


Abstract
Previous studies showed that an extended 6-h session duration produced an increasing rate of cocaine self-administration in rats. The present study further investigated the effect of dose and session duration on cocaine self-administration. Eight groups of rats (4 doses × 2 session durations) self-administered one of four cocaine doses (0.25, 0.5, 1, and 2 mg/kg/injection) in either 1- or 6-h sessions under a fixed-ratio schedule. In another experiment, two other groups of rats self-administered 0.5 mg/kg/injection of cocaine in either 3- or 12-h sessions. Cocaine self-administration increased at all doses in 6-h sessions but not in 1-h sessions. Cocaine intake (milligram/kilogram) reached an asymptote earlier at a higher dose, but the rate of responding increased faster when the dose was lower. In ShA rats, the cocaine dose-response function was higher in rats at the two higher unit doses than at the lower doses. Cocaine self-administration increased in 6- and 12-h sessions, but not in 1- and 3-h sessions. The increase in self-administration was faster and greater in 12-h sessions than 6-h sessions. The data suggest that cocaine self-administration increases at various doses with prolonged access and that an increase in the rate of responding is positively and inversely associated with session duration and unit dose, respectively. Results also imply that cocaine intake reaches a ceiling faster at high doses even under short session duration. Therefore, high doses or prolonged access to cocaine are more likely to result in a pattern of cocaine intake that reflects compulsive use.
Drug self-administration in laboratory animals has been extensively used in research of drug abuse/dependence. The predictive validity of self-administration in laboratory animals has been supported by the observation that a majority of drugs abused by humans are reliably self-administered by laboratory animals (Johanson and Balster, 1978; Roberts et al., 1989). However, systematic studies of different doses and session durations have been limited in animal studies. In humans, it is known that cocaine-dependent users take the drug in a cyclic manner, such as frequent intake for hours or days followed by days of drug abstinence that were followed by the next high rate of drug intake (Gawin and Ellinwood, 1989; Gawin, 1991). Moreover, drug-dependent users seem to take drugs in an increasing amount during the development of drug dependence (American Psychiatry Association, 2000). In an effort to understand factors that lead to a compulsive pattern of drug intake in humans, research on the effect of parameters on the pattern of drug self-administration in laboratory animals has been performed.
Focusing on psychostimulants, researchers found that a stable level of self-administration can be maintained over a long period of time with relatively short session durations in laboratory animals. However, Deneau et al. (1969) reported that unlimited access to cocaine and amphetamine induced an erratic pattern of self-administration in rhesus monkeys, such as a high rate of self-administration followed by an abrupt cessation of self-administration with lethality within 30 days. Similar observations were reported in rats and in rhesus monkeys with unlimited access to cocaine, amphetamine, or methamphetamine (Johanson et al., 1976; Bozarth and Wise, 1985). Later studies demonstrated that discrete but frequent access to cocaine for 10 or 21 days could produce a “binge”-like pattern of cocaine self-administration in rats disrupting a circadian pattern of self-administration (Fitch and Roberts, 1993; Roberts et al., 2002). With limited access, it was found that daily 6-h sessions engendered an escalating rate of cocaine and methamphetamine self-administration under a fixed-ratio (FR) schedule in rats (Ahmed and Koob, 1998; Liu et al., 2005; Kitamura et al., 2006). Therefore, a prolonged period of self-administration, either within a session or over daily sessions, was suggested to be an important determinant in producing a pattern of self-administration in laboratory animals that reflected increases in drug intake.
It was hypothesized that laboratory animals can regulate the self-administration of psychostimulants within a session, regardless of unit dose, maintaining a constant level of drug intake (Lynch et al., 1998; Panlilio et al., 2006). However, it was also suggested that by increasing a dose of cocaine, a circadian pattern of self-administration, could be disrupted, and a “binge”-like pattern of self-administration could be manifested in rats under a fixed-ratio schedule (Finch and Robert, 1993; Roberts et al., 2002). Moreover, in 7-h sessions, 2.0 mg/kg/injection of cocaine, but not 0.5 mg/kg/injection, produced increased cocaine intake in milligrams/kilogram under a fixed-ratio schedule in rats (Mantsch et al., 2004). In contrast, a study by Liu et al. (2005) demonstrated that the rate of self-administration of 0.75 mg/kg/injection of cocaine increased faster and to a greater extent than that of 1.5 mg/kg/injection of cocaine in daily 6-h sessions in rats.
In our laboratory, prolonged session duration of 6 h has been shown repeatedly to produce an increasing intake of cocaine and methamphetamine under a fixed-ratio schedule in rats (e.g., Ahmed and Koob, 1998; Kitamura et al., 2006). To test the hypothesis that escalation in cocaine self-administration changes with dose, the present study investigated the effect of four cocaine doses (0.25, 0.5, 1, and 2 mg/kg/injection) on the rate of cocaine self-administration and intake (milligram/kilogram) in 6-h sessions. Secondly, it may be easily predicted from the literature described above that extending session duration will lead to a faster escalation in cocaine self-administration. To test this hypothesis, the systematic comparison across various session durations at the same experimental dose was explored. An increase in the rate of self-administration and intake (milligram/kilogram) of 0.5 mg/kg/injection of cocaine was compared across four session durations (1, 3, 6, and 12 h) in rats. The overall hypothesis was that escalation in the intake and the rate of self-administration of cocaine would be positively related to dose and session duration.
Materials and Methods
All animal use procedures were approved by The Scripps Research Institute's Animal Care and Use Committee and were in accordance with the National Institutes of Health guidelines.
Animals and Apparatus
Sixty male Wistar rats (Charles River, Hollister, CA), each weighing between 275 and 300 g at the beginning of the study, served as subjects. They were housed in groups of three in plastic cages with a 12:12-h light/dark cycle with lights on at 6:00 AM. Food and water were available ad libitum except during the food training. During experimental sessions, each rat was placed in a standard operant chamber, which was placed in a light- and sound-attenuating cubicle (28 × 26 × 20 cm; Med Associates Inc., St. Albans, VT). The front door and the back wall of the chamber were made of transparent plastic, and the other walls were opaque metal. The chamber had two retractable response levers mounted on one side of the opaque walls and a food hopper located between the levers. A stimulus light was mounted above each lever. A drug injection was delivered by a syringe pump (Razel Scientific Instruments, Georgia, VT) located on top of the cubicle. Experimental sessions were controlled and recorded by a PC computer with custom interface and software in the experimental room.
Procedure
For surgery, rats were anesthetized with 2 to 3% isoflurane. They were implanted with a silastic catheter (0.3 mm i.d. × 0.64 mm o.d.; Dow Corning Co. Midland, MI) into the right external jugular vein under aseptic conditions. The distal end of the catheter was subcutaneously threaded to the back of the rat where it exited the rat via a metal guide cannula (22G; Plastics One Inc., Roanoke, VA) that was anchored at the back of the rat. After surgery, rats were given analgesics (Flunixin, 2.5 mg/kg s.c.). An antibiotic (Timentin, 20 mg i.v.; SmithKline Beecham, Philadelphia, PA) was administered to the rats for a minimum of 1 week. The catheter was flushed daily with heparinized saline (30 U/ml). The patency of catheters in the rats was tested using an ultra short-acting barbiturate methohexital sodium (Brevital, 10 mg/ml, 2 mg/rat) whenever catheter failure was suspected during the study. Generally, a total loss of muscle tone within 3 s after a methohexital sodium injection indicates the patency of a catheter.
Experimental sessions were conducted once a day, 7 days a week during the dark (active) cycle. Immediately before a session, rats were transferred from the vivarium to an experimental room where operant chambers were located. After being flushed with 0.9% saline, a rat's indwelling catheter was connected to a tube that exited the chamber through a metal spring and a swivel and was connected to a syringe pump. After the drug delivery system was connected to the rat, the chamber was closed and the session was started immediately. The start of a session was signaled by the presentation of two response levers into the chamber. Responding on the right lever resulted in the delivery of 0.1 ml of a drug injection over 4 s. During an injection, stimulus lights above both levers were illuminated and lasted throughout a time-out period (20 s) that followed each injection. Pressing the left lever was counted but had no other programmed consequences. No priming injection was used in this paradigm. After the session, the catheter was filled with 0.9% saline containing heparin (30 U/ml), and the rat was returned to the home cage.
Experiment 1: Effect of Dose on Cocaine Self-Administration
Animals. Forty-eight rats (n = 6 for each group × 4 cocaine doses × 2 access conditions) initially served as subjects for the experiment. Three rats that self-administered 2 mg/kg/injection died in the middle of a session during an escalation period. Consequently, the data from those rats were excluded from the data analysis. During the determination of a dose-response function in each group, two more rats at 2 mg/kg/injection died in the middle of a baseline escalation-session, and two rats within the 0.25 mg/kg/injection group lost their catheters during this period as well.
Procedure. Rats were initially food-restricted (approximately 15–20 g/rat/day) and trained to press a lever for food under a FR1 schedule of reinforcement. That is, one lever press resulted in the delivery of one food pellet (45 mg). After food-maintained responding became stable (approximately 20 responses/10 min), the training session ended and food was made available ad libitum for the remainder of the study. The rats were then implanted with intravenous catheters as described above and allowed to recover for at least 3 days. The rats were randomly divided into four groups (n = 12 per group). Each group of rats was allowed to self-administer one of four doses (0.25, 0.5, 1, or 2 mg/kg/injection) of cocaine in a 1-h session under an FR1 schedule. The baseline sessions lasted for 11 days.
After the baseline sessions, the rats in each group were subdivided into two groups balanced by the number of injections/session on the last baseline session. During an escalation period, one group of rats (LgA) was allowed to self-administer a baseline dose of cocaine for 6 h per day, whereas the other group (ShA) was allowed to do so for 1 h per day. After 21 sessions of an escalation period, a dose-response function of cocaine self-administration was determined in each group under an FR1 schedule. Various doses of cocaine were made available in each group of rats. Each dose of cocaine was tested in a 1-h session for three consecutive sessions, and the data from three sessions were averaged for analysis. Three sessions identical to those in the escalation period (LgA, 6-h sessions; ShA, 1-h sessions) separated the test sessions of two different doses of cocaine. Doses of cocaine were examined in the same sequence across groups and in an ABA design where the first dose tested was retested at the end of the experiment for comparison.
Experiment 2: Effect of Session Duration on Cocaine Self-Administration
Animals. Twelve rats (3-h session, n = 6; 12-h session, n = 6) were initially catheterized and trained to self-administer cocaine. However, one rat in the group of 3-h session duration was soon excluded from the experiment because of a catheter failure. Therefore, only five rats in the 3-h group finished the escalation sessions. Within the 12-h group, on the 19th escalation session, one rat died during a 12-h session, but the data of the rat of up to 19 sessions were included in the data analysis on 21 days of escalation sessions. During the determination of the dose-response function of cocaine in each group of rats, one rat with a 3-h session duration lost its catheter and two rats with a 12-h session duration died, one in the middle of a session and the other immediately after a session in the home cage.
Procedure. The general procedure was as described in experiment 1. Twelve rats were initially trained to press a lever for food under an FR1 schedule. After food-maintained responding became stable (approximately 20 responses/10 min), the rats were catheterized and allowed to self-administer 0.5 mg/kg/injection of cocaine in 1-h sessions for 7 days. The reason we chose 0.5 mg/kg/injection of cocaine to evaluate the effect of various durations of access on self-administration was that doses of cocaine in previous studies (Ahmed and Koob, 1998, 1999) were approximately 0.5 to 0.6 mg/kg/injection and because the self-administration of 0.5 mg/kg/injection of cocaine was examined under 1- and 6-h sessions in experiment 1. After baseline sessions, the rats were divided into two groups balanced by the number of injections/session on the last baseline session. During an escalation period, one group of rats was allowed to self-administer cocaine for 3 h per day, whereas the other group was allowed to do so for 12 h per day. An escalation period lasted for 21 days. After 21 sessions of the escalation period, the dose-response function of cocaine self-administration was determined in each group of rats, where various doses of cocaine were made available under an FR1 schedule. A dose of cocaine was tested in a 1-h session for three consecutive sessions. Three sessions identical to those in the escalation period (3 or 12 h sessions) separated the test sessions of two different doses of cocaine. Doses of cocaine were examined in an ABA design.
Data Analysis
General Analysis. The data were expressed as the mean number of injections and milligram/kilogram within a session as well as during the 1st hour of sessions for each group of rats and subjected to statistical analysis. Cocaine self-administration across daily sessions was compared within each group using a one-way analysis of variance (ANOVA) with repeated measures and the Student-Newman-Keuls post hoc test (Prism 4.0; GraphPad, San Diego, CA). Furthermore, an interaction between dose and cocaine self-administration and between session duration and cocaine self-administration was assessed using a two-way ANOVA with repeated measures (dose × session or session duration × session; Prism 4.0, GraphPad). The magnitude of a change in cocaine self-administration during the escalation period was measured as the difference in the number of injections as well as in intake (milligram/kilogram) between the 1st and the last escalation session in each group. Comparison of the magnitude of an increase in self-administration across groups was performed using a one-way ANOVA with the Student-Newman-Keuls post hoc test after the data were transformed to log values because of unequal variance across groups (Prism 4.0; GraphPad). A correlation between the magnitudes of an increase in cocaine self-administration and doses or session durations was evaluated using the Pearson's Product-Moment test on the transformed data (Prism 4.0; GraphPad). The pattern of responding for cocaine was expressed as the mean number of injections per hour over session durations in LgA rats. Differences in the rate of responding at every hour between the first and the last three escalation sessions were evaluated using a two-way ANOVA with the Bonferroni post hoc test [session (the first versus the last sessions) × hour; Prism 4.0; GraphPad].
Specific Analysis: Effect of Dose on Cocaine Self-Administration. The dose-response functions of cocaine that were obtained at the end of the escalation period were compared between LgA and ShA groups of the same unit dose using a two-way ANOVA with the Bonferroni post hoc test [session duration (1 versus 6 h) × cocaine dose; Prism 4.0; GraphPad]. Cocaine intake (milligram/kilogram/session and milligram/kilogram/1st hour) in the first escalation session was compared across doses within a short or long access condition. Because of unequal variance across the groups, the data were first transformed to log values and then analyzed using a one-way ANOVA with the Student-Newman-Keuls post hoc test (Prism 4.0; GraphPad). Cocaine intake on the 21st escalation session was also compared across doses within a short or long access condition using the same statistical analysis. Furthermore, cocaine intake in the 1st hour of a session in LgA rats was compared with that of ShA rats of the same unit dose using the Student's t test (Prism 4.0; GraphPad).
Specific Analysis: Effect of Session Duration on Cocaine Self-Administration. Data on 1- and 6-h session durations were taken from those of 0.5 mg/kg/injection of experiment 1, and they were analyzed with the data of 3- and 12-h session durations. The dose-response functions of cocaine that were obtained at the end of the escalation period were compared across four groups using a two-way ANOVA with the Bonferroni post hoc test [session duration (1, 3, 6, and 12 h) × cocaine dose; Prism 4.0;, GraphPad, San Diego, CA]. The pattern of responding for cocaine during the 1st hour of sessions was expressed as the mean number of injections per bin (10 min). Differences in the rate of responding at every bin between the first and the last three escalation sessions were evaluated using a two-way ANOVA with the Bonferroni post hoc test [session (the first versus the last sessions) × bin; Prism 4.0; GraphPad]. To evaluate the regularity of responding in rats under the two extreme session durations, the distribution of inter-reinforcement intervals (IRI) was compared between the 12- and 1-h rats. Mean IRI and coefficient of variation on IRI values were calculated in each rat and were averaged across rats within a group. Statistical difference in mean IRI and coefficient of variation between the 1- and 12-h rats were examined using Student's t test after the data were transformed to log values (Prism 4.0; GraphPad).
Cocaine intake (milligram/kilogram/session and as milligram/kilogram/1st hour) on the first escalation session was compared across groups. Because of unequal variance across the groups, the data were first transformed to log values and then analyzed using a one-way ANOVA with the Student-Newman-Keuls post hoc test (Prism 4.0; GraphPad). Cocaine intake on the 21st escalation session was also compared across groups using the same statistical analysis. A correlation between cocaine intake and session duration was evaluated using the Pearson's Product-Moment test after the data were transformed to log values (Prism 4.0; GraphPad).
Drugs. (-)-Cocaine hydrochloride was provided by the National Institute on Drug Abuse (Rockville, MD). The drug was dissolved in sterile 0.9% saline for self-administration. Doses of cocaine for self-administration were prepared by varying the concentration of cocaine in drug solutions, whereas an injection volume was kept constant at 0.1 ml/injection. Furthermore, each drug solution was prepared for each rat based on its body weight, and it was updated every 2 or 3 days. Doses of cocaine were expressed as salt.
Results
Effect of Dose on Cocaine Self-Administration. When cocaine intake (milligram/kilogram) was compared across unit doses, the intake in session 1 tended to be higher in rats at the two higher unit doses than those at the two lower doses under both ShA and LgA conditions (Table 1). At the end of the escalation period (session 21), cocaine intake per session as well as in the 1st hour of the session was significantly higher only in rats that self-administered 2 mg/kg/injection than in the rats from the other groups under both ShA and LgA conditions. Cocaine intake during the 1st hour of a session in LgA rats at the unit doses of 0.25 and 0.5 mg/kg/injection of cocaine was significantly greater than ShA rats of the same unit doses in session 21. No difference was found in cocaine intake during the 1st hour of a session between ShA and LgA rats that self-administered 1 and 2 mg/kg/injection in both sessions 1 and 21.
Comparison of cocaine intake in rats that self-administered various doses of cocaine Responding was maintained under an FR1 schedule of reinforcement.
With 6 h of session duration, the rate of cocaine self-administration in LgA rats significantly increased at all unit doses starting in sessions 5, 11, 13, and 15, respectively, at 0.25, 0.5, 1, and 2 mg/kg/injection compared with that on the first escalation session (Fig. 1). A two-way ANOVA showed that there was a significant interaction between dose and escalation in the rate of self-administration [F(60,380) = 2.90, p < 0.001]. Cocaine self-administration during the 1st hour of sessions in LgA rats also significantly increased starting in sessions 4 and 11, respectively, at 0.25 and 0.5 mg/kg/injection (Fig. 2). There was a significant interaction between dose and escalation in the rate of self-administration in the 1st hour of sessions [F(60,380) = 3.39, p < 0.001]. Although cocaine self-administration in ShA rats tended to increase at 0.25 mg/kg/injection of cocaine over the first 13 sessions, responding decreased again, and statistical significance was not achieved at the end of the escalation period at any dose (Fig. 2).
The magnitude of the increase in the rate of self-administration was greater in LgA rats that self-administered 0.25 mg/kg/injection of cocaine than in those at the other doses (0.25 mg/kg/injection, 149.3 ± 19.9 injections; 0.5 mg/kg/injection, 50.5 ± 13.0 injections; 1 mg/kg/injection, 17.5 ± 4.7 injections; 2 mg/kg/injection, 13.8 ± 5.3 injections). The magnitude of the increase in the rate of self-administration was inversely correlated with unit dose in LgA rats (Pearson r2 = 0.9410, p < 0.05). Likewise, the magnitude of the increase in the rate of self-administration during the 1st hour of sessions was greater in LgA rats at 0.25 mg/kg/injection than in those at the other doses of cocaine (0.25 mg/kg/injection, 34.2 ± 2.9 injections; 0.5 mg/kg/injection, 11.2 ± 4.2 injections; 1 mg/kg/injection, 3.0 ± 1.1 injections; 2 mg/kg/injection, 3.8 ± 0.9 injections). Furthermore, the increase in the rate of self-administration during the 1st hour of sessions was greater in LgA rats at 0.5 mg/kg/injection than at 1 and 2 mg/kg/injection of cocaine. No correlation was found between the magnitude of the increase in the rate of self-administration during the 1st hour and unit doses (Pearson r2 = 0.1404, p = 0.63). When calculated in milligram/kilogram, the increase of cocaine intake in milligram/kilogram in LgA rats did not significantly differ across four unit doses within a session as well as during the 1st hour of sessions [session, F(3,19) = 1.601; 1st hour, F(3,19) = 1.2, p > 0.2].

Self-administration of cocaine during the session under a fixed-ratio schedule of reinforcement. Each panel represents self-administration of one of four doses of cocaine (0.25, 0.5, 1, or 2 mg/kg/injection) during sessions. Data are expressed as the number of injections/session on the left axis and milligram/kilogram/session on the right axis. Error bars are mean ± S.E.M. values. Open symbols are the data in rats with 1-h session duration (ShA). Filled symbols are the data in rats with 6-h session duration (LgA). Circles, self-administration of 0.25 mg/kg/injection of cocaine; squares, self-administration of 0.5 mg/kg/injection of cocaine; triangles, self-administration of 1 mg/kg/injection of cocaine; inverted triangles, self-administration of 2 mg/kg/injection of cocaine. *, p < 0.05; **, p < 0.01; ***, p < 0.001 compared with session 1.
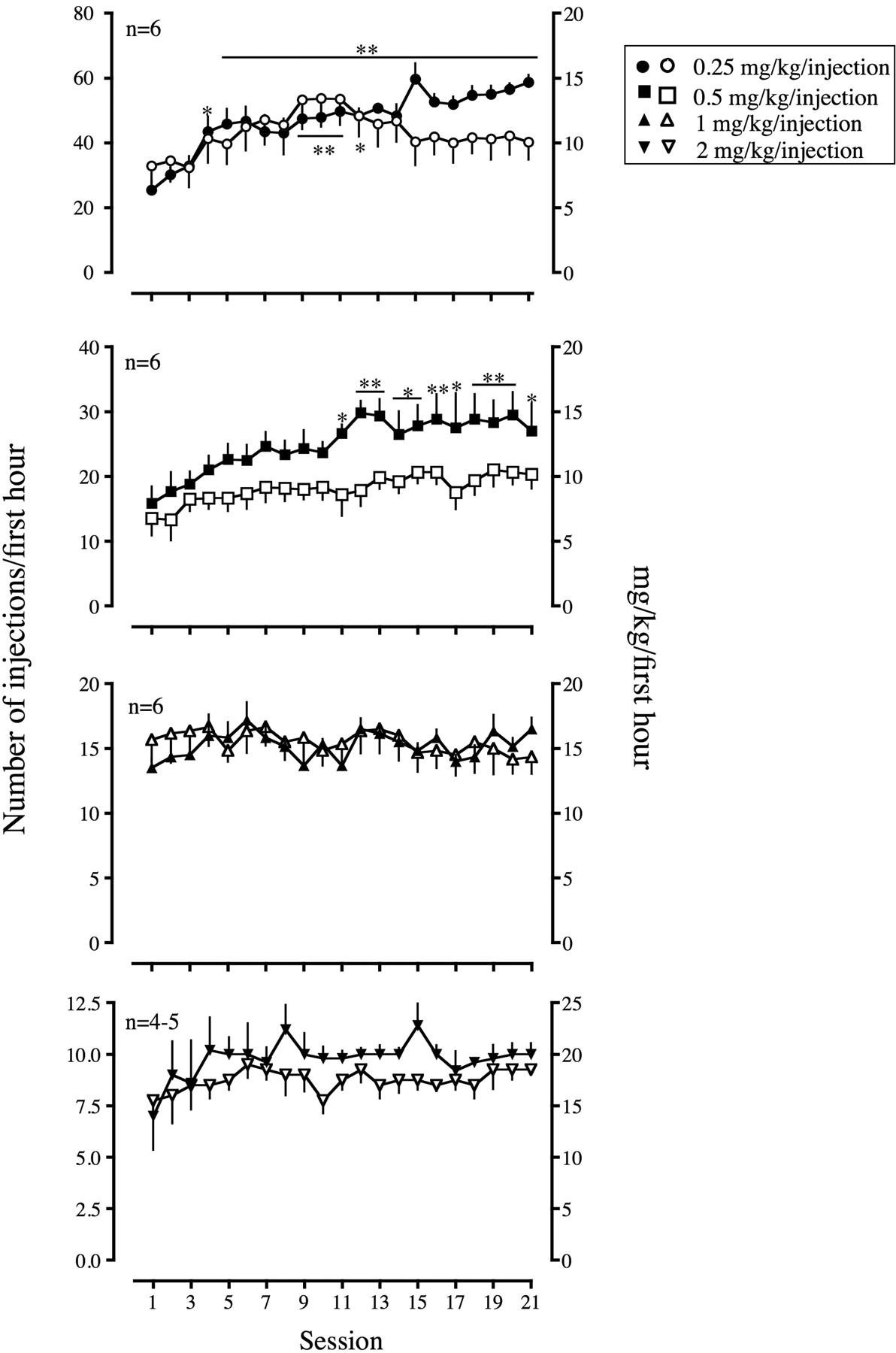
Self-administration of cocaine during the 1st hour of the session under a fixed-ratio schedule of reinforcement. Each panel represents self-administration of one of four doses of cocaine (0.25, 0.5, 1, or 2 mg/kg/injection) during the 1st hour of the session. The rest is as described in Fig. 1 legend.
When the first and the last three escalation sessions were compared, the pattern of responding showed an increased rate of self-administration in all hours of sessions in LgA rats that self-administered 0.25 and 0.5 mg/kg/injection of cocaine [0.25 group, F(1,12) = 3529, p < 0.001; 0.5 group, F(1,12) = 655.9, p < 0.001; Fig. 3]. LgA rats that self-administered 1 and 2 mg/kg/injection of cocaine did not show a significant increase in the rate of responding between the first and the last three escalation sessions.
At the end of 21 days of the escalation period, a dose-response function of cocaine was determined in each group of rats (Fig. 4). Cocaine maintained a higher rate of responding, and consequently intake at all of the doses of cocaine tested, in LgA than in ShA rats that self-administered 0.25 or 0.5 mg/kg/injection of cocaine during the escalation period [0.25 group, F(1,6) = 269.4, p < 0.001, Fig. 4A; 0.5 group, F(1,6) = 169.5, p < 0.001, Fig. 4C]. In contrast, no difference in the dose-response function was noted between LgA and ShA rats that self-administered 1 or 2 mg/kg/injection during the escalation period (Fig. 4, B and D). In addition, the dose-response functions of cocaine overlapped across all groups of LgA rats (Fig. 4F). The dose-response functions of cocaine in ShA rats that self-administered 0.25 and 0.5 mg/kg/injection of cocaine were significantly lower than those in ShA rats that self-administered 1 and 2 mg/kg/injection of cocaine (Fig. 4E).
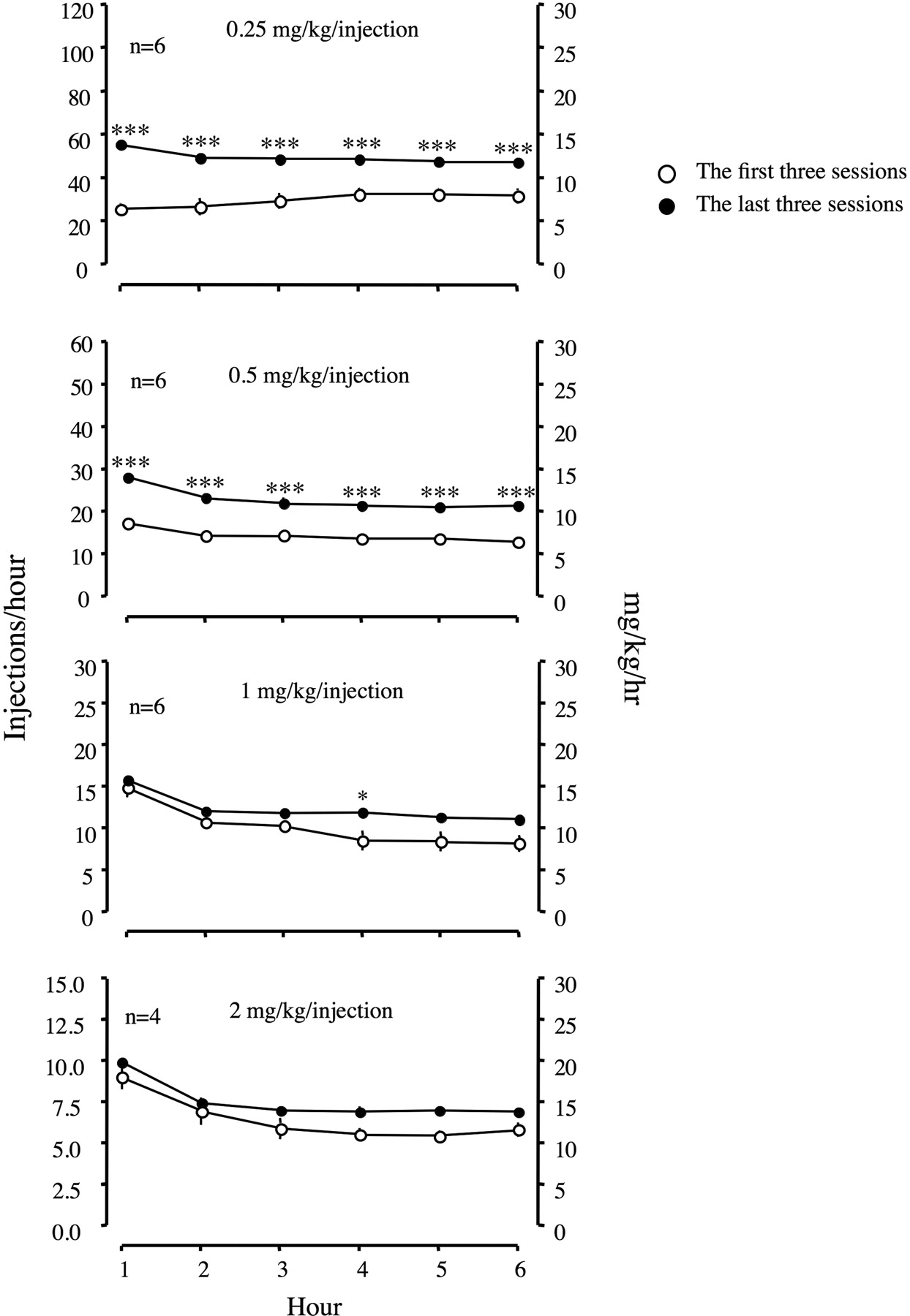
Comparison of a pattern of responding between the first and last three sessions during the escalation period in LgA rats. Responding was maintained under a fixed-ratio schedule. Data are expressed as the number of injections per hour. Error bars are mean ± S.E.M. values. Top, the data of LgA rats that were allowed to self-administer 0.25 mg/kg/injection. Middle, the data of LgA rats that were allowed to self-administer 0.5 mg/kg/injection. Bottom, the data of LgA rats that were allowed to self-administer 1 mg/kg/injection. Bottom, the data of LgA rats that were allowed to self-administer 2 mg/kg/injection. Open circles represent the mean of the first three escalation sessions, and filled circles represent the mean of the last three escalation sessions during 21 days of the escalation period. *, p < 0.05; **, p < 0.01 compared with the first three sessions at each time point.
Effect of Session Duration on Cocaine Self-Administration. Cocaine intake (milligram/kilogram) during the 1st hour of the session was significantly higher in rats under 3- and 12-h session durations in the first escalation session (Table 2, p < 0.01). However, in session 21, the intake in the 1st hour of a session was higher in rats under 12-h sessions than in rats under the other session durations (p < 0.001). In addition, cocaine intake during the 1st hour of a session was higher in rats with a 3-h session duration than in those with a 1-h session duration in session 21 (p < 0.05).
Comparison of cocaine intake in rats under various session durations The dose of cocaine was 0.5 mg/kg/injection in each group. Responding was maintained under an FR1 schedule of reinforcement.
Data on 1- and 6-h session durations were taken from those of 0.5 mg/kg/injection of experiment 1. During 21 days of the escalation sessions, rats under a 12-h session did not gain any weight, starting at an average of 382 g in session 1 and ending at an average of 364 g in session 21. Rats in the other groups generally gained weight during the escalation period (1-h group, 357–421 g; 3-h group, 343–399 g; 6-h group, 366–409 g). Two of the rats under a 12-h session duration died during an escalation session, which was inserted between dose-response test sessions as a baseline session.
With a 12-h session duration, the increase in self-administration of cocaine during a session achieved statistical significance starting in session 6, whereas it did so in session 11 with a 6-h session duration (Fig. 5, top). Likewise, cocaine self-administration during the 1st hour of sessions significantly increased starting in sessions 9 and 11, respectively, with 12- and 6-h session durations (Fig. 5, bottom). Cocaine self-administration did not increase in the 1- and 3-h sessions. There was an interaction between session duration and escalation in the rate of self-administration during a session as well as in the 1st hour of sessions [F(60,380) = 3.45 and 1.56, p < 0.001 and p < 0.01, respectively].

Dose-response function of cocaine under a fixed-ratio schedule. Various doses of cocaine were made available in each group of rats during a test session. Test sessions lasted 1 h. Data are the number of injections/session. Error bars indicate mean ± S.E.M. values. When error bars do not appear, they are within the symbols. Each graph represents responding by each group of rats that self-administered one of four doses of cocaine during an escalation period (inverted triangles, 0.25 mg/kg/injection; circles, 0.5 mg/kg/injection; squares, 1 mg/kg/injection; triangles, 2 mg/kg/injection). Open symbols are the data from ShA rats, and filled symbols are the data from LgA rats. Graph E represents responding in ShA rats. Graph F represents responding in LgA rats. **, p < 0.01; ***, p < 0.001 compared with the ShA group. ##, p < 0.01; ### p < 0.001 compared with the ShA groups of 0.25 and 0.5 mg/kg/injection of cocaine.
The magnitude of the increase during the escalation period was significantly greater in rats under a 12-h session duration than in those under a 6-h session duration (6-h group, 50.5 ± 13.0 injections/6 h; 12-h group, 165.3 ± 25.6 injections/12 h). The increase in self-administration during the 1st hour of sessions did not differ between 6- and 12-h session durations (6-h group, 11.2 ± 4.2 injections/h; 12-h group, 18.8 ± 1.3 injections/h). There was no significant correlation between session durations and the magnitude of the change in cocaine self-administration (session, Pearson r2 = 0.0383, p = 0.80; 1st hour, Pearson r2 = 0.0846, p = 0.71).

Self-administration of cocaine under various session durations. Each group of rats self-administered 0.5 mg/kg/injection of cocaine under a fixed-ratio schedule. The session duration was 1 (inverted triangles), 3 (squares), 6 (triangles), or 12 h (circles) depending on groups. Data are expressed as the number of injections on the left axis and milligram/kilogram on the right axis. Top, cocaine self-administration in sessions; bottom, cocaine self-administration during the 1st hour of the sessions. Error bars are mean ± S.E.M. values. The data under 1- and 6-h sessions were taken from Fig. 1. *, p < 0.05; **, p < 0.01; ***, p < 0.001 compared with session 1.
When the first and the last three escalation sessions were compared, the pattern of responding exhibited an increased rate of responding in all hours of sessions in rats under 6- and 12-h session durations [6-h group, F(1,5) = 268, p < 0.001; 12-h group, F(1,11) = 190.0, p < 0.001, Fig. 6, inset]. Furthermore, responding for cocaine significantly increased at all bins within the 1st hour of sessions with 6- and 12-h session durations (Fig. 6). When the distribution of injections during a session was compared in session 21 between 1- and 12-h rats, the mean IRI in 12-h rats was significantly smaller than the mean IRI in 1-h rats (Table 3). However, coefficient of variation on IRI values did not differ between the two groups.
Comparison of the inter-reinforcement intervals between the rats with one-hour session duration and the rats with 12-h session duration The dose of cocaine was 0.5 mg/kg/injection in each group. Responding was maintained under an FR1 schedule of reinforcement. Values are mean ± S.E.M. Mean IRI and CV values were calculated in each rat and were averaged across rats within a group (1-h, n = 6; 12-h, n = 5).
At the end of 21 days of the escalation period, the dose-response function of cocaine was determined in each group of rats (Fig. 7). The rats in 12-h sessions produced the highest responding for all of the doses of cocaine tested among all the groups [F(3,18) = 117.0, p < 0.001]. Furthermore, cocaine maintained higher responding for all of the doses of cocaine tested in rats under 3- and 6-h sessions than in rats under 1-h sessions. No difference in the dose-response function was noted between rats under 3- and 6-h sessions. The rate of self-administration under 12-h access to cocaine was relatively regular in all hours of a session (Fig. 8).
Discussion
The present study demonstrated that cocaine self-administration increased to an asymptote at various doses with a 6-h session duration, which supports and extends previous findings (Ahmed and Koob, 1998, 1999). The final intake (milligram/kilogram) and the asymptote of intake in LgA rats self-administering 0.25, 0.5, or 1 mg/kg/injection was not significantly different between groups, except that the LgA rats at 2 mg/kg/injection achieved a higher intake than those at the other doses. The LgA rats at 1 or 2 mg/kg/injection reached the asymptote of intake in earlier sessions than those at 0.25 or 0.5 mg/kg/injection. In addition, intake in the ShA rats at the two highest doses of cocaine also reached the same level of the 1st hour of a session as LgA rats.

Comparison of pattern of responding during the 1st hour of sessions between the first and last three sessions of the escalation period across session durations. Responding was maintained under a fixed-ratio schedule. Data are expressed as the number of injections per bin (10 min). Error bars are mean ± S.E.M. values. When error bars do not appear, the bars are within symbols. Open symbols represent the mean of the first three escalation sessions, and filled symbols represent the mean of the last three escalation sessions during 21 days of the escalation period. Inset, pattern of responding at all hours of sessions in 3- and 12-h groups. *, p < 0.05; **, p < 0.01; ***, p < 0.001 compared with the first three sessions at each time point.

Dose-response function of cocaine in rats that self-administered cocaine under various session durations during an escalation period. Various doses of cocaine were made available in each group of rats during a test session under a fixed-ratio schedule. Test sessions lasted 1 h. Data are the number of injections/session. Inverted triangles, squares, triangles, and circles represent responding by rats that self-administered 0.5 mg/kg/injection of cocaine under a 1-, 3-, 6-, and 12-h session, respectively. During the determination of a dose-response function of cocaine, two rats in a group of a 12-h session died. **, p < 0.01, ***, p < 0.001 compared with the 1-h group. ###, p < 0.001 compared with the 3- and 6-h group.
The rate of self-administration increased faster and to a greater extent when the unit dose was lower, demonstrating an inverse relationship between dose and escalation in the rate of self-administration of cocaine. This relationship is largely due to the observation that rats at a high dose had already reached an asymptote of intake even with 1-h session duration. The present data are consistent with the results by Liu et al. (2005) that 0.75 mg/kg/injection of cocaine produced a faster and greater increase in the rate of self-administration than 1.5 mg/kg/injection. In contrast, it was reported that cocaine intake increased in 7-h sessions at the unit dose of 2 mg/kg/injection but not at 0.5 mg/kg/injection in rats (Mantsch et al., 2004). The reasons for this difference are not clear. However, different self-administration paradigms might explain the different results. In the Mantsch et al. (2004) study, rats were exposed daily to three doses of cocaine (0.5, 1, and 2 mg/kg/injection) for 3 h during the training and escalation period. In contrast, in the present study and the study by Liu et al. (2005), rats were allowed access only to their baseline doses of cocaine during those periods.
When self-administration of 0.5 mg/kg/injection of cocaine was further compared across different session durations, cocaine self-administration increased in rats in 6- and 12-h sessions but not in 1- and 3-h sessions, suggesting a threshold of 3 to 6 h for session duration. No change in cocaine self-administration in 3-h sessions was previously shown in rats under an FR1 schedule (Pulvirenti et al., 1992, 1994). Furthermore, cocaine self-administration increased faster in 12-h sessions than in 6-h sessions, implying a positive relationship between escalation in cocaine self-administration and session duration beyond a threshold. The magnitude of the increase in self-administration in 12-h sessions was approximately 3-fold higher than that of 6-h sessions. Likewise, cocaine intake per session (milligram/kilogram/session) in 12-h sessions was not proportionately related to that of the other session durations at the end of the escalation period. This suggests that an increase in cocaine self-administration may be exponentially related to session duration and that this could lead to toxicity. This conclusion is corroborated by previous findings that unlimited access to cocaine resulted in binge-like cocaine intake with lethality in rats and rhesus monkeys (Johanson et al., 1976; Bozarth and Wise, 1985).
In the present study in which only a descending limb of the dose-response function of cocaine was observed, responding by LgA rats self-administering 0.25 or 0.5 mg/kg/injection was higher than responding by ShA rats at all of the doses of cocaine tested, consistent with previous reports (Ahmed and Koob, 1998, 1999). Moreover, responding by rats self-administering 0.5 mg/kg/injection in 12-h sessions was even higher than responding by the 6-h rats. Therefore, the data suggest that the dose-response function of cocaine was shifted upward as session duration increased. An upward shift of a psychostimulant dose-response function was also found in rats and rhesus monkeys after chronic self-administration/administration of the drugs (Woolverton et al., 1984; Schenk and Partridge, 1997; Mantsch et al., 2004).
In pharmacology, increased drug intake with repeated administration probably represents tolerance to a drug effect. Tolerance is often characterized by a rightward shift of the dose-response function of a drug (O'Brien et al., 1996), but an upward shift of the dose-response function may also suggest the development of tolerance. Tolerance to the rewarding effect of cocaine was proposed to underlie the increasing cocaine intake in LgA rats (“reward allostasis”) (Ahmed and Koob, 2005). To date, no changes in D2 dopamine receptors or transporters that play a central role in the reinforcing mechanism of psychostimulants were found in LgA rats (Ben-Sharhar et al., 2006; Ahmed et al., 2003; Wee et al., 2007). Neuroadaptations between brain systems rather than within a system may account for this upward shift of the dose-response function of cocaine in LgA rats (Koob and LeMoal, 1997; Ahmed and Koob, 2005).

Responses in individual rats over 12-h session duration. The data are from the rats of the 12-h group (n = 5) in the last escalation session (session 21). Numbers on the left indicate individual rat numbers. Ticks along each horizontal line are individual responses made by the rat numbered on the left.
The upward shift of the dose-response function of cocaine was also argued to reflect tolerance to nonspecific effects of cocaine in LgA rats (Zernig et al., 2004). However, in the present study, the increased rate of responding was prominent from the start of a session in the LgA rats. Furthermore, the upward shift of the dose-response function was observed only in rats at low doses of 0.25 or 0.5 mg/kg/injection. Both observations seem to argue against tolerance to the nonspecific effects of cocaine in LgA rats.
One of the main findings in this study was an inverse relationship between escalation in the rate of self-administration and dose. Although this was unexpected, a closer inspection of the data reveals that rats at high unit doses easily achieved a high initial intake of cocaine under an FR1 schedule, allowing less room for an increase in self-administration. This argument is supported by the observation that, despite different initial cocaine intake (milligram/kilogram), the final intake in LgA rats was similar in session 21 across groups of 0.25, 0.5, and 1 mg/kg/injection. Intake by LgA rats at 2 mg/kg/injection was significantly higher than that achieved at the other doses, but with lethality in two rats. These observations suggest that there is a ceiling of cocaine intake that animals can self-administer in 6 h, but a high unit dose of cocaine under a low FR schedule allows animals to push the ceiling of cocaine intake to the limits of toxicity. The overlapping dose-response functions of cocaine in all LgA groups further support this conclusion. The other major finding was a positive relationship between escalation in cocaine self-administration and session duration. The postulated mechanism underlying this relationship is that a longer daily exposure to cocaine self-administration facilitated the rate of neuroadaptations resulting in a faster and greater increase in cocaine self-administration in rats. Future research on the neuropharmacology of LgA rats with various session durations is warranted.
Interestingly, cocaine intake by ShA rats self-administering 1 or 2 mg/kg/injection did not differ from that of the 1st hour of a session by LgA rats, and the dose-response functions of cocaine in the ShA and LgA rats at the unit doses overlapped. As with LgA rats, cocaine intake by ShA rats at 2 mg/kg/injection was significantly higher than the intake at 0.25 or 0.5 mg/kg/injection with lethality in two rats. These data imply that a high dose of cocaine allowed animals to reach a high level of intake, even with short session duration.
Total cocaine intake achieved by 2 mg/kg/injection and 12-h sessions was clearly at the limit of toxicity under the present conditions. Nonetheless, this observation does not generalize across all rodents. For example, in a study by Mantsch et al. (2004), 2 mg/kg/injection of cocaine did not result in lethality in 7-h sessions despite the fact that rats in that study achieved a higher cocaine intake (approximately 150 mg/kg/7 h) than rats at 2 mg/kg/injection in the present study (93.6 mg/kg/6 h). Likewise, Liu et al. (2005) did not report any toxic effect of cocaine intake of 1.5 mg/kg/injection in 6-h sessions (101 mg/kg/6 h). Again, reasons for differences are not clear. However, a genetic difference between Wistar rats (the present study) and Sprague-Dawley rats (Mantsch et al., 2004; Liu et al., 2005) may explain the different findings.
Finally, it is worth commenting that the rate of self-administration under 12-h access to cocaine was relatively regular in all hours of a session, despite the fact that half of the rats in 12-h sessions died during the study. Although, as expected, the mean IRI was smaller in the 12-h rats than in the 1-h rats, the coefficient of variation did not differ, suggesting no significant loss of control with a 12-h session duration. A report that the interinjection interval showed a significant increase in variability starting at 22 h of continuous access to 0.25 mg/injection of cocaine (approximately 0.6 mg/kg/injection) corroborates this conclusion (Tornatzky and Miczek, 2000).
In conclusion, the present study showed that escalation in cocaine self-administration occurs at various doses and interacts with session duration. Cocaine intake per session reached a ceiling at all doses, and the time to reach the ceiling was directly related to dose. Escalation in the rate of self-administration is positively associated with session duration beyond a threshold. Escalation in the rate of cocaine self-administration is inversely related to unit dose due to initial high level of intake at high doses. Results suggest that either high dose or prolonged session duration at a given dose can facilitate the intake of cocaine that reflects compulsive use in humans.
Acknowledgments
We thank Dr. William L. Woolverton for comments on the early version of the manuscript. We gratefully acknowledge the excellent technical assistance of Yanabel Grant and Robert Lintz. We also thank Mike Arends for editorial assistance.
Footnotes
-
This study was supported by the National Institute on Drug Abuse Grant DA004398 (G.F.K). This is publication number 18044-MIND from The Scripps Research Institute.
-
Article, publication date, and citation information can be found at http://jpet.aspetjournals.org.
-
doi:10.1124/jpet.106.113340.
-
ABBREVIATIONS: FR, fixed-ratio; LgA, long access; ShA, short access; ANOVA, one-way analysis of variance; IRI, inter-response interval.
- Received August 31, 2006.
- Accepted December 27, 2006.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}