

Abstract
Our laboratory has shown that the ability of the suprachiasmatic nuclei (SCN) to regulate a number of rhythmic processes may be compromised by the time females reach middle age. Therefore, we examined the effects of aging on the rhythmic expression of two neuropeptides synthesized in the SCN, vasoactive intestinal polypeptide (VIP) and arginine vasopressin (AVP), using in situhybridization. Because both VIP and AVP are outputs of the SCN, we hypothesized that age-related changes in rhythmicity are associated with alterations in the patterns of expression of these peptides. We found that VIP mRNA levels exhibited a 24 hr rhythm in young females, but by the time animals were middle-aged, this rhythm was gone. The attenuation of rhythmicity was associated with a decline in the level of mRNA per cell and in the number of cells in the SCN producing detectable VIP mRNA. AVP mRNA also showed a robust 24 hr rhythm in young females. However, in contrast to VIP, the AVP rhythm was not altered in the aging animals. The amount of mRNA per cell and the number of cells expressing AVP mRNA also was not affected with age. Based on these results we conclude that (1) various components of the SCN are differentially affected by aging; and (2) age-related changes in various rhythms may be attributable to changes in the ability of the SCN to transmit timing information to target sites. This may explain why the deterioration of various rhythmic processes occurs at different rates and at different times during the aging process.
Virtually all organisms exhibit 24 hr rhythms in numerous physiological and behavioral processes. In mammals, these rhythms are entrained to the light/dark (LD) cycle by a circadian pacemaker located in the suprachiasmatic nuclei (SCN) (Meijer and Rietveld, 1989). With age, many endogenous rhythms are blunted, and their entrainment to the LD cycle changes (Wise, 1984; Cohen and Wise, 1988; Weiland and Wise, 1989; Monk, 1991; Copinschi and Van Cauter, 1994; Li and Satinoff, 1995; Monk et al., 1995; Van Cauter et al., 1996; Wise et al., 1996). Evidence suggests that these disruptions may be attributable to alterations in the circadian pacemaker. First, some (Pittendrigh and Daan, 1974) but not all (Vaswanathan and Davis, 1995;Duffy and Davis, 1997) studies demonstrate that the period of activity rhythms (Ralph et al., 1990) shortens with age. Second, the ability of aged animals to shift their activity patterns in response to photic and nonphotic stimuli changes (Rosenberg et al., 1991; Penev et al., 1995;Zhang et al., 1996). Third, endogenous rhythmicity in spontaneous firing of SCN neurons in vitro is blunted (Satinoff et al., 1993). Some of these alterations can be reversed by implanting fetal SCN tissue into the third ventricle (Van Reeth et al., 1994;Vaswanathan and Davis, 1995; Cai et al., 1997a,b) suggesting that these changes in the expression of circadian rhythms result in part from changes in SCN function.
Neurons in the SCN synthesize numerous neuropeptides (van den Pol and Tsujimoto, 1985) that may constitute output pathways to other brain regions. Few studies have examined the effects of aging on the rhythmic expression of these peptides. We assessed the effects of aging on the rhythmic gene expression of vasoactive intestinal polypeptide (VIP) and arginine vasopressin (AVP). VIP is synthesized primarily in the ventral SCN, whereas AVP is produced in a different population of neurons in the dorsomedial SCN (van den Pol and Tsujimoto, 1985; Watts and Swanson, 1987). Although these neuronal populations synapse with each other (Kaikoku et al., 1992), little is known about how they interact. Both VIP and AVP neurons relay circadian information to the basal forebrain and various hypothalamic and thalamic nuclei (Watts and Swanson, 1987). These projections regulate rhythms of gonadotropin-releasing hormone (GnRH) release (Cheesman et al., 1977;Osland et al., 1977; van der Beek et al., 1995; Harney et al., 1996;Palm et al., 1997) and glucocorticoid release (Scarbrough et al., 1996;Buijs, 1997) and activity (Pickard and Turek, 1983; Sollars and Pickard, 1995; Murphy et al., 1997). Thus, age-related alterations in VIP or AVP expression may affect the transmission of rhythmic information from the SCN to target sites. We also chose to examine the effects of aging on these two peptides because aging differentially affects their expression in other brain regions. In the cortex, there is a dramatic decline in VIP cell number with aging (Andreose et al., 1994; Cha et al., 1995, 1997; Huh et al., 1997). In contrast, AVP synthesized in the paraventricular nuclei (PVN) and supraoptic nuclei of the hypothalamus is unaltered or mildly depressed by aging (Dobie et al., 1991; Sladek and Olschowka, 1994), whereas AVP synthesized in the bed nucleus of the stria terminalis is reduced (Lucassen et al., 1998). Thus, the effects of aging on these two neuropeptides may be region-dependent.
We assessed the effects of age on the rhythmicity of these neuropeptides in females because cyclic reproduction, which is intimately linked to the circadian clock (Turek and Van Cauter, 1988;Hastings, 1991), exhibits profound changes by the time animals are middle-aged (Wise et al., 1996). In young females, the timing of the proestrous (Legan and Karsch, 1975) and estrogen-induced (Legan et al., 1975) surge of luteinizing hormone (LH) is tightly coupled to the LD cycle. However, in middle-aged animals, the LH surge is delayed, and the amplitude of the surge is attenuated (Wise, 1982; Nass et al., 1984). We have demonstrated that the rhythmic expression of multiple neurotransmitters and receptors (Weiland and Wise, 1986, 1989; Cohen and Wise, 1988; Lloyd et al., 1991) is blunted by middle age, suggesting that fundamental changes in the SCN may cause a deterioration in the ability to maintain regular reproductive cycles. In the present experiment, we assessed VIP and AVP mRNA levels in the SCN of young, middle-aged, and old females. If aging involves a change in all the essential elements (i.e., inputs, oscillators, and outputs) of the SCN, the pattern of expression of both these peptides would be affected. On the other hand, if only some of the components of the clock are affected with age, we might observe differential effects on VIP compared with AVP.
MATERIALS AND METHODS
Animals. Young, regularly cycling (2–4 months; six to eight animals per time point), middle-aged, irregularly cycling (10–12 months; six animals per time point), and old, persistent diestrous (18–20 months; six animals per time point) females were housed in a 14/10 hr LD cycle (lights on at 0400 hr) with food and water available ad libitum. Vaginal cytology was checked daily for at least 2 weeks to determine the reproductive state of animals. All animals were ovariectomized under Metofane (methoxyflurane) inhalant anesthesia. One week later, SILASTIC capsules containing 17-β-estradiol (180 μg/ml in sesame oil; young, 20 mm capsule; middle-aged and old, 30 mm capsule) were implanted subcutaneously to clamp plasma estradiol at equivalent and physiological levels in all experimental groups (Wise, 1984). This is critical because ovarian steroids modulate the period of circadian activity (Morin et al., 1977; Takahashi and Menaker 1980; Albers et al., 1981) and the precise pattern of expression of VIP mRNA (Krajnak et al., 1997). Animals were killed at the following times after estrogen treatment: 2400 hr (day 1) and 0300, 0800, 1200, 1600, 2000, and 2300 hr (day 2).
VIP and AVP in situ hybridization. In situ hybridization methods were the same as previously described (Wise et al., 1992; Krajnak et al., 1997). Briefly, brains were removed, rapidly frozen, and stored at −70°C until sectioned. Frozen coronal sections (12 μm) through the basal forebrain and hypothalamus were sliced, thaw-mounted onto slides, and stored at −80°C until processed for in situ hybridization (ISH). Slides containing sections from the middle to midcaudal SCN (three slides or six sections per animal) were chosen for VIP ISH, and alternate slides (two slides or four sections per animal) were chosen for AVP ISH. The riboprobe for VIP was generated using a 500 bp human cDNA directed against exons 3–6 of the VIP–peptide histidine isoleucine gene (from Dr. R. H. Goodman, Vollum Institute). The riboprobe for AVP was generated using a 241 bp cDNA directed against exon C of the rat AVP gene (provided by Dr. T. Sherman, Georgetown University). Both riboprobes were transcribed using 50 μm total UTP. Because each cDNA contains a different percentage of UTP, we used different amounts of35S-UTP to produce comparable incorporation of the radiolabeled nucleotide (VIP, 12.5 μm35S-UTP and 37.5 μm unlabeled UTP and SP6 polymerase; AVP, 37.5 μm35S-UTP and 12.5 μmunlabeled UTP and SP6 polymerase). Slides were thawed, fixed with 4% paraformaldehyde, and dehydrated using a series of increasing concentrations of ethanol. Hybridization buffer (50 μl) containing 400 ng/ml labeled VIP cRNA or 200 ng/ml AVP cRNA was applied to each slide. In preliminary studies, saturation curves were generated and revealed that these concentrations of cRNA produced maximal labeling without significantly increasing background. Slides were incubated in humid chambers at 55°C for 18 hr, washed under stringent conditions, dehydrated with ethanol, coated with Kodak NTB2 emulsion (Eastman Kodak, Rochester, NY; diluted 1:1 with distilled water), and stored at 4°C. Slides processed for AVP were developed 5 d after emulsion coating, and slides processed for VIP were developed 10 d after emulsion coating. All slides were counterstained with 0.05% toluidine blue so that individual cell bodies could be identified.
All slides were examined for the presence of labeling in the SCN. If the SCN from an individual animal was damaged, mRNA levels were not quantified in those slides. Therefore, in some animals, AVP was not quantified, and in others, VIP was not quantified. Gene expression was quantified using the Bioquant OS/2 image analysis system. Slides from a number of animals were examined, and a single threshold for determining grains versus background was set. The perimeter of an individual cell was outlined, and both the area of the cell and portion covered by grains (i.e., above threshold) were quantified. Lighting and contrast levels were standardized before taking measurements to assure that all slides were assessed under the same conditions. Background was assessed by taking measurements over unlabeled cells outside the area of interest. Cells having a value five times higher than background were considered labeled.
Analyses. To determine whether aging altered VIP or AVP gene expression per cell in the SCN or the number of cells labeled for these peptides, 3 (age) × 7 (time of day) ANOVAs were performed. Planned comparisons using one-way ANOVAs examining the effects of time on gene expression in each age group were performed to determine whether gene expression was rhythmic. Post hoc comparisons were made using Newman–Kuels tests. Differences were considered significant ifp < 0.05.
RESULTS
VIP gene expression
Two-way ANOVA revealed a main effect of age on VIP mRNA per cell (Fig. 1) (F(2,101)= 5.99; p < 0.004) but no effect of time (F(6,101) = 0.81; p = 0.57) and no interaction (F(12,101) = 1.50;p = 0.14). Further analyses showed that VIP mRNA levels per cell were lower in middle-aged and old than in young females (p < 0.05). VIP-expressing cells in all animals were seen predominantly in the ventrolateral SCN. However, as animals aged, there appeared to be a loss of VIP gene expression in the most medially located neurons (Fig. 2). The two-way ANOVA on the number of VIP-expressing cells in the SCN revealed a main effect of age (F(2,101) = 4.53;p < 0.02) but no effect of time (F(6,101) = 1.27; p = 0.28) and no interaction (F(12,101) = 0.80;p = 0.65). Further analyses showed that the number of cells expressing VIP mRNA was lower in the middle-aged and old females than in the young females (Fig. 3).

Overall levels of VIP mRNA per cell in young, middle-aged, and old females (mean ± SEM). The 3 (age) × 7 (time of day) ANOVA revealed a significant effect of age (F(2,101) = 5.99; p < 0.004) on mRNA levels per cell. VIP gene expression was significantly lower in middle-aged and old females compared with young (p < 0.05).

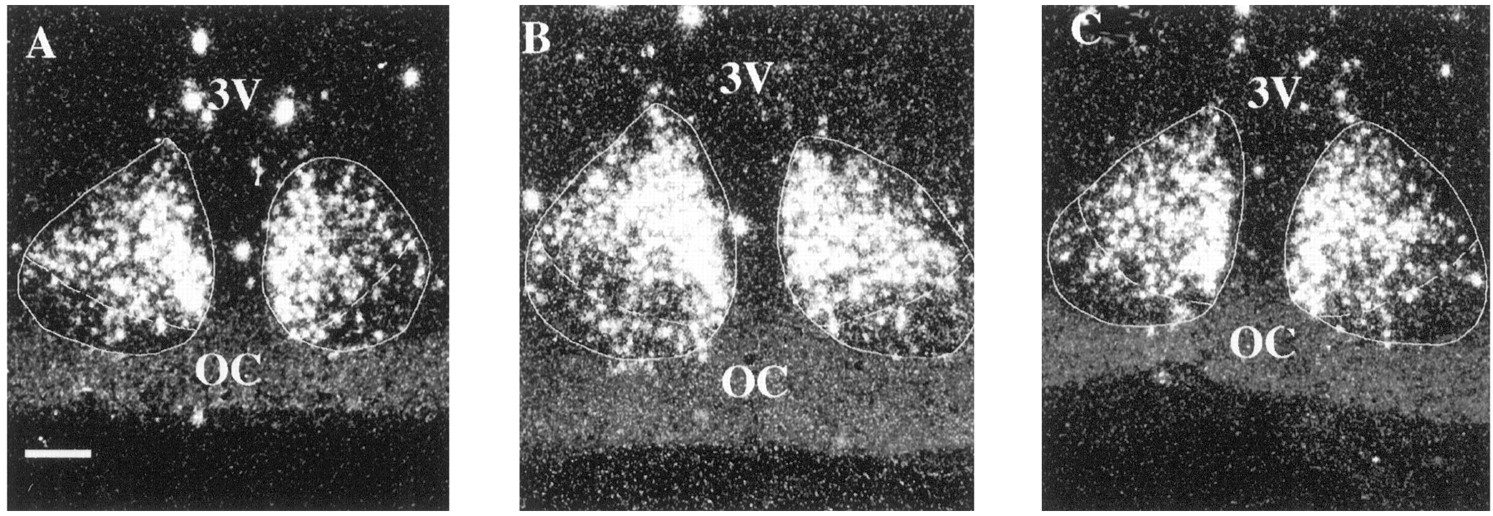
This photomicrograph shows VIP labeling in the SCN of young (A), middle-aged (B), and old (C) females at 1200 hr. VIP labeling was seen predominantly in the ventrolateral portion of the SCN in all animals. However, as animals age, there appears to be a decrease in the number of VIP-expressing cells in the medial portion of the nucleus. 3V, Third ventricle;OC, optic chiasm. Scale bar, 25 μm.

VIP-expressing cells per section (mean ± SEM) in young, middle-aged, and old females. The two-way ANOVA analyzing the effects of age and time of day on the number of VIP-expressing cells revealed a main effect of age (F(2,101) = 4.53; p < 0.02), with the number of VIP-expressing cells per section being higher in young than in middle-aged or old females (p < 0.05).
To determine whether there was a rhythm in VIP gene expression in young, middle-aged, and old females, we performed planned comparisons using one-way ANOVAs on VIP mRNA levels at each age. In young females, VIP gene expression was rhythmic (F(6,43) = 3.03; p < 0.02), with mRNA levels being lower at 0300, 0800, and 2000 hr than at the other time points (p < 0.05; Fig.4). By the time animals reached middle age, a rhythm in VIP gene expression was no longer detectable (middle age, F(6,31) = 0.61; p = 0.72; old, F(6,27) = 0.93, p = 0.49).

VIP mRNA levels per cell (mean ± SEM) over time in young (•), middle-aged (○), and old (▾) females. One-way ANOVA revealed that VIP mRNA was rhythmic in young females (F(6,43) = 3.03; p < 0.02), with VIP gene expression being higher at 2400, 1200, and 2300 hr than at other times of day (*p < 0.05). VIP mRNA levels did not significantly fluctuate over the day in middle-aged or old females. The black bar under thex-axis represents the dark phase of the cycle, and thewhite bar represents the light phase.
AVP gene expression
Two-way ANOVA revealed a main effect of time on the amount of AVP mRNA per cell (Fig. 5) (F(6,108) = 18.73; p < 0.001) but no effect of age (F(2,108) = 0.64;p = 0.53) and no interaction (F(12,108) = 0.55; p = 0.87).Post hoc analyses examining the effect of time revealed a prominent rhythm in AVP gene expression in all groups. AVP mRNA levels were low between 2400 and 0300 hr and exhibited significant increases at each time point from 0800 to 1600 hr (p < 0.05). After 1600 hr, AVP mRNA returned to baseline levels, showing a significant decrease at both 2000 and 2300 hr (p< 0.05). Planned comparisons using one-way ANOVAs to determine whether AVP mRNA expression was rhythmic were consistent with the two-way ANOVA; AVP gene expression was rhythmic in all age groups (young,F(6,54) = 10.17; p < 0.001; middle-aged, (F(6,28) = 6.99; p< 0.001; and old, F(6,26) = 3.81;p < 0.008), with AVP mRNA levels being highest at 1600 hr and lowest at 0300 hr (p < 0.05 in all age groups).

AVP-expressing cells per section (mean ± SEM) in young, middle-aged, and old females. Aging did not alter the number of AVP-expressing cells in the SCN of females (F(2,108) = 0.928; p = 0.399).
AVP gene expression was seen predominantly in the dorsomedial portion of the SCN in all animals examined (Fig.6). The two-way ANOVA on the number of cells expressing AVP mRNA per section revealed a main effect of time on cell number (F(6,108) = 11.19; p< 0.001) but no effect of age (F(2,108) = 0.93;p = 0.40) and no interaction (F(12,108) = 1.43; p = 0.16). The effect of time on cell number was similar to that seen on AVP mRNA levels (Fig. 7). The number of cells expressing AVP mRNA was low between 2400 and 0800 hr and increased to reach peak levels by 1600 hr (p < 0.05). The number of AVP-expressing cells then began to fall and decreased significantly by 2300 hr (p < 0.05). Aging did not alter this pattern, nor did it affect the number of cells expressing AVP mRNA (Fig. 8).

These photomicrographs show AVP mRNA labeling in the SCN of young (A), middle-aged (B), and old (C) females at 1200 hr. AVP expression was seen primarily within the dorsomedial portion of the SCN in all groups of animals. 3V, Third ventricle; OC, optic chiasm. Scale bar, 25 μm.

AVP-expressing cells per section (mean ± SEM) over time in young (•), middle-aged (○), and old females (▾). The two-way ANOVA examining the effects of age and time of day on AVP cell number revealed a significant effect of time (F(6,108) = 11.19; p < 0.001), with the number of AVP-expressing cells gradually increasing between 0300 and 1600 hr and then declining to baseline levels by 2300 hr (different letters are significantly different from each other; p < 0.05). The black bar under the x-axis represents the dark phase of the cycle, and the white bar represents the light phase.

AVP mRNA levels per cell (mean ± SEM) over time in young (•), middle-aged (○), and old females (▾). The 3 (age) × 7 (time of day) ANOVA revealed a main effect of time on AVP mRNA levels (F(6,108) = 18.73;p < 0.001), with AVP mRNA levels increasing between 0300 and 1600 hr and then declining to baseline levels by 2300 hr (different letters are significantly different from each other; p < 0.05). The black bar under the x-axis represents the dark phase of the cycle, and the white bar represents the light phase.
DISCUSSION
Previous studies demonstrate that aging alters the expression of numerous physiological and behavioral rhythms that are driven by the SCN. This raises the possibility that age-related deterioration of the pacemaker itself may underlie the changes in rhythmicity of these diverse physiological endpoints. Therefore, the purpose of this study was to examine the effects of aging on two major peptides synthesized within the SCN, VIP and AVP, to determine whether the rhythmic expression of their mRNAs was altered.
We focused our attention on these two neuropeptides because (1) they are critical neuropeptides synthesized rhythmically in the SCN (Inouye et al., 1993); this rhythmic expression dictates the ability of the SCN to interpret environmental cues (Kiss et al., 1984; Bosler and Beaudet, 1985; Hisano et al., 1988; Ibata et al., 1989) and drive multiple outputs (Osland et al., 1977; Sodersten et al., 1983; Sollars and Pickard, 1995; van der Beek et al., 1995; Harney et al., 1996;Scarbrough et al., 1996; Palm et al., 1997); and (2) these peptides also modulate the timing of GnRH secretion (van der Beek et al., 1995;Huhman and van der Beek, 1996; Palm et al., 1997), which is critical to cyclic reproduction in females. Our study was performed in females because reproductive success in this sex is intimately tied to the SCN and begins to display overt signs of senescence by middle age (Wise et al., 1996, 1997). Our results clearly demonstrate a selective age-related alteration in VIP gene expression in the absence of any change in AVP gene expression.
The rhythm of VIP mRNA was not detectable by the time animals reached middle age. In young females, VIP mRNA showed a peak at ∼1200 hr and a smaller rise during the midevening. This finding is consistent with our previous work, which showed that mRNA levels in females are high during the day, and that this daytime rise is coincident with the timing of the LH surge in young females (Krajnak et al., 1997). Other studies also indicate that if VIP activity in young females is blocked in the hour just before the surge with either antisense oligonucleotides to VIP (Harney et al., 1996) or VIP antibodies (van der Beek et al., 1995), the timing of the surge is delayed, and the amplitude is attenuated. Thus, the loss of the daytime peak of VIP mRNA in middle-aged animals may be responsible for the delay in the timing and attenuation in the amplitude of the LH surge.
The rise in VIP mRNA during the middle of the dark phase (2300–2400 hr) was also attenuated in aging animals. In young males, VIP mRNA levels and peptide concentrations in the SCN are high at night and decline after lights on (Albers et al., 1990; Inouye et al., 1993;Krajnak et al., 1997). Thus, it has been hypothesized that these light-induced changes in VIP may provide environmental LD information to the pacemaking cells in the SCN (Albers et al., 1991). However, it is unclear whether the rise in VIP mRNA at 2300–2400 hr that we observe in young females serves the same function. If it does, this signal is absent by the time females reach middle age. Because VIP neurons receive input from all major afferents to the SCN (Kiss et al., 1984; Bosler and Beaudet, 1985; Hisano et al., 1988; Ibata et al., 1989), this decline in mRNA could be in part responsible for the decreased ability of both photic and nonphotic stimuli to phase shift rhythms in aged animals (Rosenberg et al., 1991; Penev et al., 1995;Zhang et al., 1996). Similar changes may also occur in males. Kawakami et al. (1997) found a slight decrease in VIP mRNA at night in old males (22–24 months) when they monitored gene expression at one time during the light phase and one time during the dark phase. The number of VIP-immunolabeled cells within the SCN during the light phase of the cycle may also decline in very old (33–34 months) males (Chee et al., 1988).
Several possible mechanisms may underlie the age-related decline in the rhythm of VIP mRNA in the SCN. First, both photic input from the retina (Ibata et al., 1989) and serotonin (5-HT) input from the raphe (Kiss et al., 1984; Bosler and Beaudet, 1985) regulate the rhythm of VIP gene expression and peptide concentrations. Eliminating photic input to the SCN of adult males by placing animals in persistent darkness (Inouye et al., 1993) or by enucleation (Okamoto et al., 1990) results in a loss of VIP rhythmicity and an overall increase in VIP expression. Thus, it is possible that alterations in the retinal input to the SCN lead to the loss of rhythmic VIP gene expression by middle age. In aged animals, light-induced fos expression is attenuated in the SCN (Sutin et al., 1993; Zhang et al., 1996; Cai et al., 1997), which is concomitant with a decline in the magnitude of light-induced phase shifts (Zhang et al., 1996). However, subjecting aged animals to brighter light pulses partially reverses these dampened responses (Zhang et al., 1996), suggesting that there may be age-related affects on the ability of the retina to perceive and transmit photic information. However, retinal degenerative mice, whose retina show signs of aging relatively early, do not exhibit any decline in the ability to respond to phase-shifting light pulses (Garcia-Fernandez et al., 1995). Together, these observations suggest that the retinal signal may be maintained with aging, but that the ability of the SCN to receive this signal may be altered. Because retinal input appears to synapse directly onto VIP-producing neurons (Ibata et al., 1989), and light induces fos expression within VIP neurons (Daikoku et al., 1992;Speh and Moore, 1996; Rominj et al., 1996), it is possible that age-related changes in VIP alter the transmission of photic information to other pacemaking target cells within the SCN or efferent targets in other regions of the brain. However, the phenotype of neurons showing reduced fos activation in aged animals needs to be determined before we know exactly which outputs of the SCN might be altered.
VIP concentrations (Kawakami et al., 1985) and gene expression (Okamura et al., 1995) are also modulated by 5-HT. 5-HT terminals from the raphe synapse directly on VIP-producing neurons in the SCN (Kiss et al., 1984), and lesioning this input results in a loss of VIP and a dramatic decline in VIP expression within the SCN rhythmicity (Kawakami et al., 1985, 1994; Okamura et al., 1995). Studies have shown that the rhythm in 5-HT turnover is altered by the time females reach middle age (Cohen and Wise, 1988) and that 5-HT projections to the SCN are diminished (Van Luijtelaar et al., 1989). The ability of 5-HT to cause phase shifts in activity rhythms is also reduced in aged animals (Penev et al., 1995). Thus, the age-related changes in 5-HT input to the SCN may have dramatic effects on both the rhythmic expression of VIP and the amount of VIP expressed.
The most surprising finding of this study is that the AVP rhythm remains completely intact in the same animals in which the rhythmic expression of VIP was absent. The rhythm in gene expression, the number of cells expressing AVP, and the amount of AVP mRNA per cell were unaltered, even in the oldest group of females. These results are consistent with data collected from aged males showing that the number of AVP-immunopositive neurons in the SCN was not altered in males 16–18 months of age (Roozendaal et al., 1987; Lucassen et al., 1995). These findings are remarkable in that most entrained rhythms that have been examined show age-related alterations in rhythmic expression (Wise et al., 1987, 1997; Satinoff et al., 1993; Sutin et al., 1993; Zhang et al., 1996; Cai et al., 1997).
AVP projections from the SCN serve as an output, carrying timing information to a number of target areas, including regions involved in generation of the LH surge (Watson and Langub, 1996) and generation of the rhythm in corticosteroid release (Buijs, 1997). Our findings showing that neither the rhythmic expression of AVP nor the amount of AVP expressed per cell changes during aging helps explain the maintenance of the daily rhythm in corticosterone despite the absence of a rhythm in corticotropin-releasing hormone (CRH) mRNA in the PVN of aged animals (Cai and Wise, 1996). Buijs (1997) has reported a multisynaptic pathway between SCN AVP neurons and the adrenal that bypasses PVN CRH. In addition, our data lead us to conclude that changes in the timing or amplitude of the LH surge seen in middle-aged animals are unlikely attributable to changes in the AVP signal. However, it is possible that changes in the ability of target sites to recognize the AVP signal occur in aging females.
The rhythm in AVP synthesis and release is endogenous and driven by the SCN (Gillette and Reppert, 1987; Inouye et al., 1993). Thus, these rhythms are maintained in constant conditions. However, under LD conditions, the AVP rhythm is also tightly coupled to the LD cycle (Burbach et al., 1988; Inouye et al., 1993). Our findings show that this rhythm remains tightly coupled to the LD cycle throughout aging. However, other entrained rhythms regulated by the SCN and synchronized to the LD cycle, such as glucose utilization (Wise et al., 1987), Fos induction by the LD cycle (Sutin et al., 1993; Zhang et al., 1996; Cai et al., 1997), and activity rhythms (Satinoff et al., 1993), lose their tight coupling to the LD cycle during aging. In fact, when temperature, activity, and drinking rhythms are monitored in the same elderly animal, a deterioration in the entrainment and amplitude of one rhythm is not necessarily correlated with a deterioration in the other rhythms (Satinoff et al., 1993). Together these studies support the hypothesis that age-related changes in the expression of rhythmic processes are the result of the uncoupling of various oscillators within the SCN and not necessarily attributable to complete deterioration of the ability of the SCN to generate rhythms.
In summary, our results show that the rhythmic expression of VIP in the SCN is undetectable by the time females reach middle age, but AVP expression in these same animals is unaltered. Based on these findings, we suggest that the differential effects of age on these two outputs may explain why certain rhythms are altered with age, whereas others remain intact. Future studies should concentrate on changes in the inputs to the SCN to determine how age-related alterations in these inputs may affect the ability of the SCN to entrain rhythms to the environment. Other studies, concentrating on how individual oscillators are coupled and how this coupling is altered by aging, will also help us determine why some rhythms are more resistant to the effect of aging than others.
Footnotes
This work was supported by National Institutes of Health Grants AGO2224 to P.M.W., AGO5755 to K.K., and AGO5762 to M.L.K. We thank Susan Steman and Dr. Jacob P. Harney for technical assistance. We also thank Dr. R. H. Goodman (Vollum Institute) and Dr. T. Sherman (Georgetown University) for supplying us with cDNAs to VIP and AVP.
Correspondence should be addressed to Dr. Kristine Krajnak, Department of Physiology, MS508 Chandler Medical Center, University of Kentucky, Lexington, KY 40536-0084.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}