellular ATP suppresses proliferation and induction of differentiation of human HL-60 leukemia cells, partly mediated by adenosine (after breakdown of ATP) and partly by ATP (Conigrave et al., 2000). P2X7 receptor expression in the evolutive form of chronic β-lymphocyte leukemia has been identified; ATP decreased proliferation of lymphocytes in this form of leukemia (Adinolfi et al., 2002). β-Cell chronic lymphocyte leukemia is unique in showing a 3-fold increased incidence in closely related family members compared with other lymphoproliferative diseases; a candidate gene for this familial incidence is the P2X7 gene (Dao-Ung et al., 2004). Expression of P2X7 receptor mRNA is higher in most types of leukemia, although there is loss of P2X7 receptor function (Zhang et al., 2004c).
Recent studies from our laboratory have analyzed the P2 receptor subtypes that contribute to ATP suppression of malignant melanomas (White et al., 2005a,b) in basal and squamous cell tumors (Greig et al., 2003b) and prostate and bladder cancers (Shabbir et al., 2003, 2004; Calvert et al., 2004). In general, P2Y1 and P2Y2 receptors mediate proliferation or antiproliferation, P2X5 receptors mediate cell differentiation, which in effect is antiproliferative, and P2X7 receptors mediate apoptotic cell death. ATP administration is particularly effective in treating bladder tumors when combined with the more commonly used anticancer drug mitomycin (Shabbir et al., 2004).
Intracarotid administration of ATP selectively increases blood flow in transplanted, malignant gliomas and suggests that this might enhance the delivery of anticancer agents to malignant brain tumors (Baba et al., 1989). P2Y4 receptors have been shown recently to participate in the commitment to differentiation and cell death of human neuroblastoma cells (Cavaliere et al., 2005).
For years, surgeons have washed the abdominal cavity with distilled water to lyse isolated cancer cells left after surgery. A recent study has shown that the hypotonicity produced by the water leads to cell swelling and release of ATP, which then acts on P2X7 receptors on cancer cells, leading to death (Selzner et al., 2004). 17β-Estradiol has been shown to block the apoptosis induced by P2X7 receptor occupation of human cervical cancer cells (Wang et al., 2004b).
XII. Future Developments
Although in its infancy, the clinical manipulation of purinergic signaling has begun. Several clinically relevant pharmacological interventions are already part of day-to-day practice. However, one of the main reasons that we do not yet have more purinergic therapies in our formularies is the current scarcity of receptor subtypespecific agonists and antagonists that are stable and effective in vivo. In addition to the development of selective agonists and antagonists for the different P2 receptor subtypes, therapeutic strategies are likely to include agents that control the expression of P2 receptors, inhibitors of extracellular breakdown of ATP, and enhancers or inhibitors of ATP transport. Investigating the interactions of purinergic signaling with other established signaling systems will also be very important.
Footnotes
-
↵1 Abbreviations: NANC, nonadrenergic, noncholinergic; NO, nitric oxide; E-NTPDase, ectonucleoside triphosphate diphosphohydrolase; NA, noradrenaline; DRG, dorsal root ganglia; PPADS; pyridoxal-5′-phosphate-6-azophenyl-2′,4′ disulfonic acid; TNP, trinitrophenyl; NMDA, N-methyl-d-aspartate; AIT-082, 4-[[3-(1,6-dihydro-6-oxo-9-purin-9-yl)-1-oxopropyl]amino]benzoic acid; LTP, long-term potentiation; NTS, nucleus tractus solitarius; Ap4A, diadenosine tetraphosphate; IL, interleukin; COX, cyclooxygenase; CNS, central nervous system; IB4, isolectin B4; IBD, inflammatory bowel disease; MRS2500, 2-iodo-N6-methyl-(N)-methanocarba-20-deoxyadenosine-30,50-bisphosphate; A-317491, 5-([(3-phenoxybenzyl)[(1S)-1,2,3,4-tetrahydro-1-naphthalenyl]amino]carbonyl)-1,2,4-benzenetricarboxylic acid; INS37217, P1-(uridine 5′)-P4-(2′-deoxycytidine 5′)tetraphosphate, tetrasodium salt.
-
Article, publication date, and citation information can be found at http://pharmrev.aspetjournals.org.
-
doi:10.1124/pr.58.1.5.
- The American Society for Pharmacology and Experimental Therapeutics
References
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
-
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
In this issue





{kind=link}
{kind=link}
{kind=link}
Jump to section
- Article
- Abstract
- I. Introduction
- II. Cardiovascular Diseases
- III. Neurology
- IV. Immune System and Inflammation
- V. Endocrinology, Diabetes, and Obesity
- VI. Gastroenterology
- VII. Urogenital Diseases
- VIII. Dermatology
- IX. Respiratory Diseases
- X. Musculoskeletal Diseases
- XI. Oncology
- XII. Future Developments
- Footnotes
- References
- Figures & Data
- Info & Metrics
- eLetters