


Abstract
The influence of dosing time on the anti-immobility effect of antidepressants and mechanisms underlying this phenomenon were investigated in mice. In the forced swimming test (FST), the immobility time of mice treated with amitriptyline (15 mg/kg) and fluvoxamine (30 mg/kg) showed a significant 24-h rhythm. The anti-immobility effect of fluvoxamine in FST was potent at the early part of the dark phase without increasing locomotor activity. Concerning pharmacokinetics, although Ke of fluvoxamine was approximately 1.3-fold higher in mice injected with fluvoxamine at 9:00 PM than at 9:00 AM, no dosing time dependence was demonstrated for either plasma or brain fluvoxamine concentration at 0.5 h after the drug injection. On the other hand, serotonin transporter (SERT) mRNA expression and 5-hydroxytryptamine (5-HT) uptake activity in the mouse midbrain showed significant time-dependent changes with higher levels during the dark phase and lower levels during the light phase. These results suggest that the reuptake of 5-HT might be more increased during the dark phase. Since the reuptake of 5-HT is inhibited almost completely by injection with 30 mg/kg fluvoxamine at any time, the extracellular 5-HT level may be more increased by the injection of fluvoxamine at the early part of the dark phase. The present results suggest that the anti-immobility effect of fluvoxamine in FST increases depending on dosing time. Furthermore, the time-dependent change of SERT mRNA expression and uptake activity in the midbrain is suggested to be the mechanism underlying the 24-h rhythm of anti-immobility effect of fluvoxamine.
A large number of physiological rhythmic variables are apparent in the central nervous system, in hormone secretion, and so on (Thomson et al., 1980; Kafka et al., 1981). In addition, many drugs vary in potency and/or toxicity according to the time in the circadian cycle when they are administered (Walker and Owasoyo, 1974; Frederickson et al., 1977; Ohdo et al., 1995, 1996, 2001). However, several drugs cause alterations in the 24-h rhythms of biochemical, physiological, and behavioral processes (Akiyama et al., 1999; Horikawa et al., 2000), depending on the dosing time. It is thus very important to consider the administration schedule of drugs in pharmacotherapy.
The major theory of depressive disorder, the monoamine hypothesis, proposes that decreasing the levels of one or more of the brain monoamine neurotransmitters, such as 5-hydroxytryptamine (serotonin) (5-HT), noradrenaline, or dopamine, can produce such diseases. A refinement of this hypothesis is that depressive illness may arise, specifically, from decreased brain 5-HT function (Coppen, 1967), and a number of factors have been proposed to account for a reduced functionality of the 5-HT system in depression. The 5-HT system has formed the basis of investigation into currently available antidepressants and still represents an area of drug development. The forced swimming test (FST) is a behavioral model developed to predict the efficacy of antidepressants in humans. Many classes of antidepressants decrease the immobility time in FST, including tricyclic antidepressants, monoamine oxidase inhibitors, and atypical antidepressants. However, 5-HT uptake inhibitors have sometimes failed to produce positive effects in FST (Detke et al., 1997), although they have therapeutic efficacy in depressive patients.
The serotonin transporter (SERT), a member of the Na+/Cl--dependent transporter family, plays a key role in central serotonergic neurotransmission (Rudnick, 1977). SERT is the target of antidepressant drugs, in particular SSRIs (Thomas et al., 1987; Dechant and Clissold, 1991), which enhance 5-HT neurotransmission by increasing the extracellular 5-HT level (Invernizzi et al., 1995). SERT mRNA expression is detected in several brain areas, including the brain stem, hippocampus, and frontal cortex. In particular, the midbrain raphe complex shows higher expression levels than other areas (Bengel et al., 1997). Concerning the circadian change of SERT expression, 3H-labeled imipramine binding sites showed marked increase in suprachiasmatic nuclei of the anterior hypothalamus during the dark phase (Wirz-Justice et al., 1983).
Chronopharmacological studies of psychotropic drugs have been reported using animals, and trials for clinical application considering the dosing-time have already been undertaken (Nagayama, 1993). However, the mechanisms underlying the phenomenon in which the activity of psychotropic drugs, including antidepressants, varies depending on the dosing schedule are still unclear. The purpose of this study was to examine the 24-h rhythm of antidepressant activity in FST. The mechanisms underlying this phenomenon were also investigated from the perspectives of pharmacokinetics and pharmacodynamics.
Materials and Methods
Animals. Five-week-old male ICR mice were purchased from Charles River Japan (Kanagawa, Japan). Mice were housed eight to 10 per cage under standardized light/dark cycle condition (lights on at 7:00 AM, lights off at 7:00 PM) at a room temperature of 24 ± 1°C and humidity of 60 ± 10% with food and water ad libitum. All mice were exposed to the light/dark cycle for 1 week before the experiments. During dark periods, dim red light was used to minimize the light stimuli to mice.
Drugs and Treatment Procedures. The following drugs were used: amitriptyline hydrochloride (Sigma-Aldrich, St. Louis, MO) and fluvoxamine maleate (Solvay Pharmaceuticals Ltd., Weesp, The Netherlands). Both drugs were dissolved in sterilized saline, and the volume of injection was 0.01 ml/g body weight. In preliminary experiments, we examined whether the anti-immobility effect of antidepressants in FST showed any dose dependence. This experiment was performed at 9:00 AM, and the dosages were 5, 15, and 30 mg/kg for amitriptyline and 15, 30, and 60 mg/kg for fluvoxamine.
To investigate the influence of dosing time on anti-immobility effect, groups of six to 12 mice were injected intraperitoneally with saline, amitriptyline (15 mg/kg), or fluvoxamine (30 mg/kg) at 9:00 AM, 1:00 PM, 5:00 PM, 9:00 PM, 1:00 AM, or 5:00 AM. FST was performed at 30 min after the drug injection. To study the influence of fluvoxamine on locomotor activity, groups of five to six mice were injected intraperitoneally with saline or fluvoxamine (30 mg/kg) at 9:00 AM or 9:00 PM. To study the influence of dosing time on pharmacokinetics of fluvoxamine, groups of four to eight mice were injected intraperitoneally with fluvoxamine (30 mg/kg) at 9:00 AM or 9:00 PM. Blood and whole brain samples were obtained at 0.25, 0.5, 1, 2, or 4 h after the drug injection, and blood samples were centrifuged at 3000 rpm for 15 min to get the plasma. Both plasma and brain samples were stored at -80°C until assay. To clarify the time-dependent changes of SERT mRNA expression and 5-HT uptake activity in mouse midbrains, whole brain samples were removed from groups of five mice, and the midbrain was separated at 9:00 AM or 9:00 PM. For assay of 5-HT activity, crude synaptosomes were prepared at each time, and all samples were stored at -80°C until assay.
Forced Swimming Test. The procedure was performed based on a previous report (Porsolt et al., 1977). Briefly, mice were placed individually into plastic cylinders (height, 25 cm; diameter, 10 cm) containing 10 cm of water, maintained at 21 to 23°C, and left there for 5 min. A mouse was judged to be immobile when it floated in an upright position and made only small movements to keep its head above water. The duration of immobility was recorded during the 5-min testing period.
Monitoring Locomotor Activity. To assess locomotor activity, mice were housed individually in plastic cages (24 × 16 × 12 cm), and their locomotor activity was measured by area sensors (model NS-AS01) with a thermal radiation detector. The sensors were set at a 15-cm height from the cage floor. The data were stored on a personal computer. Activity counts (number of movements) were recorded at 1-min intervals. Activity was recorded 15 min before and 30 min after injection. For visualization of locomotor activity, activity counts were calculated using a 3-min window.
Determination of Fluvoxamine Concentration in Plasma and Brain. Measurement of plasma fluvoxamine concentration was performed as described previously (Wong et al., 1994). Then, 200 μl of plasma sample spiked with solution of clomipramine, as the internal standard, was alkalinized by addition of 200 μl of 1 N NaOH, followed by 1 ml of n-hexane/3-methylbutanol (99:1) and shaken for 30 min and centrifuged at 10,000 rpm for 10 min. The organic layer was transferred to another tube for acid back-extraction with 100 μl of 0.05% phosphoric acid, which was then shaken for 20 min and centrifuged at 10,000 rpm for 10 min. The top organic layer was discarded, and 25 μl of the remaining aqueous layer was injected into the HPLC system for analysis. To determine the brain fluvoxamine concentration, mouse whole brain was homogenized in 1 ml of methanol and centrifuged at 12,000 rpm for 15 min. The supernatant spiked with clomipramine solution was alkalinized by addition of 1 ml of 1 N NaOH, followed by 4 ml of hexane/3-methyl-butanol (99:1), and then was shaken for 30 min and centrifuged at 4500 rpm for 10 min. The organic layer was transferred to another tube for acid back-extraction with 200 μl of 0.05% phosphoric acid, which was then shaken for 20 min and centrifuged at 4500 rpm for 10 min. The top organic layer was discarded, and 25 μl of the remaining aqueous layer was injected into the HPLC system for analysis. The HPLC apparatus consisted of a LC-10AS liquid chromatograph (Shimadzu, Kyoto, Japan); a 5-μm particle size (4.0 mm i.d. × 10 mm) CAPCELL PAK C18 MG S-5 guard column (Shiseido, Tokyo, Japan); a 5-μm particle size (4.6 mm i.d. × 250 mm) CAPCELL PAK C18 MG analytical column (Shiseido); a SPD-10A UV-Vis detector (Shimadzu), set at 245 nm; and a C-R7A plus Chromatopac (Shimadzu). The column temperature was maintained at room temperature. The mobile phase was 50 mM KH2PO4 (pH 4.7 with KOH) and acetonitrile [60:40 (v/v)]. The mobile phase was filtered and degassed before use and the flow rate was 1 ml/min.
RNA Extraction and RT-PCR. Total RNA was extracted from mouse midbrains by using TRIzol solution (Invitrogen, Carlsbad, CA). Reverse transcription of RNA and cDNA amplification was performed with a one-step RT-PCR system (Invitrogen). The following oligodeoxynucleotide primers were used: sense primer 5′-ATCATAGCCTGGGCGCTCTAC-3′ and antisense primer 5′-CATGTAGCCAAGCACCGTGAA-3′ for SERT (GenBank accession no. AF013604) and sense primer 5′-GACCTCAACTACATGGTCTACA-3′ and antisense primer 5′-ACTCCACGACATACTCAGCAC-3′ for GAPDH (GenBank accession no. M32599). The PCR product was electrophoresed through a 3% agarose gel. The amounts of RT-PCR products of each mRNA were quantified using Kodak 1D image analysis software and normalized against GAPDH.
Preparation of Crude Synaptosome and 5-HT Uptake Assay. Crude synaptosome was prepared as described previously (Gray and Whittaker, 1962). Midbrain samples were homogenized by 0.32 M sucrose and centrifuged at 1000 rpm at 4°C for 10 min. The pellet was discarded and the supernatant was centrifuged at 12,000 rpm at 4°C for 20 min. The supernatant was discarded, and the pellet was resuspended in the original volume of 0.32 M sucrose used as crude synaptosome. 5-HT uptake assay was performed as described previously (Snyder and Coyle, 1969). Crude synaptosome (final concentration 0.25 mg of protein) was preincubated at 37°C for 5 min with Krebs-HEPES buffer (127 mM NaCl, 5 mM KCl, 1.3 mM NaH2SO4, 15 mM HEPES, 10 mM glucose, and 1.2 mM MgSO4). The buffer was gassed with oxygen for 30 min before use. Then, 3H-labeled 5-HT (final concentration 50 nM; GE Healthcare, Little Chalfont, Buckinghamshire, UK) was added and incubated at 37°C for 5 min followed by rapid filtration under vacuum (GF/B filters; Whatman, Maidstone, UK). The filters were washed three times with cold Krebs-HEPES buffer, dried, and placed in ACSII scintillation cocktail (GE Healthcare). Nonspecific uptake was calculated from data obtained incubation at 0°C.
Statistical Analysis. One-way ANOVA, two-way ANOVA, repeated measures ANOVA, and Scheffé's test were applied for the multiple comparison. The Student's t test was used for independent comparison between two groups. The 5% level of probability was considered to be significant. The population pharmacokinetic parameters were calculated on an HP-9000 series 700 (Yokogawa-Hewlett Packard Ltd., Tokyo, Japan) with the NONMEM program (version IV, level 1.1), following the one-compartment model (PREDPP program, subroutines ADVAN1 and TRANS1). Since pharmacokinetic parameters were analyzed by NONMEM, statistical significance was performed by χ2 test. NONMEM is widely used for fragmentary data such as a limited number of samples in clinical practice and drug concentrations in animal tissues.
Results
Dose-Dependent Manner of Anti-Immobility Effect of Antidepressants. The immobility time in the FST is shown in Fig. 1, a and b. This experiment was performed at 9:00 AM. The anti-immobility effect showed dose dependence for both antidepressant, and the immobility time was significantly decreased at doses of 15 and 30 mg/kg amitriptyline (P < 0.01) and 30 and 60 mg/kg fluvoxamine (P < 0.05 and P < 0.01, respectively). Therefore, 15 mg/kg amitriptyline and 30 mg/kg fluvoxamine were selected in the present study.

Dose dependencies of anti-immobility effects of amitriptyline (a) and fluvoxamine (b). This experiment was performed at 9:00 AM. The immobility time was significantly decreased at doses of 15 and 30 mg/kg amitriptyline and 30 and 60 mg/kg fluvoxamine. Each value is mean with S.E. of 8 to 12 mice. *, P < 0.05; **, P < 0.01 compared with saline group.
Influence of Dosing Time on the Anti-Immobility Effect of Antidepressants. The results are shown in Fig. 2, a and b. There was no significant interaction between the drug injection and the dosing time in Fig. 2, a and b. As for the effect of the drug injection, the immobility times of mice treated with amitriptyline and fluvoxamine were significantly decreased compared with those of mice treated with saline (F = 225.49, df= 1, P < 0.01 for Fig. 2a; F = 83.19, df = 1, P < 0.01 for Fig. 2b). The multiple comparison test was performed separated by dosing time, and the immobility times of mice treated with amitriptyline and fluvoxamine were significantly decreased compared with mice treated with saline at any six different times (P < 0.01 at 9:00 AM, 1:00 PM, 5:00 PM, 9:00 PM, 1:00 AM, and 5:00 AM for Fig. 2a; P < 0.05 at 5:00 PM and P < 0.01 at 9:00 AM, 1:00 PM, 9:00 PM, 1:00 AM, and 5:00 AM for Fig. 2b). As for the effect of dosing time, the immobility time showed a significant time-dependent change (F = 4.00, df = 5, P < 0.01 for Fig. 2a; F = 6.60, df= 5, P < 0.01 for Fig. 2b). The multiple comparison test was performed separated by saline or antidepressants, and the immobility time of mice treated with saline was decreased slightly during the dark phase but showed no significant 24-h rhythm. In contrast, the immobility time of mice treated with amitriptyline and fluvoxamine showed a significant 24-h rhythm (F = 3.29, df= 5, P < 0.05 for amitriptyline group; F = 6.27, df = 5, P < 0.01 for fluvoxamine group). The anti-immobility effect of antidepressants in FST was potent at the early part of the dark phase and weak during the light phase. In particular, immobility time when treated with fluvoxamine at 9:00 PM was significantly decreased compared with 9:00 AM, 1:00 PM, and 5:00 PM (P < 0.01).

Influence of dosing time on immobility time of mice injected with amitriptyline (a, 15 mg/kg i.p.) or fluvoxamine (b, 30 mg/kg i.p) at six different times in forced swimming test. Each value is the mean with S.E. of 6 to 12 mice. The immobility time of mice injected with amitriptyline and fluvoxamine showed a significant 24-h rhythm (F = 3.29, P < 0.05 for amitriptyline group; F = 6.27, P < 0.01 for fluvoxamine group). *, P < 0.05; **, P < 0.01 compared with saline group at the corresponding dosing time. ##, P < 0.01 compared with 9:00 PM of fluvoxamine group. ○, saline; ▪, amitriptyline; •, fluvoxamine.
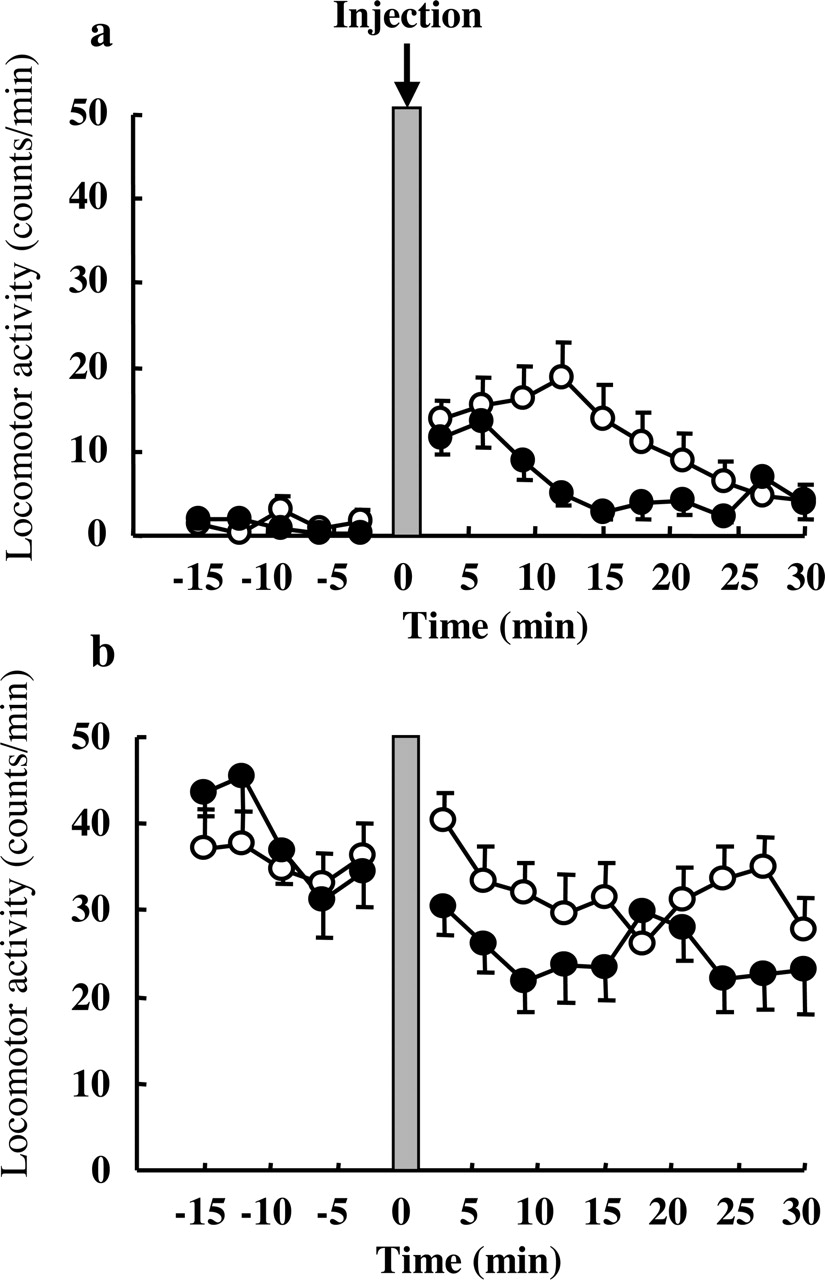
Influence of Dosing Time of Fluvoxamine on Locomotor Activity. The results are shown in Fig. 3, a and b. The basal locomotor activity was much higher at 9:00 PM (Fig. 3b), when antidepressants seemed to inhibit immobility most, than at 9:00 AM (Fig. 3a). The locomotor activity was slightly increased immediately after injection at 9:00 AM, but not at 9:00 PM. However, the activity counts after injection of fluvoxamine were not significantly different from those of saline at both 9:00 AM and 9:00 PM. Therefore, fluvoxamine showed no dosing time-dependent difference effect on locomotor activity.

Influence of dosing time on locomotor activity after saline or fluvoxamine (30 mg/kg i.p.) injection at 9:00 AM (a) or 9:00 PM (b). The activity counts (number of movements) were calculated using a moving average with a 3-min window. Each value is the mean with S.E. of five to six mice. No significant difference between two groups at either dosing time. ○, saline; •, fluvoxamine.
Influence of Dosing Time on Fluvoxamine Pharmacokinetics. The time courses of fluvoxamine concentration in plasma and brain are shown in Tables 1 and 2 and Fig. 4, a and b. There was no significant interaction between the time after the injection and the dosing time. As for the effect of dosing time, plasma concentration showed a significant time-dependent change (F = 5.46, df = 1, P < 0.05 for Fig. 4a), but brain concentration showed no significant time-dependent change. The multiple comparison test was performed separated by the time after the injection, and plasma concentration at 1 h after the injection at 9:00 AM was significantly higher than that at 9:00 PM (P < 0.05; Fig. 4a). Calculating pharmacokinetic parameters (Table 3), the Ke of fluvoxamine was 1.06 or 1.40 when injected at 9:00 AM or 9:00 PM, respectively. It was approximately 1.3-fold higher in mice injected at 9:00 PM than at 9:00 AM, and these two values showed significant difference (P < 0.01). However, the Vd of fluvoxamine was not different between injection at 9:00 AM and at 9:00 PM.
Influence of dosing time on plasma fluvoxamine concentration after fluvoxamine injection at 9:00 AM or 9:00 PM Each value is the mean with S.E. of six to eight observations.
Influence of dosing time on brain fluvoxamine concentration after fluvoxamine injection at 9:00 AM or 9:00 PM Each value is the mean with S.E. of four to eight observations. Statistical analysis by two-way ANOVA. As for the effect of the dosing time, brain concentration showed no significant time-dependent change.
Final estimates of population pharmacokinetic parameters
Time-Dependent Change of SERT mRNA Expression and 5-HT Uptake Activity in Mice Midbrain. SERT mRNA expression levels and 5-HT uptake activity in mice midbrains are shown in Fig. 5, a and b. The relative expression levels were normalized against GAPDH as an internal standard. The mRNA expression of SERT that SSRIs bind specifically was significantly higher at 9:00 PM than at 9:00 AM in mouse midbrains (P < 0.05). The 5-HT uptake activity was measured by an assay system using crude synaptosome. Preparation of crude synaptomes from mouse midbrains was performed at 9:00 AM or 9:00 PM. Associated with SERT mRNA expression level, 5-HT uptake activity in mouse midbrains was also significantly higher at 9:00 PM than at 9:00 AM (P < 0.01).
Discussion
In the present study, the immobility times of mice treated with amitriptyline (15 mg/kg) and fluvoxamine (30 mg/kg) were decreased compared with that of mice treated with saline. And the immobility times of mice treated with amitriptyline and fluvoxamine in the FST showed significant 24-h rhythms. From these results, the anti-immobility effect of antidepressants was potent at the early part of the dark phase and weak during the light phase. According to other reports, melatonin receptor antagonist luzindole (N-0774) reduces the immobility time in FST pronouncedly in the middle of the dark phase rather than in the light phase (Dubocovich et al., 1990). Behavioral profiles and responses in several tests vary under diurnal and nocturnal conditions (Kelliher et al., 2000; Bertoglio and Carobrez, 2002).
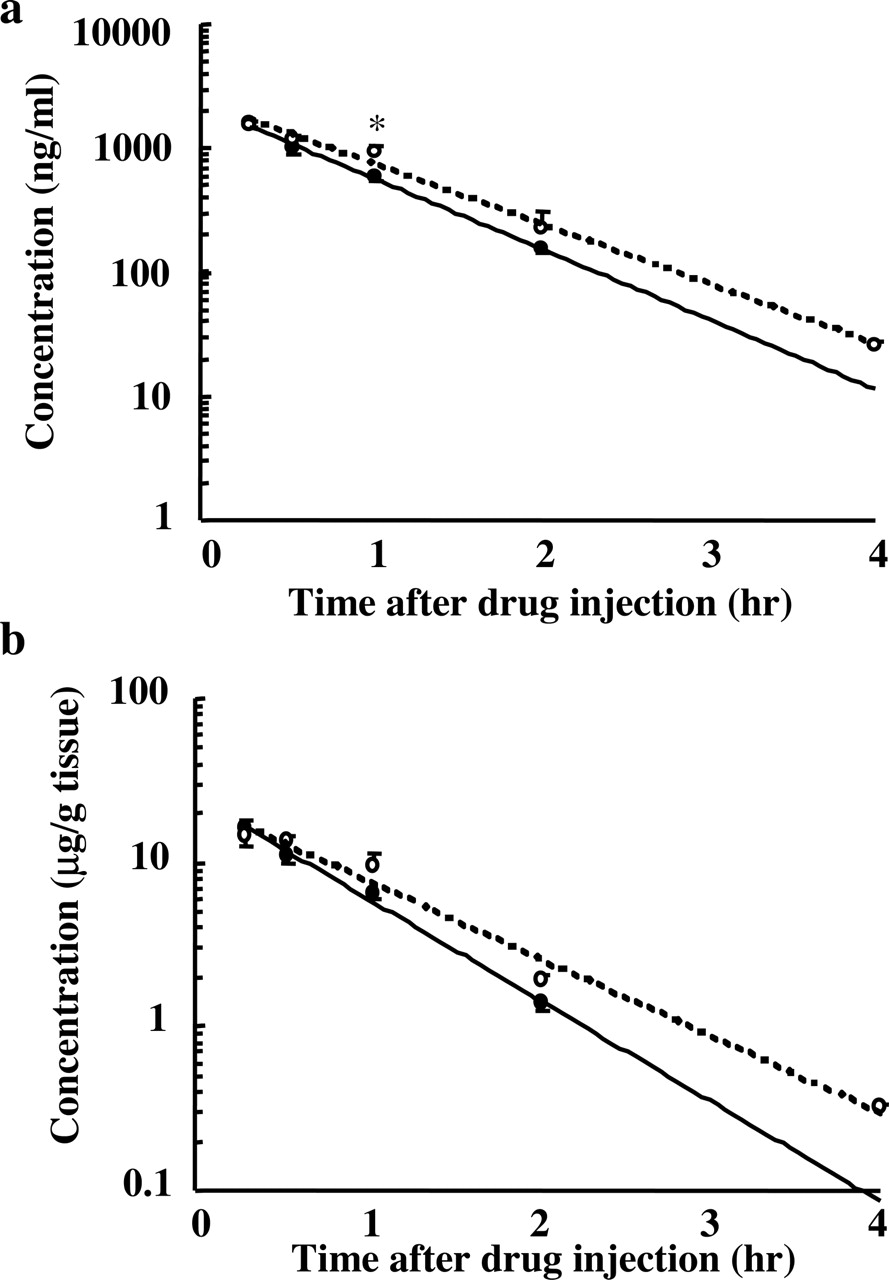
Influence of dosing time on plasma (a) and brain (b) fluvoxamine concentrations after fluvoxamine (30 mg/kg i.p.) injection at 9:00 AM (○) or 9:00 PM (•). Each value is the mean with S.E. of four to eight mice. *, P < 0.05 compared between two groups.
In FST, antidepressant effects could be clearly distinguished from the effects of psychostimulants, which markedly increased locomotor activity (Porsolt et al., 1978). Although the locomotor activity was increased during the dark phase rather than during the light phase, the immobility time of mice treated with saline showed no significant 24-h change. Furthermore, since fluvoxamine had no effect on the locomotor activity of mice at 9:00 AM and 9:00 PM, fluvoxamine seemed to decrease the immobility time in FST without increasing locomotor activity. FST was developed by Porsolt in rats and mice and is the most widely used tool for preclinically assessing antidepressant activity. However, FST is considered to be insensitive in evaluating the effect of SSRIs, since it is less effective for SSRIs than drugs inhibiting noradrenaline reuptake such as amitriptyline (Lucki, 1997). The present results suggest that FST is more sensitive to SSRIs by considering the dosing time. Then, we investigated the mechanisms underlying the dosing-time dependent anti-immobility effect of fluvoxamine from the perspectives of pharmacokinetics and pharmacodynamics.

Time-dependent changes of SERT mRNA level (a) and 5-HT uptake activity (b) in mouse midbrains. The relative expression levels were normalized against GAPDH. Each value is the mean with S.E. of five mice. Both mRNA expression and 5-HT uptake activity were significantly higher at 9:00 PM than at 9:00 AM. *, P < 0.05; **, P < 0.01 compared between two groups. Open column shows 9:00 AM group and shadowed column shows 9:00 PM group.
First, we examined the influence of dosing time on fluvoxamine pharmacokinetics. In the preset study, the plasma fluvoxamine concentration at 1 h after injection at 9:00 AM was higher than that at 9:00 PM. The Ke was approximately 1.3-fold higher in mice injected at 9:00 PM than at 9:00 AM. The major metabolic pathway of fluvoxamine is oxidative demethylation and oxidative deamination in the liver (Overmars et al., 1983), and fluvoxamine is metabolized by CYP1A2 and CYP2D6 in human (Carrillo et al., 1996; Spigset et al., 1997). The expression of CYP1A2 mRNA is observed in mice (Dey et al., 1999), but 24-h rhythms of mRNA expression and enzyme activity of mouse CYP1A2 have not been clarified yet. Although the exact mechanism underlying the dosing time dependence of fluvoxamine pharmacokinetics is not clear, a significant 24-h rhythm is demonstrated for hepatic blood flow (Lemmer and Nold, 1991). Therefore, this factor may be considered to be one of the mechanisms underlying the time dependence change of fluvoxamine pharmacokinetics. Nevertheless, both plasma and brain fluvoxamine concentrations at 0.5 h after the drug injection, when FST was performed, showed no significant difference between 9:00 AM and 9:00 PM. Consequently, the dosing time dependence in the anti-immobility effect of fluvoxamine does not seem to be related to the pharmacokinetics of the drug.
Second, we investigated the mechanism from the perspective of pharmacodynamics, focusing on the mRNA expression and uptake activity of SERT associated with the reuptake of 5-HT from the synapse cleft. Since high expression levels of SERT mRNA are detected in the mouse midbrain raphe complex, including dorsal and median raphe nuclei, it may be better to examine the mouse midbrain in detail. However, since only a small volume of RNA is obtained from each raphe nucleus, the whole midbrain complex was used in the present study. Expression levels of SERT mRNA in mouse midbrains showed a significant time-dependent change with higher level at 9:00 PM and lower level at 9:00 AM. In addition, 5-HT uptake activity in crude synaptosome prepared from mouse midbrains showed a significant time-dependent change associated with mRNA expression. These results suggest that the reuptake of 5-HT from the synapse cleft may be more increased during the dark phase than during the light phase.
According to previous reports, the extracellular 5-HT level in the dorsal raphe nucleus in freely moving rats is higher during the dark phase than during the light phase (Portas et al., 1998). And 5-HT neurotransmission in brain slices prepared from the rat hypothalamus shows time-dependent difference between the dark phase and the light phase (Blier et al., 1989). Furthermore, it is thought that the reuptake of 5-HT from the synapse cleft is inhibited almost completely by injection with 30 mg/kg fluvoxamine at any time (Claassen et al., 1977). Nevertheless, extracellular 5-HT levels could be further increased by the injection of fluvoxamine at 9:00 PM than at 9:00 AM. Therefore, the time-dependent change of extracellular 5-HT levels may contribute to the anti-immobility effect of fluvoxamine in FST. Since the anti-immobility effect of amitriptyline showed a significant 24-h rhythm, expression of norepinephrine transporter (NET) or activity of MAO enzyme might show a time-dependent change. However, since we focused on the mechanisms underlying the dosing time-dependent anti-immobility effect of fluvoxamine in the present study, we did not measure expression of NET or activity of MAO. Further study may be necessary to investigate whether expression of NET or activity of MAO shows a significant 24-h rhythm.
The time-dependent change of SERT mRNA expression might be affected by circadian change of neuronal and/or hormonal signals in the transcriptional stage. However, adrenalectomy does not affect [3H]citalopram binding in the rat midbrain (Kulikov et al., 1997) or [3H]imipramine binding in the frontal cortex (Arora and Meltzer, 1986). On the other hand, [3H]paroxetine binding to the rat cortical membrane under restriction conditions significantly reduces without any change of transporter affinity (Zhou et al., 1996). The exact mechanisms underlying the time-dependent change of SERT mRNA expression should be clarified.
In the present study, the anti-immobility effect of fluvoxamine in FST was increased depending on dosing time. In other words, the sensitivity of FST to SSRIs was improved by considering the dosing time. Furthermore, time-dependent change of SERT mRNA expression and uptake activity in the midbrain, probably producing the rhythmicity of extracellular 5-HT levels, is suggested to be the mechanism underlying 24-h rhythm of anti-immobility effect of fluvoxamine in FST.
Acknowledgments
We are indebted to Solvay Pharmaceuticals Ltd. for supplying the fluvoxamine used in this study.
Footnotes
-
Article, publication date, and citation information can be found at http://jpet.aspetjournals.org.
-
doi:10.1124/jpet.105.088849.
-
ABBREVIATIONS: 5-HT, 5-hydroxytryptamine (serotonin); FST, forced swimming test; SERT, serotonin transporter; SSRI, selective serotonin reuptake inhibitor; HPLC, high-performance liquid chromatography; RT-PCR, reverse transcription-polymerase chain reaction; ANOVA, analysis of variance; NONMEM, nonlinear mixed effect model; NET, norepinephrine transporter; MAO, monoamine oxidase.
- Received April 29, 2005.
- Accepted August 1, 2005.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}