

Abstract
In rat hippocampal cultures, we show by multilabeling immunocytochemistry that pyramidal cells, which receive little or no GABAergic input, mistarget α2-GABAA receptors and gephyrin to glutamatergic terminals. This mismatch does not occur in neurons innervated by numerous GABAergic terminals. A similar phenomenon has been reported for isolated autaptic hippocampal neurons (Rao et al., 2000). GABAergic synapses typically form multiple release sites apposed to GABAA receptor and gephyrin clusters. Remarkably, dystrophin, a protein highly abundant in skeletal muscle membranes, is extensively colocalized with α2-GABAA receptors exclusively opposite GABAergic terminals. In addition, selective apposition of syntrophin and β-dystroglycan to GABAergic presynaptic terminals suggests that the entire dystrophin-associated protein complex (DPC) clusters at GABAergic synapses. In contrast to gephyrin and GABAAreceptors, DPC proteins are not mistargeted to glutamatergic synapses, indicating independent clustering mechanisms. This was confirmed in hippocampal neurons cultured from GABAA receptor γ2 subunit-deficient mice. Clustering of GABAA receptor and gephyrin in these neurons was strongly impaired, whereas clustering of dystrophin and associated proteins was unaffected by the absence of the γ2 subunit. Our results indicate that accumulation of dystrophin and DPC proteins at GABAergic synapses occurs independently of postsynaptic GABAAreceptors and gephyrin. We suggest that selective signaling from GABAergic terminals contributes to postsynaptic clustering of dystrophin.
- GABAergic synapse
- dystrophin
- dystrophin-associated protein complex
- presynaptic signaling
- clustering
- hippocampus
- cell culture
- immunofluorescence
The mechanisms regulating the differentiation and molecular composition of the postsynaptic apparatus are best understood for the neuromuscular junction (NMJ). The tyrosine receptor kinase MuSK plays an essential role in initiating prepatterning of acetylcholine receptors (AChRs) in muscle cells and mediates trans-synaptic signaling of agrin, which is required for proper formation of the NMJ and its association with proteins forming the dystrophin-associated protein complex (DPC) (for review, see Sanes and Lichtman, 1999; Davis et al., 2001; Ferns and Carbonetto, 2001). In the CNS, the postsynaptic density of excitatory synapses is well characterized, containing a complex of >70 identified proteins, including glutamate receptors, scaffolding proteins, and signaling molecules (Husi et al., 2000; Walikonis et al., 2000; for review, see Craig and Boudin, 2001). An immediate-early gene product, Narp, has been proposed as a putative candidate fortrans-synaptic signaling-inducing clustering of AMPA receptors (O'Brien et al., 1999). In contrast, inhibitory synapses are much less well understood. Gephyrin was identified as a scaffolding protein essential for clustering of both glycine and GABAA receptors, and only a few additional proteins have been identified so far (for review, see Kneussel and Betz, 2000; Luscher and Fritschy, 2001; Moss and Smart, 2001). They include dystrophin, which is present in a subset of GABAergic synapses (Knuesel et al., 1999, 2001), raising an interesting analogy to the NMJ.
Apposition of neurotransmitter receptors to the appropriate presynaptic terminals requires trans-synaptic signaling, as shown in several experimental preparations (Kirsch and Betz, 1998; Levi et al., 1998, 1999; Rao et al., 2000). Activity-dependent neurotransmitter release is an attractive candidate for this signal. However, except for glycine receptors (Kirsch and Betz, 1998; Levi et al., 1998), activity blockade or receptor blockade does not affect receptor clustering (Craig et al., 1994; Verderio et al., 1994; Mammen et al., 1997;Cottrell et al., 2000; Rao et al., 2000). Furthermore, there is evidence that presynaptic signaling from neurochemically distinct types of interneurons might determine postsynaptic receptor composition in GABAergic synapses on hippocampal pyramidal cells (Maccaferri et al., 2000; Nyiri et al., 2001). Thus, one can expect such signals to play a role in the recruitment of specific scaffolding and signaling molecules to the postsynaptic apparatus.
Rao et al. (2000) showed mismatched apposition of presynaptic and postsynaptic components in isolated hippocampal neurons grown on permissive microislands. Thus, pyramidal cells clustered gephyrin and GABAA receptors precisely opposite to glutamatergic autapses. This finding shows that GABAA receptor and gephyrin clustering are independent of GABAergic input. It raises important questions, such as whether all proteins of the postsynaptic apparatus are independent of appropriate innervation and how presynaptic terminals recruit appropriate receptors and associated proteins in multi-innervated neurons.
We used hippocampal primary cultures that contain only a few GABAergic interneurons to explore immunocytochemically the influence of GABAergic innervation, visualized with markers for GABAergic presynaptic terminals, on the distribution of different components of the postsynaptic apparatus, including the GABAAreceptor α2 subunit, gephyrin, and dystrophin. We show that gephyrin and dystrophin respond in distinct ways to the presence or absence of a GABAergic presynapse, suggesting independent clustering mechanisms for these two proteins.
MATERIALS AND METHODS
Animals. Rat embryos [embryonic day (E) 17 or E18] were obtained from timed mated pregnant OFA [Icolbm: OFA (SPF)] or Wistar rats (RCC, Füllingsdorf, Switzerland). The γ2 subunit-deficient mice were described previously (Gunther et al., 1995). The animals used here were bred in a C57BL/6 background. Timed matings between γ20/+ mice were set up, and cultures were prepared for each embryo separately at E15 and correlated with genotypes as described (Essrich et al., 1998). All experiments were approved by the cantonal veterinary office of Zurich.
Cell culture. Primary cultures of hippocampal neurons were prepared as described previously (Berninger et al., 1995). Embryos were taken from pregnant dams anesthetized with ether. The hippocampus was dissected on ice and incubated for 15 min at 37°C in PBS, pH 7.4, containing 1 mg/ml bovine serum albumin and 12 μg/ml papain (Sigma, St. Louis, MO). Neurons were subsequently dissociated by gentle trituration with a fire-polished Pasteur pipette and suspended in DMEM containing 10% fetal calf serum (Invitrogen, San Diego, CA). They were then plated on poly-l-lysine (Sigma)-coated 35 mm Petri dishes (Invitrogen) at a density of 1.5 × 10−4 to 2 × 10−4cells/cm2. After 24 hr, the medium was exchanged with a defined, serum-free medium (Brewer and Cotman, 1989;Zafra et al., 1990). Feeder layers, prepared from postnatal day (P) 0 rat cortex and plated on coverslips, were placed upside down above the neurons. Cultures were kept at 37°C in a 5% CO2 humidified incubator.
Immunocytochemistry on cell cultures. All experiments were performed on mature cultures (21–28 d in vitro). To ensure selective detection of GABAA receptors in the cell membrane, living cells were incubated with α2 subunit-specific antibodies raised against extracellular epitopes (for characterization, see Fritschy and Mohler, 1995). The living cultures were incubated for 90 min at room temperature with antibodies (affinity-purified, 1.2 μg/ml) diluted in Ringer's solution (in mm): CaCl2 2, MgCl2 2, glycine 0.001, TTX 0.0005, glucose 30, HEPES 25, KCl 5, NaCl 119, pH 7.4 (Archibald et al., 1998). They were subsequently washed three times for 10 min with Ringer's solution and fixed with methanol for 10 min at −20°. Fixed cultures were rinsed extensively with PBS and incubated for 90 min at room temperature with one of the following primary antibodies in PBS containing 10% normal goat serum (NGS): gephyrin [monoclonal antibody (mAb)7a; Alexis Corporation, San Diego, CA; 1:400]; rabbit anti-glutamic acid decarboxylase (GAD65/67) (Affiniti, Exeter, UK; 1:2000) or GAD65 (mAb, GAD-6; Developmental Studies Hybridoma Bank, University of Iowa, Iowa City, IA; supernatant 1:10); rabbit anti-GAD67 (Alpha Diagnostics, San Antonio, TX; 1:1000); rabbit anti-GABA transporter 1 (GAT-1, DiaSorin, Stillwater, MN; 1:3000); rabbit anti-vesicular inhibitory acid transporter (VIAAT) (Dumoulin et al., 1999) (kindly provided by Dr. B. Gasnier, Paris, France; 1:5000); rabbit anti-vesicular glutamate transporter (vGluT1) (BNP1) (Bellocchio et al., 2000; Takamori et al., 2000) (Synaptic Systems, Goettingen, Germany; 1:10,000); rabbit anti-synapsin I (Molecular Probes, Eugene, OR; 1:300); anti-synaptophysin (mAb, Roche Diagnostics, Rotkreuz, Switzerland; 1:1000); anti-dystrophin, C terminus (mAb, Anawa Trading SA, Wangen, Switzerland; 1:50); anti-β-dystroglycan (DG) (mAb, Novocastra, Newcastle, UK; 1:20); and anti-syntrophin (mAb, pan, provided by Dr. S. C. Froehner, University of Washington, Seattle, WA; 1:20). Cultures were subsequently washed three times for 10 min with PBS and incubated with a mixture of secondary antibodies conjugated to Cy3 or Cy5 (Jackson ImmunoResearch, West Grove, PA; 1:500) or Alexa 488 (Molecular Probes; 1:1000) for 60 min at room temperature in PBS plus 10% NGS. After three washes in PBS, cells were coverslipped in 50% glycerol in 0.2 m sodium bicarbonate buffer, pH 9.2.
Data analysis. All experiments were analyzed by conventional fluorescence microscopy using a high-resolution digital camera (Hamamatsu Orca, Hamamatsu Photonics, Hamamatsu City, Japan) and the OpenLab imaging Software (Improvision, Coventry, UK). Semiquantitative analyses were performed on randomly selected samples of 80–120 segments of dendrites (50 or 100 μm) from at least 12 cells in three independent cultures from E17 and E18 rats (see Fig. 2, Tables 1, 2). Clusters were counted with the OpenLab counting tool. All measurements were expressed as mean ± SD. Statistical analysis was performed using Student's t test. Digital images were processed using the software Imaris (Bitplane, Zurich, Switzerland).
RESULTS
Cultures of hippocampal neurons prepared from E17 or E18 rat embryos, plated at a density of ∼20,000 cells/cm2 and maintained for 21–28 din vitro, were characterized by morphologically mature cells and a high density of synapses, as detected with markers of both glutamatergic and GABAergic presynaptic and postsynaptic elements. Staining for GAD65 revealed a low density of GABAergic interneurons (0.2–5% of total cell number), which form a population of large cells (mean soma diameter, 60 μm) with smooth dendrites and expressing a prominent GABAA receptor α1 subunit-immunoreactivity (IR) (Brünig et al., 2002). However, GAD-positive axons typically formed an extensive plexus of beaded fibers innervating neighboring cells. As a result, only some areas of the culture were covered with GABAergic axons, whereas adjacent regions were devoid of GABAergic innervation. Cultures derived from E18 rats contained on average more interneurons and matured faster than E17 cultures. The neurochemical identity of GAD-positive axons was verified with antibodies against the GABA transporters VIAAT and GAT-1, which produced staining patterns identical to those of GAD and therefore selectively labeled GABAergic axons. In contrast, no specific staining was obtained with antibodies against GAD67 (data not shown), suggesting that this isoform is not expressed in vitro. The inhomogeneous distribution of GABAergic axons in our cultures provided the opportunity to compare the distribution of GABAA receptors and associated postsynaptic proteins, such as gephyrin and dystrophin, in cells receiving strong GABAergic input and in cells receiving little or no GABAergic innervation.
Mismatched apposition of GABAA receptors and gephyrin to glutamatergic terminals
As reported previously (Essrich et al., 1998; Brünig et al., 2001, 2002), staining for the GABAA receptor α2 subunit revealed a clearly punctate immunoreactivity distributed on the soma and dendrites of pyramid-like cells (Fig. 1a). These puncta were extensively colocalized with gephyrin-IR (Fig. 1a,insets), representing postsynaptic clusters. Because a diffuse, presumably extrasynaptic, α2 subunit staining was also seen, clusters were defined by their intensity (more than twice the intensity of the surrounding membrane) and their apparent diameter (>0.3 μm; 100× oil immersion lens, numerical aperture 1.4). Postsynaptic receptor clusters can also be revealed by staining with the β2,3 or γ2 subunit, indicating that they represent functional GABAA receptors (Craig et al., 1994,1996; Essrich et al., 1998; Brünig et al., 2001). In cultures with a low density of GABAergic axons, some α2 subunit and gephyrin clusters were closely apposed to GABAergic terminals, as shown by triple immunostaining with GAD (Fig. 1a), VIAAT, or GAT-1 (data not shown). These synapses were characterized by groups of GABAA receptor and gephyrin clusters surrounding a GABAergic bouton (Fig. 1b), suggesting that each presynaptic terminal formed as many release sites. These clusters represented appropriately matched GABAergic presynaptic and postsynaptic elements.

Formation of appropriately matched and mismatched synapses in the same neuron. a, Triple immunofluorescence staining for the α2 subunit (red), gephyrin (green), and GAD (blue) (see also insets) showing a pyramid-like cell contacted by a single GABAergic axon. Particularly brightly stained α2 and gephyrin clusters are grouped along the trajectory of a GAD-positive axon. In addition, α2 and gephyrin colocalize in many smaller clusters evenly distributed over dendrites (arrowheads), which are not apposed to GABAergic boutons. Diffuse α2 subunit staining of dendrites is attributable to nonclustered, extrasynaptic GABAA receptors. Scale bar, 5 μm. b, Triple staining of the α2 subunit (red), gephyrin (green), and GAD (blue). Each GABAergic bouton is surrounded by multiple α2- and gephyrin-positive clusters, suggesting the presence of as many release sites facing postsynaptic specializations. The α2 and gephyrin clusters that are not apposed to GABAergic terminals (arrows) form single clusters. Scale bars, 2 μm.c, Triple staining of the α2 subunit (red), vGlut1 (green), and GAD (blue) of a cell contacted by a single GABAergic axon. The α2 subunit clusters, which are not apposed to GAD-positive boutons, are apposed to a Glut1-positive, glutamatergic terminals (inset) and represent mismatched synapses. Scale bar, 5 μm. d, Double staining of gephyrin (red) and vGlut1 (green), depicted at high magnification. Again, most gephyrin clusters are apposed to vGlut1-positive boutons. Scale bar, 1 μm.
The remaining α2 subunit and gephyrin clusters not apposed to GABAergic boutons (Fig. 1a,b,arrowheads) corresponded to mismatched synapses apposed to glutamatergic terminals (90.4 ± 10.6%) (Table1), as shown by triple staining for α2, GAD, and the glutamate transporter vGlut1 (Fig. 1c). Gephyrin clusters also were frequently apposed to vGlut1-positive terminals, as expected from their colocalization with the α2 subunit (Fig. 1d). Therefore, mistargeting of gephyrin and GABAA receptors to glutamatergic terminals occurs in hippocampal neurons that receive limited GABAergic input. We have shown previously that gephyrin does not colocalize with postsynaptic density protein 95 (PSD95) or the glutamate receptor GluR1 subunit at postsynaptic sites (Brünig et al., 2002). Thus, mismatch of GABAergic postsynaptic elements to glutamatergic terminals apparently leads to the formation of nonfunctional synapses.
GABAA receptors cluster in both GABAergic and mismatched synapses, whereas the DPC is found exclusively in appropriately matched GABAergic synapses
To determine whether the density of GABAergic input influences the formation of mismatched synapses, the distribution of α2 subunit clusters was compared in cells contacted by low, intermediate, or high numbers of GAD-positive terminals (Fig. 2). In a given culture dish, pyramidal cells receiving no GABAergic input had numerous clusters of α2 subunit-IR evenly distributed over dendrites, presumably targeted to glutamatergic terminals (Fig.2a–c). Cells contacted by only one or a few GABAergic axons formed characteristic groups of α2 subunit clusters along trajectories of GAD-positive fibers (Figs.1a, 2d–f), representing GABAergic synapses. In the remaining dendrites of these cells, numerous α2 subunit clusters were evident, corresponding to mismatched synapses. Finally, in cells contacted by many GABAergic axons, typically surrounding the soma and proximal dendrites, nearly all α2 subunit clusters were apposed to GAD-positive varicosities (Fig. 2g–i). In such cells, there was no evidence of mismatched synapses (Fig. 2i). This was confirmed by triple staining with gephyrin (data not shown), which in these cells was found exclusively in α2 subunit clusters apposed to GAD-positive terminals. A semiquantitative analysis of this effect revealed a negative correlation between the number of mismatched and GABAergic synapses on dendrites (Fig. 2j) (correlation coefficient −0.71; p < 0.001). These results suggest that GABAergic terminals provide an anterograde signal leading to preferential recruitment of GABAAreceptors and gephyrin.

The formation of mismatched synapses depends on the density of GABAergic input, as illustrated for three representative cells found in the same culture dish. a,d, g, α2 subunit staining (red); b, e,h, GAD staining (green);c, f, i, overlay for the boxed areas. a–c, Pyramid-like cell displaying numerous α2 subunit-positive clusters (a) evenly distributed over its dendrites, although it receives no detectable GABAergic input (b). These clusters obviously represent mismatched synapses. d–f, Example of a cell with brightly stained “chain-like” α2clusters (d, arrows) along the GABAergic fiber running over its dendrites (e) representing appropriately matched GABAergic synapses. In addition, this cell displays numerous evenly distributed α2 subunit clusters on its dendrites, corresponding to mismatched synapses.h, i, Pyramid-like cell innervated by numerous GAD-positive axons (h). Clusters of α2 subunit staining (i) are seen only on dendrites contacted by the GABAergic fiber. The overlay (i) shows that every α2 subunit cluster is apposed to a GAD-positive bouton. The remaining staining represents nonclustered, extrasynaptic α2GABAA receptors. j, Number of mismatched synapses as a function of the density of GABAergic synapses on dendrites. Synapses were counted on 50 μm segments of dendrites (n = 64) from 20 cells with variable GABAergic innervation. GABAergic synapses were identified by α2subunit clusters closely apposed to GAD-positive boutons. Isolated α2 subunit clusters were counted as mismatched synapses. Each dot represents one segment. An inverse correlation is evident. Black line, Linear regression. Correlation coefficient, −0.71, p < 0.001. Scale bars: a, b,d, e, g, h, 10 μm; c, f, i, 5 μm.
Dystrophin and the dystrophin-associated protein complex cluster exclusively opposite GABAergic terminals
In rodent hippocampus, cortex, and cerebellum, dystrophin is colocalized with GABAA receptor subunits and gephyrin in presumptive GABAergic synapses (Knuesel et al., 1999). We show here that dystrophin is detectable also in vitro, forming strongly labeled puncta distributed on the soma and dendrites of cultured hippocampal neurons (Figs.3a, 4b). The antibody used is directed against the C terminus of the protein and recognizes both full-length and N-terminally truncated isoforms of dystrophin. Double immunofluorescence staining with GAD confirmed that dystrophin clusters were associated with GABAergic synapses, because they were always apposed to GAD-positive boutons (Fig.3a–c). In many cases, several dystrophin clusters surrounded a GAD-positive bouton, as described for the α2 subunit and gephyrin in GABAergic synapses (Fig. 3c, arrows). The fraction of GAD-positive boutons surrounded by dystrophin clusters was on average 89.9 ± 6%, as measured on dendrites (Table 1). As was the case in vivo, dystrophin-IR was restricted to GABAergic postsynaptic sites (96.3 ± 1.7% of the dystrophin clusters were associated with GAD) (Table 1).

Dystrophin clusters opposite GABAergic terminals. Double immunofluorescence staining of dystrophin (a,red) and GAD (b, green), with overlay (c). Staining of dystrophin (antibody recognizing the C terminus) revealed strongly stained puncta aligned in chains (a). Sometimes groups of clusters were apparent (c, arrows). Costaining with GAD demonstrates that virtually all dystrophin clusters were apposed to GAD-positive terminals. Scale bars, 10 μm.
Double- and triple-labeling studies with dystrophin, the α2 subunit, and GAD revealed that dystrophin clusters were colocalized with GABAA receptors in GABAergic synapses. In cells innervated by a few GAD-positive axons, forming both matched and mismatched synapses, dystrophin-IR was detected only in the groups of clusters typically surrounding GAD-positive boutons (Fig.4a–c). The colocalization of α2 and dystrophin was confirmed in high-magnification images showing that dystrophin clusters were directly apposed to a GAD-positive bouton and were colocalized with the α2 subunit (Fig. 4d–g). Mismatched α2 subunit clusters in the same neuron lacked detectable dystrophin-IR (Fig. 4, arrowheads). In dendrites, only a few appropriately matched GABAergic synapses lacked dystrophin, which was evidenced by the fact that 87.2 ± 10.5% of α2 clusters apposed to GAD were colocalized with dystrophin (Fig. 4, Table 1). Altogether, these results indicate that, in contrast to gephyrin, dystrophin was never mistargeted to glutamatergic terminals. Thus, clustering of dystrophin and of gephyrin is regulated by distinct mechanisms.

Dystrophin is colocalized with the α2 subunit in GABAergic synapses and is not mistargeted to glutamatergic terminals. a–c, Double staining of the α2 subunit (a, red) and dystrophin (b, green), with overlay (c). Dystrophin-positive clusters are colocalized with brightly stained α2 subunit clusters (c, yellow). They appear in chain-like groups, indicating the presence of a GABAergic axon. Many additional α2 subunit clusters, presumably corresponding to mismatched synapses, are not labeled with dystrophin.d–g, Representative examples of triple staining of the α2 subunit (red), dystrophin (green), and GAD (blue).d, f, g, α2and GAD; e, f′, g′, dystrophin and GAD in the same fields. Dystrophin clusters are found only at sites apposed to GABAergic terminals, where they are colocalized with the α2 subunit (d,e, thin arrows). d–g,Arrowheads point to mismatched synapses labeled with the α2 subunit but not with dystrophin. The α2and dystrophin did not colocalize without apposition to a GABAergic terminal. However, occasional α2 subunit clusters in a GABAergic synapse were not matched by dystrophin (f). Scale bars: a–c, 10 μm;d, e, 5 μm.
At the NMJ, dystrophin is associated with a multimeric transmembrane protein complex, the DPC. We have therefore investigated whether other members of the DPC, such as β-dystroglycan and syntrophin, are also located at GABAergic synapses in vitro. Immunofluorescence staining with monoclonal antibodies recognizing β-DG or all three isoforms of syntrophin (α1-, β1-, and β2-syntrophin), respectively, revealed, for both markers, brightly labeled puncta arranged in lines running over cell bodies and dendrites (Fig.5a–l) and apposed to GAD-positive terminals (Fig. 5a–i). Sometimes, as seen for dystrophin, groups of clusters surrounded one bouton (Fig.5d–f) exhibiting the typical morphology for appropriately matched GABAergic synapses. Both β-DG (Fig.5j–l) and syntrophin (data not shown) were colocalized extensively with the α2 subunit. Mismatched synapses did not contain β-DG (Fig. 5l,arrowheads) or syntrophin. The fraction of GABAergic synapses positive for either of these DPC proteins was smaller than for dystrophin, however (73.1 ± 8% of GAD boutons apposed to syntrophin and 74 ± 2% of GAD boutons apposed to β-DG) (Table1). This implies either that only a subpopulation of GABAergic synapses contains dystrophin together with additional DPC proteins or that the antibodies specific for β-DG or syntrophin are less sensitive than those for dystrophin. Conversely, clusters of β-DG-IR or syntrophin-IR were always associated with GABAergic terminals (95.3 ± 4% for β-DG and 94.1 ± 5% for syntrophin) (Fig.5, Table 1) and not mistargeted to glutamatergic synapses. Therefore, we show that three proteins of the DPC, dystrophin, β-DG, and syntrophin, are part of the postsynaptic specialization of GABAergic synapses in cultured hippocampal neurons. The fact that DPC proteins are never mistargeted to glutamatergic presynaptic terminals suggests a common clustering mechanism that involves signaling with GABAergic presynaptic terminals.

Syntrophin- and β-dystroglycan cluster opposite GABAergic terminals and colocalize with α2 GABAA receptors. a–f, Double immunofluorescence staining of pan-syntrophin (a, d, red) and GAD (b, e, green), with overlay (c, f).a–c, Low magnification; d–f, high magnification. Staining of syntrophin revealed bright clusters (a, arrows) aligned in chains coinciding with GABAergic terminals stained with GAD (b,arrows). d, e, Typical grouping of postsynaptic specializations around GABAergic boutons. g–i, Double immunofluorescence staining of β-dystroglycan (d, red) and GAD (e, green); overlay in i. Strongly stained β-dystroglycan clusters are apposed to GAD-positive boutons. j–l, Double immunofluorescence staining of α2 (j,red) and β-dystroglycan (k,green); overlay in l. The two markers colocalize in chain-like structures representing appropriately matched GABAergic synapses, but not in mismatched synapses (arrowheads). Scale bars: a–c,g–i, 10 μm; d–f, 2 μm;j–l, 5 μm.
Clustering of DPC proteins is independent of postsynaptic GABAA receptors and gephyrin
The GABAA receptor γ2 subunit is essential for postsynaptic clustering of GABAA receptors and gephyrin, as shown in vitro and in vivo in neurons from γ20/0 mice (Essrich et al., 1998), indicating that gephyrin and GABAA receptors are interdependent components of the GABAergic postsynaptic apparatus. We used hippocampal primary cultures of E15 γ20/0 mouse embryos to investigate whether clustering of the DPC is dependent on the presence of postsynaptic gephyrin and GABAAreceptors.
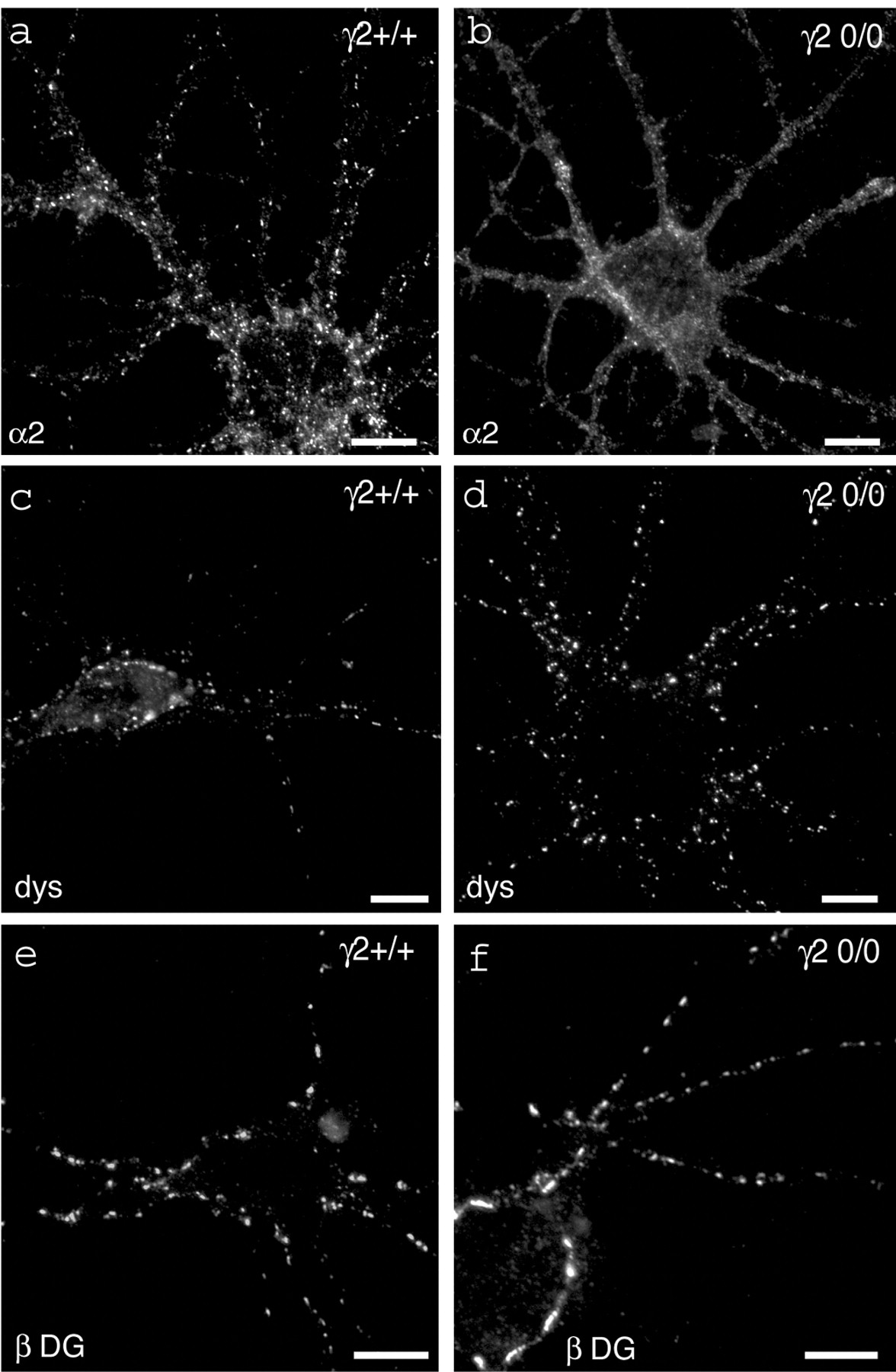
As expected, clustering of the α2 subunit was intact in γ2+/+neurons (Fig. 6a) but greatly reduced in neurons from γ20/0 mice (Fig.6b). Likewise, gephyrin-positive clusters were dramatically reduced in cultures from mutant mice (data not shown). Remarkably, however, the punctate staining of dystrophin (Fig.6c,d) and β-DG (Fig.6e,f) was unaffected by the absence of postsynaptic GABAA receptors and gephyrin. The size and density of clusters covering dendrites and somata of neurons were similar in cultures of wild-type and γ20/0 mice, as revealed by statistical analysis (Fig. 6c–f, Table 2). This result confirms that gephyrin and the DPC are independent postsynaptic components of GABAergic inhibitory synapses and clustered by distinct mechanisms.

Clustering of DPC-proteins is unaffected in γ20/0 mutant mice. a,b, Loss of α2 subunit-positive clusters in pyramid-like cells from a γ20/0culture (b) compared with a wild-type, γ2+/+ culture (a). This confirms the requirement of the γ2 subunit for postsynaptic clustering of α2-GABAA receptors. c,d, Dystrophin staining in wild-type (c) and mutant (d) neurons. The staining pattern of dystrophin was unaffected by the mutation: in both genotypes, dystrophin displayed strongly stained clusters on the soma and dendrites of cultured neurons. e,f, β-Dystroglycan staining of wild-type (e) and mutant (f) cultures. Like dystrophin, β-dystroglycan staining revealed bright staining of synaptic clusters independently of the genotype. See Table1 for semiquantitative results. Scale bars, 10 μm.
Clustering of the DPC is unaffected in γ20/0 mice
DISCUSSION
In this study, we show that mismatched apposition of GABAergic postsynaptic components to glutamatergic presynaptic terminals can occur in multi-innervated hippocampal cultures. The proportion of mismatched synapses is inversely proportional to the extent of GABAergic input. Furthermore, we show that dystrophin is clustered in association with β-dystroglycan and syntrophin at GABAergic postsynaptic sites. The molecular organization of these GABAergic synapses therefore appears homologous to the NMJ. Unlike gephyrin and GABAA receptors, these DPC proteins were never mistargeted to the membrane opposite glutamatergic terminals, suggesting distinct clustering mechanisms for gephyrin and the DPC. Supporting this idea, DPC clustering is unaffected in γ20/0 mice, although postsynaptic accumulation of GABAAreceptor and gephyrin is strongly impaired. These results suggest that clustering of the DPC at GABAergic synapses selectively depends on signaling from presynaptic terminals.
Mismatched apposition of GABAA receptors and gephyrin to glutamatergic terminals
The occurrence of mismatched synapses in isolated hippocampal neurons (Rao et al., 2000) was taken as evidence for the existence of both a “general” factor, leading to unspecific clustering of receptors and associated proteins at postsynaptic sites, and “transmitter-specific” factors responsible for appropriate matching of presynaptic and postsynaptic elements in multi-innervated cells. Here, we show that mismatched apposition of GABAAreceptor and gephyrin clusters to glutamatergic terminals occurs even in neurons receiving sparse GABAergic innervation, suggesting that the general synaptogenic factor(s) also operates when neurons receive input from more than one neurotransmitter. Nevertheless, the absence of mismatched synapses in cells innervated by multiple GABAergic axons (Fig. 2g–j) indicates that the specific signal(s) is dominant over the general clustering signal.
Because GABAA receptors and gephyrin do not seem to interact directly, it is assumed that they are linked by additional protein(s). Mistargeting of gephyrin and GABAAreceptor clusters to sites of glutamatergic input shows, however, that these putative linker proteins cannot be specific for GABAergic synapses. They either are found in all postsynaptic sites or, more likely, are preassociated with either gephyrin or GABAA receptors before coclustering at postsynaptic sites. The mismatched synapses rule out a requirement of local GABAA receptor activation for clustering, as has been postulated for glycine receptors (Kirsch and Betz, 1998;Levi et al., 1998). Therefore, clustering of glycine and GABAA receptors, although requiring gephyrin in both cases, probably is governed by distinct mechanisms.
Mismatched synapses are most likely nonfunctional, as shown by mutual exclusion of glutamatergic and GABAergic postsynaptic proteins (Rao et al., 2000; Brünig et al., 2002), raising the question of their physiological significance. In vivo, mistargeting of GABAA receptors to glutamatergic terminals is unlikely to occur in the hippocampus, because on CA1 pyramidal cells, GABAergic innervation is established before glutamatergic innervation (Tyzio et al., 1999). However, Nusser et al. (1996) reported the presence of the GABAA receptor α6 subunit postsynaptic to mossy fiber inputs in cerebellar glomeruli, suggesting that misplaced GABAA receptors are not just an in vitro artifact.
Differential mechanisms of gephyrin and dystrophin clustering
Our results demonstrate two principal differences in clustering of gephyrin and dystrophin at postsynaptic sites: (1) gephyrin, but not dystrophin, requires the GABAA receptor γ2 subunit for clustering at GABAergic synapses; (2) dystrophin, but not gephyrin, clusters selectively opposite GABAergic terminals. The two proteins therefore appear to serve independent functions. In agreement, clustering of gephyrin was unaffected by the lack of dystrophin in mdx mice (Knuesel et al., 1999), although the number and size of GABAAreceptor clusters was significantly reduced in these mutants.
In vivo, residual α2, α3, β2/3, and γ2 GABAA receptor clusters have been observed in spinal cord and organotypic retina cultures of neonatal gephyrin0/0 mice (Fischer et al., 2000; Kneussel et al., 2001). The significance of this observation is unclear, but it suggests the existence of gephyrin-independent clustering mechanisms. Dystrophin is unlikely to mediate this type of gephyrin-independent clustering, because it is detected at late postnatal stages only (Knuesel et al., 2000).
Putative role of dystrophin and the DPC in central synapses
The demonstration that dystrophin is closely associated with β-DG and syntrophin suggests that the entire DPC, including dystrobrevin and α-DG (Ervasti and Campbell, 1991; Grady et al., 2000), is enriched in a subset of central inhibitory synapses. Given the high degree of homology between GABAA and AChR subunits (Ortells and Lunt, 1995), it is not surprising to find that corresponding synapses share at least some of their protein components. At the NMJ, the DPC is essential for maturation of the postsynaptic apparatus and for long-term maintenance of AChR clusters, but not for initial receptor clustering (Grady et al., 2000). A similar role of the DPC at GABAergic synapses is consistent with the fact that dystrophin is first detectable only at P14 in rats (Knuesel et al., 1999). Thus, initiation of GABAA receptor clustering is clearly independent of dystrophin.
Interestingly, glutamatergic synapses on mature neurons are unsusceptible to actin depolymerization, but in young neurons, the synaptic apparatus depends on F-actin (Zhang and Benson, 2001). The actin independence correlates with the acquisition of scaffolding molecules, such as Bassoon, on the presynaptic site and PSD95 in excitatory postsynapses (Zhang and Benson, 2001). Allison et al. (2000)have shown that GABAA receptor and gephyrin clusters in hippocampal neurons also are unaffected by depolymerization of microtubules or actin or by detergent extraction, suggesting a stabilizing scaffold at inhibitory synapses. The DPC might play this role in GABAergic postsynapses, which would explain the altered clustering of GABAA receptors in adultmdx mice (Knuesel et al., 1999).
Given the fact that dystrophin is expressed in regions of the brain that display the highest levels of plasticity (cerebral cortex, hippocampus, cerebellum), two general possibilities for the function of the DPC are conceivable. First, because dystrophin expression is increased late in development (Knuesel et al., 1999), it might be paralleled by a decrease in synaptic plasticity in rodent brain. By stabilizing the postsynaptic apparatus, the DPC might “freeze” GABAergic synapses to maintain a certain status of the network once learning processes have been primarily completed in rats. Alternatively, the DPC might provide a scaffold, enabling changes in clustered GABAA receptor number without losing the postsynaptic apparatus, as might be required in circuits with a high degree of synaptic plasticity. Assuming that the DPC is stably associated with the presynapse, the existence of “empty” synapses, transiently devoid of GABAA receptors, would be possible.
The selective presence of dystrophin and associated proteins in GABAergic synapses in our cultures suggests that their aggregation is triggered by a signal specific for GABAergic terminals. Although agrin is well known to induce neurotransmitter receptor clustering at the NMJ, homologous synapse-specific signals have not yet been identified in the CNS. Whether soluble factors or, alternatively, transmembrane cell adhesion molecules operate at GABAergic synapses to recruit the DPC remains to be elucidated.
Footnotes
This project was supported by the Swiss National Science Foundation (Grant 31-52869.97). We thank Dr. H. Möhler for his continuous support and insight and Corinne Sidler for technical assistance. We are grateful to Dr. B. Gasnier (Centre National de la Recherche Scientifique, Institut de Biologie Physico-Chimique, Paris, France) and to Dr. S. C. Froehner (University of Washington, Seattle, WA) for a generous supply of antibodies against VIAAT and syntrophin, respectively.
Correspondence should be addressed to Dr. Jean-Marc Fritschy, Institute of Pharmacology and Toxicology, University of Zurich, Winterthurerstraße 190, CH-8057 Zurich, Switzerland. E-mail:fritschy{at}pharma.unizh.ch.
I. Knuesel's present address: California Institute of Technology, Mail Code 216-76, Pasadena, CA 91125.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}