

Abstract
Reaction time variability across trials to identical stimuli may arise from both ongoing and transient neural processes occurring before trial onset. These processes were examined with dense-array EEG as humans completed saccades in a “gap” paradigm known to elicit bimodal variability in response times, including separate populations of “express” and regular reaction time saccades. Results indicated that express reaction time trials could be differentiated from regular reaction time trials by (1) pretrial phase synchrony of occipital cortex oscillations in the 8–9 Hz (low alpha) frequency range (lower phase synchrony preceding express trials), (2) subsequent mid- and late-gap period cortical activities across a distributed occipital-parietal network (stronger activations preceding express trials), and (3) posttarget parietal activations locked to response generation (weaker preceding express trials). A post hoc path analysis suggested that the observed cortical activations leading to express saccades are best understood as an interdependent chain of events that affect express saccade production. These results highlight the importance of a distributed posterior cortical network, particularly in right hemisphere, that prepares the saccade system for rapid responding.
Introduction
Multimodal reaction time distributions to identical stimuli may indicate important features of neural processing dynamics. A classic example of such phenomena is the “gap effect” in the saccade literature (Saslow, 1967; Fischer and Boch, 1983; Dorris and Munoz, 1995). Visually guided saccades normally occur between 170 and 230 ms (Fischer et al., 1993) and are initiated by signals from frontal and parietal ocular-motor cortices to brainstem nuclei (Leigh and Zee, 2006). One means of eliciting faster responses is to separate the fixation point and saccadic target by a subsecond blank screen, or “gap.” Under such conditions, primates, including humans, have a bimodal latency distribution, including a population of “express saccades” with a mode occurring 90–120 ms after target appearance, and a second population of more regular reaction time saccades (Fischer et al., 1993; Clementz, 1996).
Investigations of the gap effect have revealed important characteristics of superior colliculus (SC) functioning that support rapid responding. For instance, the pattern of tonic activity across the SC ocular-motor map determines saccadic reaction time (Dorris et al., 1997). If SC buildup cell activity is augmented before onset of a visual target (e.g., insertion of a gap), a visually guided saccade can be generated early; otherwise, saccadic responses are associated with motor-cortical signaling and are of normal latency (Edelman and Keller, 1996; Munoz and Fecteau, 2002).
Investigations of cortical mechanisms underlying gap-induced bimodality in latency distributions have primarily involved single-unit recordings of frontal eye fields in monkeys (Dias and Bruce, 1994; Everling and Munoz, 2000) and electroencephalography (EEG) measurements without respect to cortical sources of signals measured at the sensors (Everling et al., 1996). The neurophysiology studies have identified gap period firing in cortical neurons specific to express saccades, and EEG research confirmed in humans the existence of unspecified gap period preparatory processes peculiar to express saccades. It is likely that many cortical locations simultaneously contribute to the gap phenomenon, consistent with there being multiple cortical inputs to brainstem ocular-motor nuclei (Leigh and Zee, 2006). Understanding the distributed nature of presaccadic activities would provide important information regarding how the cerebral cortex, in a few hundred milliseconds, can register a transient cue, prepare sensory processing centers, and prime subcortical nuclei to enable rapid action.
An additional focus of the current study concerns the stochastic nature of saccadic gap effects. Latencies in the gap task depend on experience and context (Paré and Munoz, 1996), but it remains unclear why, even after considerable practice, participants generate express saccades on only a proportion of trials (Fischer and Ramsperger, 1986). Power and/or phase in the alpha band before trial onset may influence visual target detection (Ergenoglu et al., 2004; Busch et al., 2009; Mathewson et al., 2009), and the nature of such occipital cortex activity in the pregap period could help determine saccadic latency variability. The influence of such a mechanism on saccadic latency variability in the gap paradigm has yet to be investigated. The present study used dense-array EEG and analysis of data in source space to investigate these phenomena.
Materials and Methods
Participants
Female participants (N = 15) were recruited through the undergraduate psychology research pool at the University of Georgia (mean age, 19.5 years; SD, 1.2). Subjects were in good physical health, absent of known neurological hard signs, and provided informed consent. Exclusion criteria, which were assessed via individual interviews, included neurological disease, a history of severe head trauma or brain injury, and current or past drug or alcohol abuse. This project was approved by the University of Georgia Institutional Review Board.
Stimuli and procedure
Stimuli (Fig. 1) were presented on a 21 inch flat-surface high-resolution color monitor (60 Hz vertical refresh) situated 100 cm from the participant's nasion. The task and visual stimuli were the same for all trials. To start the trial, a fixation cross within a square (2.5°) was presented at the center of the monitor and remained for 1600 ms. The center stimulus was extinguished and the screen remained blank for 200 ms (gap), at which point a 2.5° gray ring was presented for 1000 ms in one of two locations (±8° from fixation; randomly determined) in the horizontal plane (one-half in each visual field). Participants were instructed to move their eyes to the ring as quickly and accurately as possible. Participants completed 144 saccade trials per session over four sessions.

Stimuli and analysis periods. Results from each time period are presented in the figures referenced at that segment.
Electrophysiological recording and data preprocessing
Data collection.
EEG data were vertex-referenced and recorded using a 256-sensor Geodesic Sensor Net and NetAmps 200 amplifiers (EGI). Individual sensor impedances were kept below 50 kΩ (Ferree et al., 2001). In addition, an electrolyte bridge test was conducted between all pairs of sensors before recording (Tenke and Kayser, 2001), and if there was evidence of bridging, sensors were adjusted until bridging was no longer evident (this was rarely required). Data were sampled at 500 Hz with an analog filter bandpass of 0.1–200 Hz. Sensors located at the outer canthi of each eye and below and above both eyes recorded horizontal and vertical eye movements and eye blinks. After data collection, the three-dimensional locations of sensors were acquired using a photogrammetry rig (EGI).
Eye movement data.
The position data from the horizontal eye sensors for individual trials were visually inspected and scored for saccadic latencies (time from peripheral target appearance to saccade initiation) for each trial as by Dyckman and McDowell (2005) using routines written in MATLAB (The MathWorks). Trials with blinks in the gap and late pregap period (from 350 ms before target appearance until saccade onset), and trials with no saccades, were eliminated (17 per session on average). Additionally, eye movements in the pregap fixation period (500 ms after fixation appearance to fixation disappearance) were quantified and compared between saccade types. Express and regular saccades did not differ in the percentage of trials containing blinks (t(14) = 1.05; p = 0.310; two-tailed), and there was no evidence of horizontal or vertical saccades in this time range. A reaction time histogram was created to determine outlier values and to confirm modes for express and regular saccades in the distributions (see Fig. 2).

Reaction time histogram of all saccades made by all participants. The solid lines indicate express saccade range (90–135 ms), and the dotted lines indicate regular saccade range (145–240 ms).
EEG data.
EEG sensors located at the neck and cheeks were excluded, leaving 211 sensors for data analyses. Raw data were visually inspected offline for bad sensor recordings (BESA 5.1; MEGIS Software). Bad sensors (no more than 5% of sensors for any subject) were interpolated using spherical spline interpolation method of BESA. Cardiac artifacts were eliminated using the Independent Component Analysis module in EEGLAB 6.0 (Delorme and Makeig, 2004). The artifact-free data were then transformed to an average reference.
Data analysis
Bimodality in saccadic latency distributions.
For confirmation of the production of express saccades in all participants, the following steps were taken (for an example, see also Clementz, 1996). For each participant, reaction times within the defined range of 90–240 ms were standardized (493 trials on average), adjusted for skewness using the generalized Box–Cox power transformation (Box and Cox, 1964), and tested for bimodality using the SKUMIX algorithm (MacLean et al., 1976). For tests of bimodality (admixture) within response latency distributions for each participant, F values for three- (unimodal distribution) and five-parameter (bimodal distribution) models generated by SKUMIX were compared using a χ2 test on 1 df. Associated values of p < 0.05 indicated the presence of two modes. After these analyses, individual model parameters (means and SDs for the two modes) were averaged to produce a model of the latency distributions that informed the definitions of express and regular saccades used for subsequent analyses.
Pregap period.
To investigate the possibility that neural activity before trial onset may influence gap onset detection and subsequent saccadic reaction time, the following procedure was used. Single-trial voltage data from 950 ms before to 50 ms after gap onset (500 samples) was multiplied by a 500 point Hanning window (this period was extended 50 ms into gap period to account for attenuated activity at the edge as a consequence of windowing). Single-trial data for each sensor were then Fourier-transformed to 1 Hz resolution using the fast Fourier transform (FFT) algorithm in MATLAB. Single-trial power was obtained by squaring the absolute value of the complex result of the FFT. Intertrial coherence (ITC) (a measure of similarity in phase of oscillatory activity over trials in relation to gap onset) (Busch et al., 2009) was calculated by dividing the complex result of the FFT by its absolute value (the amplitude) for each trial and sensor. These values were then summed across trials within subjects and divided by the corresponding number of trials. Resulting values were the Rayleigh statistic R, which is bound between 0 and 1 (1 indicating perfect phase alignment across trials). Pregap power and ITC of individual sensors were compared between express and regular saccades within subjects.
To explore differences in ITC found between express and regular saccades, trials were segregated into four phase bins (0–90, 90–180, 180–270, 270–360°; mean phase for a subject, 0°) for 8 and 9 Hz activity recorded at Oz (central sensor in significant ITC cluster) in this pretrial period. Percentage of saccades in the express range were calculated for each phase bin for each participant, and these data (deviation scores from mean percentage express saccades) were subject to a 4 (phase bin) × 2 (express, regular) two-way repeated-measures ANOVA.
Gap period analyses.
To investigate neural activities during the 200 ms gap period, EEG data from 150 ms before gap onset to 50 ms after target appearance (400 ms total) were baseline adjusted using the 150 ms pregap period and averaged over trials using BESA 5.1. Both subsequent target directions were averaged together since, before target occurrence at the end of the gap period, subjects did not know direction of the required response. Voltage data were collapsed into bins of 10 ms duration before making saccade-type comparisons.
Response period analyses.
To investigate neural activities specifically related to saccade generation, response-locked data (from 250 ms before to 100 ms after saccade initiation) were baseline-adjusted using the 150 ms pregap period and were averaged over trials using BESA 5.1. As can be seen from Figure 5A, the saccade voltage spike started 15 ms before the peak saccadic response. As a result, voltage data were analyzed using the same procedure as described for the gap period analyses only up to 15 ms before the saccade voltage peak in the response-locked averages.
Statistical criteria.
Paired-samples t tests (n = 15; two-tailed) were conducted separately for each sensor for comparing power, ITC, gap period voltages, and response period voltages between saccade types. For significance thresholding, a clustering method (Forman et al., 1995) was used to take account of the nonindependence of data from adjacent EEG sensors with significance levels determined based on the noise level of the data (estimated from the prestimulus period) (for examples, see Clementz et al., 2008; Krusemark et al., 2008) and Monte Carlo simulations calculated using AlphaSim (Cox, 1996). To maintain the family-wise α < 0.01, individual t tests for a given sensor required at least six neighboring sensors with effects statistically significant at p < 0.035.
Source analyses.
After analyses calculated on voltage data at the sensors, standardized low-resolution brain electromagnetic tomography (sLORETA) (Pascual-Marqui, 2002) was used to estimate brain regions involved in determining the between-saccade-type differences on EEG signals observed in the sensor space data (Clementz et al., 2007; Krusemark et al., 2008). sLORETA is a modification of minimum norm least squares (Hämäläinen and Ilmoniemi, 1994) that uses the standardization of the minimum norm inverse solution to infer high-probability regions of brain activation given the measured EEG data. sLORETA solutions yield pseudostatistics that are not appropriate for determining strength of activity, but they provide accurate information about the regions of activity that can account for the voltage pattern recorded at the sensors (Soufflet and Boeijinga, 2005).
An averaged magnetic resonance (MR) image from the Montreal Neurological Institute (Collins et al., 1994) was used to construct a three-compartment boundary element method (BEM) model (Fuchs et al., 2001) before source localization. The MR images were segmented into skin surface, inside of the skull, and cortex. Standard homogeneous conductivities were assumed for the skin, skull, and brain (0.33, 0.0042, 0.33−1 · m−1). For this BEM model, the average triangle edge lengths were 9 mm for the skin, 7.1 mm for the skull, and 5 mm for the brain compartment. Sensors were mapped to the head surface by first matching the fiducial locations (nasion, left and right preauricular points) from the EEG data collection session to the fiducial locations on the averaged segmented skin surface and then projecting the sensors to the skin surface using a nonlinear least-square fitting procedure. The sLORETA calculations then were obtained for each saccade type for each individual. Differences between saccade types were calculated for each individual and averaged to produce source difference maps in brain space. Response period difference maps were created for left and right targets separately. These source localization procedures were implemented using CURRY (version 5.0; Neuroscan). To determine whether these source estimations represented unique or equivalent cortical generators for the EEG voltage differences recorded at separate time points during a trial, local maxima of each sLORETA solution (mid-gap, late-gap, left target response period, and right target response period) in parietal cortex were identified for each participant and compared using separate one-way repeated-measures ANOVAs for x–z coordinates.
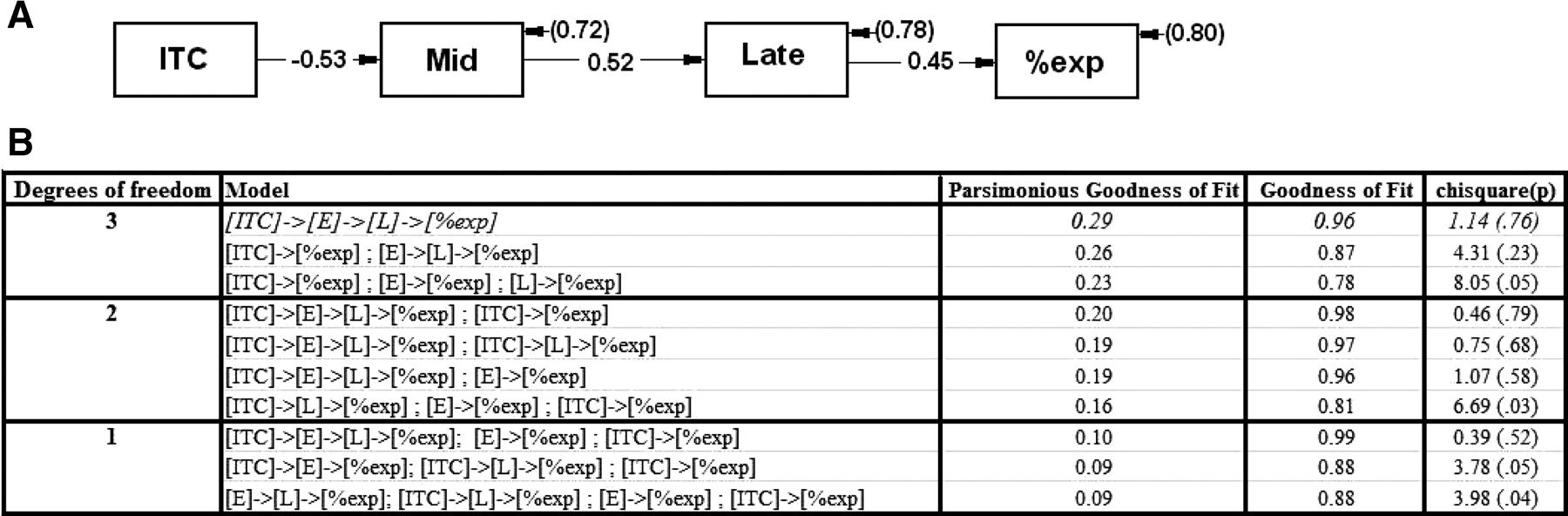
Post hoc path analyses. For each participant, ITC, mid-gap voltage, and late-gap voltage values were averaged across their respective significant clusters to produce single values for a path analysis along with percentage express saccade production (LISREL; Scientific Software International). All temporally plausible path models were analyzed and compared on χ2, and parsimonious goodness-of-fit index (Mulaik et al., 1989) to identify the model that best explained the intervariable relationships.
Results
Behavior
All participants produced saccades with a latency mode in the express range of 90–120 ms as defined in the literature (Fischer and Boch, 1983; Fischer et al., 1993; Clementz, 1996), and most participants (13 of 15) produced response latencies with significantly bimodal distributions. The two subjects without significantly bimodal distributions produced primarily express saccades. A response latency histogram for all saccades for all participants (Fig. 2) confirmed by visual inspection the existence of separate peaks for express and regular saccades. The estimated mean express saccade mode across subjects was 112.8 ms (SD, 12.2), and the estimated mean regular reaction time saccade mode was 164.2 ms (SD, 22.7). Based on the estimated SKUMIX latency mode parameters and the histogram of saccadic response latencies (Fig. 2), and in concordance with previous studies (Fischer et al., 1993; Clementz, 1996), saccade types were defined for all participants: (1) express saccades had latencies between 90 and 135 ms, and (2) regular saccades had latencies between 145 and 240 ms. The following analyses of brain activity are based on this classification.
Brain activity
Pregap period
For express and regular saccade trials, alpha-band (8–12 Hz) power (in square microvolts) was identically distributed across the back of the scalp for occipital and parietal sensors (Fig. 3A) during the pregap period; paired-samples t tests (df = 14, two-tailed) indicated no significant difference in power between saccade types for any alpha-band frequency (p > 0.035 for all sensors). The two saccade types, however, did differ on ITC for both 8 and 9 Hz during this time range (df = 14, two-tailed) (Fig. 3A). Before gap period onset, on trials that would subsequently yield a regular reaction time saccade, participants had significantly larger ITC values than on trials that would subsequently yield an express saccade (averaged across significant sensors for both 8 and 9 Hz ITC). Radial plots of pregap 8 and 9 Hz phase from the significant sensor cluster over posterior cortex (Fig. 3B) indicate that pregap phase was essentially random over trials on which subjects eventually produced an express saccade, but had specific phase characteristics as a function of frequency for trials on which subjects eventually produced a regular reaction time saccade. Subsequent two-way repeated-measures ANOVA results revealed a phase by frequency interaction (F(3,42) = 2.88; p = 0.047), indicating that proportion of express saccades differed between phase bins across frequencies (Fig. 3C).

Pregap period: two-dimensional topographies reveal significant t values in warm colors; 8–9 Hz intertrial phase coherence (with respect to fixation disappearance) is higher for regular saccades (graph represents data from Oz) (A); 8–9 Hz phase is random across express, but not regular saccade trials (B); trials with phase values below mean phase at 8 Hz have higher proportions of express saccades, and trials with phase values above mean phase at 9 Hz have higher proportions of express saccades (C). Error bars reflect within-subject SE (Loftus and Masson, 1994).
Gap period
Paired-samples t tests (df = 14, two-tailed) were used to compare neuralactivities between trials that would subsequently yield express versus regular saccades. There were two time ranges that differentiated groups (Fig. 4A); the voltage topographies within saccade type remained constant within each time range. The first difference occurred from 80 to 110 ms into the gap (mid-gap period) across middle and right parietal and occipital sensors and surrounding FCz (Fig. 4B); the second difference occurred from 170 to 200 ms into the gap (at the very end of the gap; late-gap period) across middle parietal, occipital, and right frontal sensors (Fig. 4C). No other time bins yielded significant between-saccade type differences. For both time periods, trials producing express saccades had greater evoked voltage difference from baseline than trials producing regular reaction time saccades, which is reflected in two positive peaks in sensor Pz (Fig. 4D).
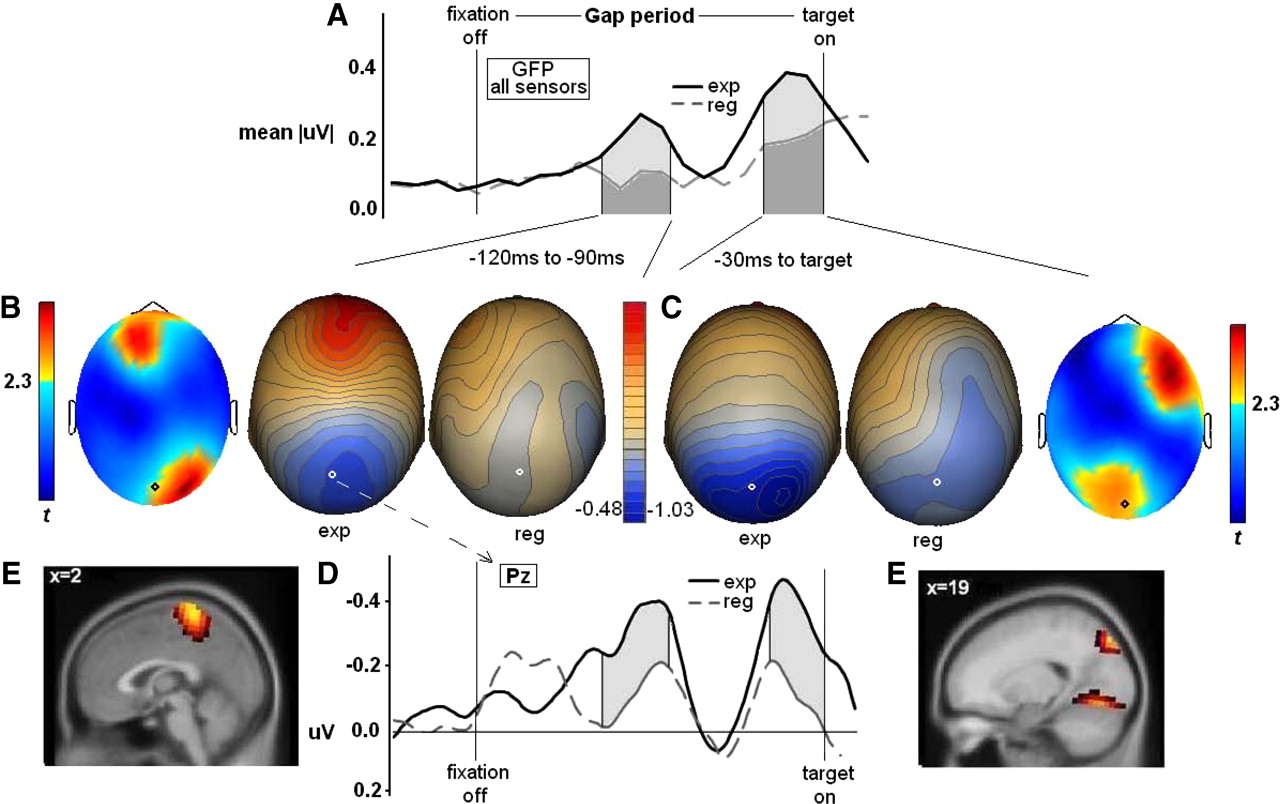
Gap period analyses: time by amplitude plots show global field power over all sensors, indicating two significant voltage peaks (A); two-dimensional topographies reveal significant t values in warm colors for mid-gap (B) and late-gap (C) periods, with corresponding voltage topographies (B, C); event-related potential data from Pz sensor (indicated on heads) (D); express–regular source difference maps (F-distributed) on the MNI brain thresholded at >2 SDs above the mean voxel strength for each time period (E).
Source analyses indicated that neural activations in specific brain regions differentiated between saccade types (Fig. 4E). For the mid-gap period, trials on which there would be express saccades had higher activity in superior dorsal medial parietal lobe (Brodmann's area 7). In late-gap period, neural activation in two distinct brain regions distinguished trials leading to express versus regular saccades: (1) the right posterior cortex at the junction of parietal and occipital cortices (at the junction of Brodmann's areas 7 and 19) and (2) bilateral calcarine fissure in primary visual cortex (Brodmann's area 17). In both instances, express saccade trials had greater cortical activity than regular saccade trials.
Response period
Paired-samples t tests (df = 14; two-tailed) (Fig. 5B) were used to compare neural activities between express and regular saccade trials for the period preceding saccade onset. Mid-central, parietal, occipital, and right temporal and right frontal sensor voltages during the time period immediately before saccade initiation (from 45 to 15 ms before onset of the response) (Fig. 5A,B) significantly differentiated saccade types for both leftward and rightward responses. The voltage topographies within saccade type remained constant within this time range and had marginally different distributions for left and right targets (topography combined for left and right targets for simplicity of presentation) (Fig. 5B). No other time ranges yielded significant between-saccade type differences. Cortical activity immediately preceding regular reaction time saccades was stronger than immediately preceding express saccades, which is reflected in a negative peak in CPz (Fig. 5C). Source analyses indicated that neural activations in Brodmann's area 7 (regular-express differences plotted in Fig. 5D) differentiated saccade types as a function of saccade direction. In response to left targets, regular saccades showed increased activity in right parietal cortex, and for right targets, regular saccades showed increased activity in left parietal cortex.

Response period: time by amplitude plots show global field power over all sensors, indicating a significant voltage peak before regular saccades (beginning of saccade spike is at far right) (A); two-dimensional topographies reveal significant t values in warm colors for this time period; corresponding voltage topographies (combined for left and right stimuli) (B); event-related potential data from CPz sensor (indicated on heads) (C); express–regular source difference maps on the MNI brain thresholded at >2 SDs above the mean voxel strength for each stimulus location (D).
Source location comparisons
Repeated-measures ANOVAs for the parietal cortical locations of sLORETA solutions indicate that mid-gap, late-gap, left target response period, and right target response period signals originate from spatially distinct locations in terms of lateral (x-coordinate, F(3,42) = 3.29, p = 0.030) and anterior–posterior distributions (y-coordinate, F(3,42) = 3.56, p = 022), but not dorsal–ventral distribution (z-coordinate, F(3,42) = 2.49, p = 0.073). Simple comparisons (paired-samples t tests, df = 14, two-tailed, 12 comparisons) revealed that laterality differences stem from the right target response period source in left hemisphere differing from all other activations. The anterior–posterior differences stem from the late-gap source being distinct from all other sources (the late-gap source is more caudal). The mid-gap and left-target response locked sources did not differ significantly in either laterality or anterior–posterior location (supplemental Fig. 1, available at www.jneurosci.org as supplemental material).
Post hoc path analyses
Of all path models evaluated (i.e., those fitting a logical temporal sequence with df > 0) (Fig. 6), the best-fitting and most parsimonious model (χ2 = 1.14; p = 0.760; parsimony goodness-of-fit index = 0.29) (Mulaik et al., 1989) had the form {pregap ITC} → {mid-gap EEG} → {late-gap EEG} → [%express saccades]. The only other model that approached the same combination of good fit and high parsimony included an additional path {pregap ITC} → [%express saccades]. These best-fitting models were also significantly better than a simple multiple regression model for predicting percentage express saccades; including response-locked EEG significantly worsened all model fits.

Post hoc path analyses of participant scores for intertrial phase coherence across occipital sensors [ITC], early/mid-gap peak amplitude [E], late-gap period peak amplitude [L], and proportion of saccades in the express range [%exp]. A, The displayed path was the best fitting and most parsimonious model. Estimated correlation coefficients are embedded in paths, and disturbances are in parentheses. B, Also displayed are results associated with the next best fitting and parsimonious candidate path models (truncated from 20); they are organized by degrees of freedom and parsimonious goodness-of-fit index. The arrows indicate paths.
Discussion
Four aspects of cortical activity differentiated express from regular latency saccades. Neural activities associated with saccade generation were in parietal-occipital cortical regions, highlighting the importance of a posterior cortical network, particularly in right hemisphere (Luna et al., 1998; Corbetta and Shulman, 2002), for preparing the saccade system for rapid responding. This obviously does not diminish the importance of subcortical (Edelman and Keller, 1996; Dorris et al., 1997; Munoz and Fecteau, 2002) and frontal cortical (Dias and Bruce, 1994; Everling and Munoz, 2000) brain regions for saccade generation generally, and express saccades particularly. In what follows, the distinct cortical activations will be discussed separately and then integrated to illustrate a coherent model associated with saccadic gap effects.
Pretrial preparatory activity
Pretrial oscillatory phase at 8–9 Hz over occipital cortex was more aligned to fixation disappearance (trial onset) before regular than express saccades. This could indicate that (1) a particular alpha-band phase window interferes with gap onset detection and, thus, with initiation of gap period brain processes necessary for express saccade production, and/or (2) express saccades result when occipital activity is less organized with respect to a pretrial fixation point.
The former possibility is consistent with research indicating neural oscillations mark rhythmic changes in cortical excitability (Ergenoglu et al., 2004; Jin et al., 2006; Busch et al., 2009). The current finding is also consistent with reports (Mathewson et al., 2009) indicating a potent effect of pretrial alpha-band phase, especially in the low-alpha range (Busch et al., 2009), on performance in visual tasks. That ITC was higher on regular saccade trials is consistent with the account by Mathewson et al. (2009) of alpha as “pulsed inhibition” (i.e., alpha oscillations in particular phase windows diminish probability of target detection). The latter possibility is also consistent with associations between visual attention and stimulus-evoked neuronal entrainment (Schroeder and Lakatos, 2009). In the present study, neuronal entrainment to fixation onset as measured by ITC may index attentional engagement with the fixation point. Visual fixation is known to directly inhibit saccadic initiation (Munoz and Fecteau, 2002), so increased neural investment in fixation could delay neural activations necessary for saccade generation (Dorris et al., 1997).
Primates generate more express saccades with repeated exposure to gap tasks (Fischer and Ramsperger, 1986), which theoretically results from optimization of response preparation (Dorris and Munoz, 1998) and/or target detection with practice (Fine and Jacobs, 2002). Subjects in the present study altered pretrial alpha-band phase characteristics over sessions, with pretrial alpha-band ITC decreasing (sessions 1–4: 0.104, 0.093, 0.091, 0.079) and percentage express saccades increasing (sessions 1–4: 54, 59, 60, 65%). These cross-session changes represent significant linear trends: F(1,28) = 19.719, p < 0.001; and F(1,28) = 30.08, p < 0.001, respectively. This may indicate that a mechanism by which humans speed response times is through early preparation of sensory systems for detection of change. Additional study of practice-induced changes in alpha-band phase may be a promising avenue for investigating behavioral plasticity associated with performance enhancements.
Roles for alpha-band oscillations in express saccade production have been suspected (Kirschfeld et al., 1996; Skrandies and Anagnostou, 1999). The current study did not find differences between preexpress and preregular saccade trials in alpha-band power, so the results are not consistent with the prediction by Kirschfeld et al. (1996) that alpha power at fixation disappearance predicts express saccade probability. Skrandies and Anagnostou (1999) used an analog of alpha-band phase (normalized amplitude), limited their investigation to high-range alpha frequencies (11.72 Hz), did not include pretrial or early-gap periods in their analyses, and found express–regular saccade differences at only one frontal and one parietal sensor. Multiple investigations indicate that the most profound effects of ongoing alpha are at lower alpha frequencies (Busch et al., 2009), before trial onset (Ergenoglu et al., 2004), and over occipital cortex (Capotosto et al., 2009; Mathewson et al., 2009).
Gap period cortical activities
At 80–110 ms into the gap period (mid-gap), superior parietal cortex activity increased on trials preceding express but not regular saccades. This region is associated with visual spatial attention and detection of stimulus spatial change (Corbetta et al., 1993). This neural activity may signal detection of fixation disappearance, a relevant visual-spatial cue before initiation of saccade preparation during gap paradigms.
At 170–200 ms into the gap period (late-gap), there were two cortical activations that differentiated express and regular reaction time saccade trials: primary visual cortex and inferior parietal cortex near the junction of Brodmann's areas 7 and 19 (in the putative location of human parietal eye fields) (McDowell et al., 2008). In both instances, neural activity was higher on trials eventually producing express saccades. A similar pattern has been observed during sustained attention tasks, in which activity levels in specific neural ensembles precede presentation of relevant stimuli (Chawla et al., 1999; Driver and Frith, 2000; Pessoa et al., 2003) and influence subsequent behavioral responses.
The mid- and late-gap activations may prime saccade-generating circuitry in anticipation of target appearance. Visual cortex has access to the brainstem saccade generators through the superior colliculus (Collins et al., 2005). Likewise, neurons in parietal eye fields have access to SC (Lynch et al., 1985), have influence on eye movements through this SC input (Keating et al., 1983), and discharge similarly, but not identically, to SC neurons during interstimulus gap periods (Paré and Wurtz, 2001; Ben Hamed and Duhamel, 2002). Given this circuitry, and the time course of the observed neural activations, these posterior cortex regions may prime brainstem nuclei necessary for express saccade production. This priming could involve the preparatory excitatory modulation of saccadic buildup neurons in SC necessary for express saccade generation (Dorris et al., 1997).
Response-locked activity
From 45 to 15 ms before saccade onset, neural activity in the hemisphere contralateral to the response direction, in the vicinity of precuneus, preceded regular but not express saccades. This same region is consistently activated during a number of saccade tasks (Dyckman et al., 2007; McDowell et al., 2008). The exact contribution of this region to saccadic generation is unclear, but the time course of its activation suggests a role in saccade triggering. This cortical activity marked a later processing stage necessary for the production of regular but not express saccades (which occurred 50 ms earlier on average). This finding supports the hypothesis that preparation for express saccades occurred at an earlier stage, most likely through priming of subcortical nuclei (Edelman and Keller, 1996; Dorris et al., 1997; Munoz and Fecteau, 2002), secondary to the gap period activations in visual and parietal cortices observed here (Fischer et al., 1993).
Cortical network model
The model that best explained the interrelations of ITC, mid-gap (superior parietal cortex) activity, late-gap (parietal eye field and visual cortex) activity, and proportion of express saccades produced was also the most temporally linear. This analysis suggested that the cortical activations leading to express saccades can be understood as occupying a single interdependent chain of events rather than as separate mechanisms that affect express saccade production independently. That is, disengaged (or random) low-frequency alpha-band activity predicts the amount of parietal signaling midway through the gap, which predicts visual cortical and parietal eye field activations later during the gap, which determines express saccade probability. Although this model will require verification in future studies, it converges with theory in many respects. For instance, similar to recent findings by Capotosto et al. (2009), this model maps visual-spatial event detection to the interplay of ongoing and transient occipital and parietal activities.
Similar to Everling et al. (1996) and Everling and Munoz (2000), this model, and the results overall, support the notion that express saccade generation is related to trial-to-trial variations in both ongoing and preparatory activities in cortical regions with access to the brainstem saccade-generating apparatus. Unlike those reports, our findings imply that express saccade-related preparatory activities are in mainly parietal, not frontal, cortices. This seeming incongruence could be associated with multiple factors, including the following: (1) our use of dense-array EEG recording and subsequent source estimation analyses that allowed for inference of location in brain space from where activity recorded at the sensors emanated [the data of Everling et al. (1996) may be consistent with the present findings if the same analyses with a similar sensor montage had been conducted, which was not technically feasible at the time]; and (2) the use by Everling and Munoz (2000) of single-unit recordings in frontal eye field of nonhuman primates, a technology much more sensitive to detecting neural activity regardless of whether it was locked to stimulus occurrence and/or response generation. Everling and Munoz (2000) did not measure activity from posterior brain regions, so the relationship between the frontal cortical signals they measured and the posterior cortical signals observed here is at present uncertain (but see Ruff et al., 2006; Ekstrom et al., 2008).
Nevertheless, the noninvasive methods of the current study for recording neural activity, and analyses of brain signals in the time, space, and frequency domains, provided a demonstration of the posterior cortical generators associated with express saccadic production. Similar to other recent visual processing investigations, these data highlight the importance of ongoing alpha-band phase on behavior and the role of posterior cortical structures in response preparation. It is possible that frontal cortical signals modulate the occipital-parietal preparatory activity observed here by providing an “attentional template” (Desimone and Duncan, 1995), in this case, one that decreases neural investment in central fixation during the pregap preparatory phase. This could be an exciting and important avenue to pursue in subsequent translational research.
Footnotes
-
This work was supported by National Institutes of Health Grants MH076998 and MH082514. We thank Noah Duncan and Emily Leonard Parks for their help with data collection.
- Correspondence should be addressed to Brett A. Clementz, Psychology Department, Psychology Building, Baldwin Street, University of Georgia, Athens, GA 30602. clementz{at}uga.edu





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}