An Update on Canine Adenovirus Type 2 and Its Vectors

1
Institut de Génétique Moléculaire de Montpellier, CNRS UMR 5535, 1919 Route de Mende Montpellier, 34293 France
2
Université de Montpellier I, 5 Bd Henri IV, 34000 Montpellier, France
3
Université de Montpellier II, place Eugène Bataillon, 34090 Montpellier, France
*
Author to whom correspondence should be addressed.
Viruses 2010, 2(9), 2134-2153; https://doi.org/10.3390/v2092134
Submission received: 25 June 2010
/
Revised: 23 September 2010
/
Accepted: 24 September 2010
/
Published: 27 September 2010
(This article belongs to the Special Issue Adenoviral Vectors)
{kind=link}
{kind=link}
{kind=link}
Abstract
:Adenovirus vectors have significant potential for long- or short-term gene transfer. Preclinical and clinical studies using human derived adenoviruses (HAd) have demonstrated the feasibility of flexible hybrid vector designs, robust expression and induction of protective immunity. However, clinical use of HAd vectors can, under some conditions, be limited by pre-existing vector immunity. Pre-existing humoral and cellular anti-capsid immunity limits the efficacy and duration of transgene expression and is poorly circumvented by injections of larger doses and immuno-suppressing drugs. This review updates canine adenovirus serotype 2 (CAV-2, also known as CAdV-2) biology and gives an overview of the generation of early region 1 (E1)-deleted to helper-dependent (HD) CAV-2 vectors. We also summarize the essential characteristics concerning their interaction with the anti-HAd memory immune responses in humans, the preferential transduction of neurons, and its high level of retrograde axonal transport in the central and peripheral nervous system. CAV-2 vectors are particularly interesting tools to study the pathophysiology and potential treatment of neurodegenerative diseases, as anti-tumoral and anti-viral vaccines, tracer of synaptic junctions, oncolytic virus and as a platform to generate chimeric vectors.
1. Introduction
Adenoviridae infect a variety of vertebrates including mammals, fish, birds and reptiles [1]. More than 50 human adenoviruses (HAd) serotypes have been identified. A handful of these are extensively studied because of their tendency to induce respiratory, ocular, and enteric infections in immunocompetent individuals as well as morbidity and mortality in severely immunodeficient patients [2,3,4,5,6]. Early generations of HAd vectors, primarily ΔE1/E3, are often efficient for a limited duration in laboratory animals that have never been exposed to the wild type viruses. However, subsequent deliveries lead to inefficient gene transfer. The poor efficacy and limited duration are mainly due to neutralizing antibodies that prevent transduction and to CD8+ and CD4+ cytotoxic T lymphocytes (CTLs) that destroy transduced cells. This memory immune response can also be problematic for clinical use of vectors. Decades ago, epidemiology studies reported that the majority of us had been exposed to multiple adenovirus serotypes by the time we were 10 years old [7,8,9]. This leads to a cross-reacting humoral, as well as cellular response against many serotypes [10,11,12]. To reduce or circumvent these drawbacks, strategies including the induction of immunotolerance, immunosuppression, or chemical and genetic modifications on viral vectors have been considered. In the early 1990s, we and others considered the possibility of using nonhuman vectors. We began designing canine adenovirus serotype 2 (CAV-2) vectors [13,14,15,16,17], while others were generating vectors from ovine, bovine, chimpanzee, simian or porcine [18,19,20,21,22]. We, and surely others, hypothesized that vectors derived from nonhuman adenoviruses would be more clinically useful than those from HAds based, in part, on the potential lack of immunological memory [23]. In spite of the approx 100,000 years of cohabitation of humans with dogs, CAV-2 has not been known to cross the species barrier and is unable to replicate in human cells. Therefore, our hope was to keep the numerous advantages associated with HAds vectors, while bypassing some of their clinical disadvantages.
2. Vectors
Plasmids harboring first-generation CAV-2 vector genomes are created by in vivo homologous recombination in E. coli BJ5183 between pTG5412, which contains the full-length CAV-2 genome, and a fragment of DNA containing the inverted terminal repeat (ITR), a transgene expression cassette and the CAV-2 protein IX and E2B coding regions [15]. This strategy is similar to that described initially by Chartier et al. [24]. Recombination gives rise to the formation of a plasmid harboring the E1-deleted CAV-2 genome. To generate the vector, CAV-2 E1-expressing cell lines derived from dog kidney cells such as DKCre and DKZeo cells are transfected and the vector serial amplified [15,25].
The latest and most efficient adenovirus vectors for long-term gene transfer are referred to as “helper dependent” (HD) and are deleted in all viral coding regions [17,26,27,28,29]. To generate the HD CAV-2 vector, an initial subcloning step consisted of the insertion of the transgene expression cassette in a plasmid called “pGut” containing a CAV-2 ITR and an overlapping region with a plasmid called pEJK25 (see below) [17]. Once a modified pGut is generated, it is linearized and subjected to homologous recombination in E. coli BJ5183 with common regions present on pEJK25, which contains two ITR’s, a packaging signal and 25 kb of stuffer sequence. Like HD HAd vectors, HD CAV-2 vectors are amplified by the co-replication and preferential packaging via a Cre recombinase induced-packaging-defective helper vector which supplies the regulatory and structural proteins in trans (see review by Philip Ng in this issue). HD vectors improved the efficiency and duration of transgene expression mainly due to the elimination of the adaptive cell-mediated immune response in immunologically naïve animals. HD CAV-2 vectors have a high cloning capacity (~30 kb), and allowed long-term (>1 year) transgene expression in the immunocompetent rat brain without immunosuppression [17].
3. CAV-2 tropism
Following the generation of CAV-2 vectors, the next challenge was to define what they would transduce in vitro and in vivo. Similar to HAd5, CAV-2 causes an upper respiratory track disease in dogs called kennel cough [30]. Moreover, anti-CAV-2 antibodies have also been found in foxes, bears and pandas [31], suggesting that either there is a cross-reacting humoral immunity to Ads that infect these species or that CAV-2 also naturally propagates in other hosts. Interestingly, CAV-2 has also been detected in the CNS of dogs and foxes.
The gene transfer field long ago moved away from the dogma that vector tropism is restricted to tissues that manifest disease symptoms in wild type infections. Via specific delivery routes (intravenous, intranasal, intracerebral, intramuscular, etc.), one can partially dictate the population of cells available for transduction, and therefore the tropism. Using intracerebral and intramuscular injections, we found that CAV-2 vectors preferentially transduce neurons [16]. When CAV-2 vectors were instilled in the rat olfactory cavity, the sensory olfactory neurons, in contrast to the columnar epithelial cells, were preferentially transduced. Injection in the hind leg muscle (gastrocnemius) and tibias anterior in newborn mice showed a poor level of transduction of the myofibers, but a surprising level of specificity for the innervating motor neurons via retrograde axonal transport. Similarly, following injection into the highly innervated diaphragm, an insignificant level of muscle cells were transduced but a significant number of neuromuscular junctions were GFP+, demonstrating again the transduction of motor neurons and axonal retrograde transport of CAV-2.
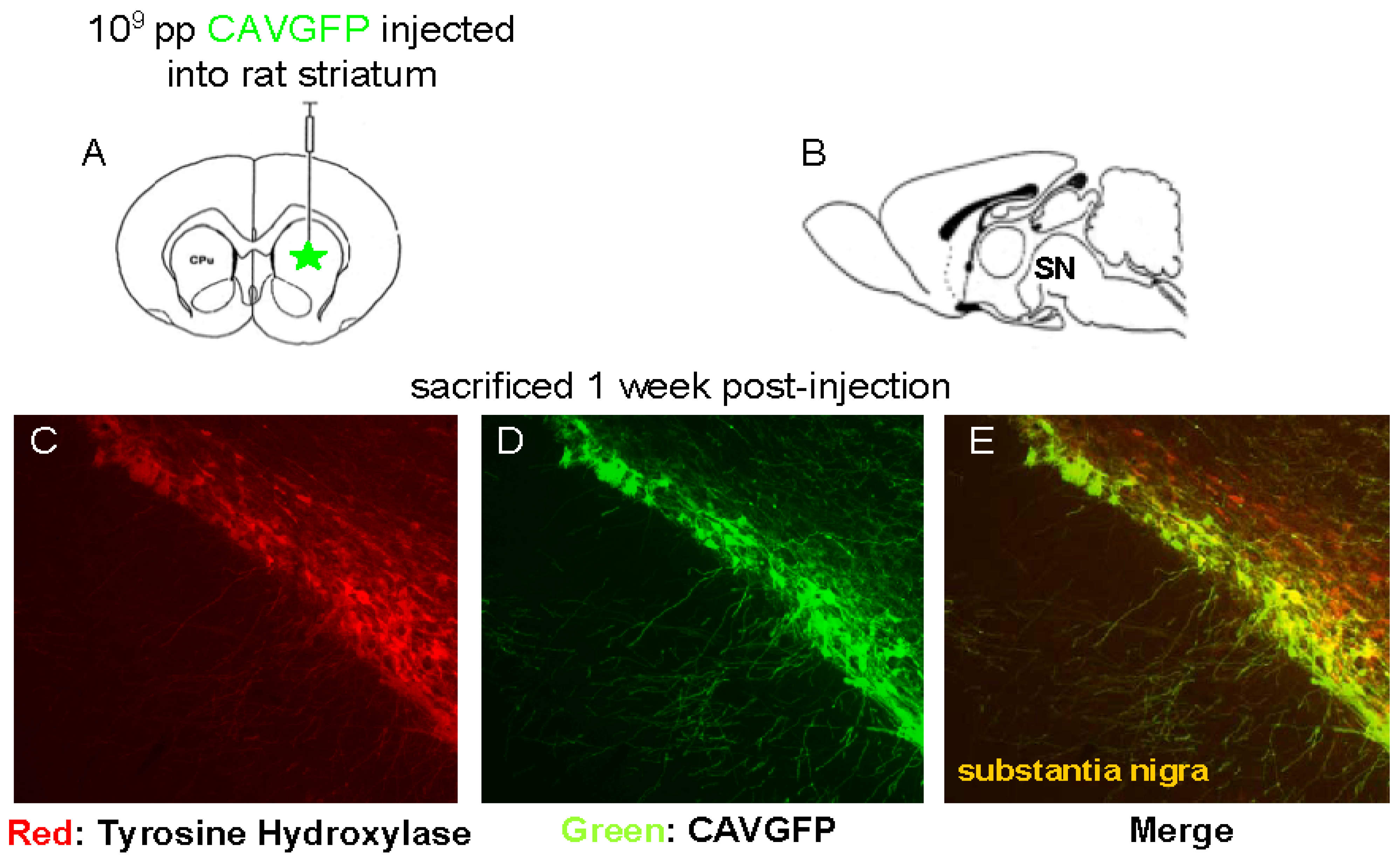
When injected into the brain parenchyma, CAV-2 vectors preferentially transduced neurons at the site of injection, as well as the neurons that project into this structure. Following injection in the striatum, the dopaminergic neurons of the substantia nigra pars compacta, neurons in layer IV of the neocortex and thalamus were transduced. We found that, via a single injection of 109 pp in the striatum of rats, CAV-2 vectors can transduce >70% of the dopaminergic neurons in substantia nigra pars compacta (Figure 1).
Not surprisingly, little was known concerning the determinants for CAV-2 long-range axonal transport. Axonal transport is essential for neuronal homeostasis as its impairment is often associated with neurodegenerative disorders. Some viruses, including rabies, herpes simplex type I (HSV-1) and poliovirus, as well as bacterial toxins such as tetanus toxin, use this process to access the nervous system [32]. These microbial agents can enter at nerve terminal situated in the periphery and use axonal transport to access the central and peripheral nervous systems. Notably, different mechanisms of axonal transport have been described: direct interaction with molecular motors for HSV-1 [33] versus endosomal trafficking for poliovirus [34] and tetanus toxin.
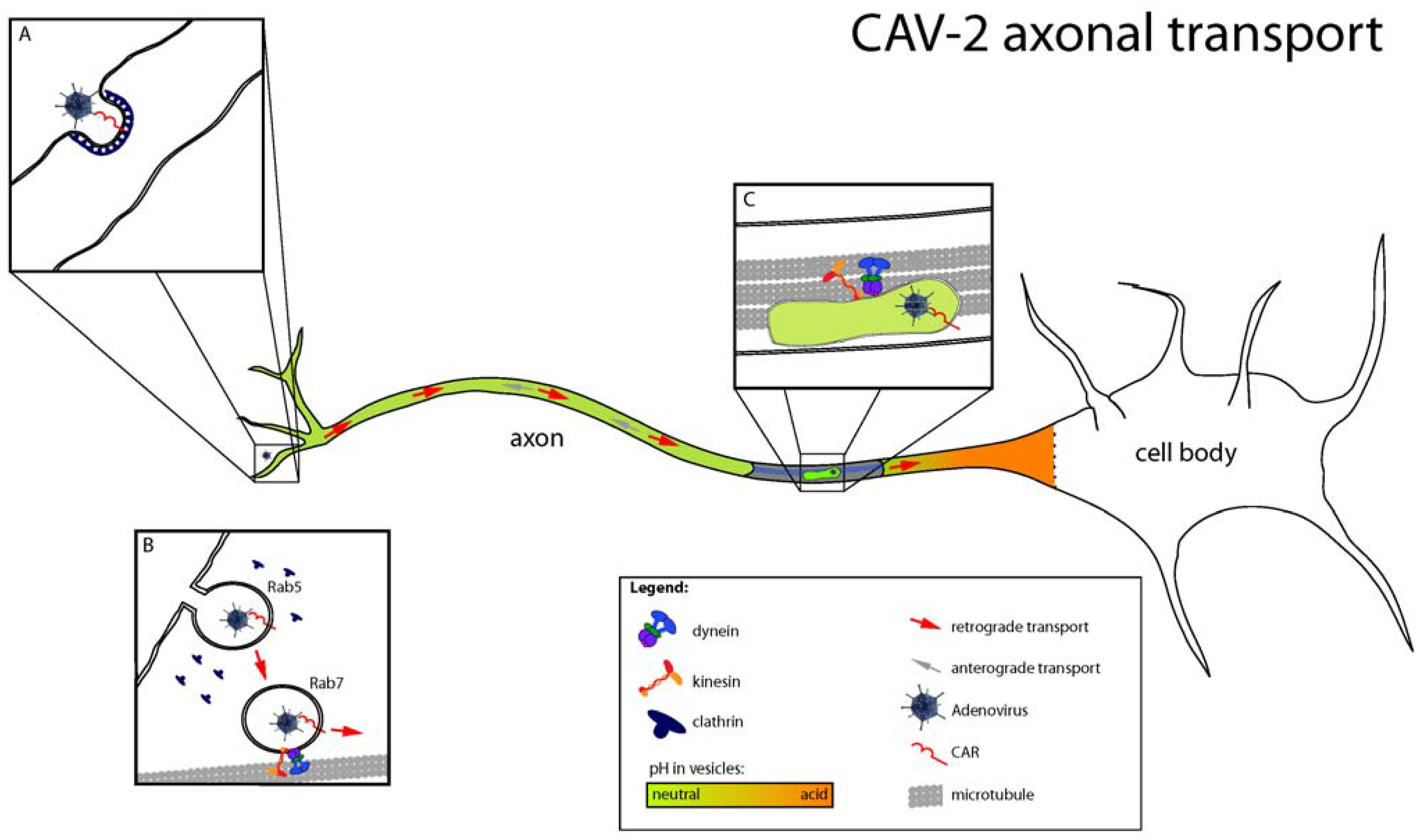
Using a combination of ultrastructural analyses, live-cell imaging and cell biology approaches, we recently characterized the molecular mechanisms regulating CAV-2 entry and transport in primary rodent motor neurons [35,36]. CAV-2 is retrogradely transported in motor neuronal axons of the sciatic nerve after intramuscular injection in hind leg of mice or primates. In vitro, we found that CAV-2 trafficking was bidirectional with a preferential retrograde transport, suggesting coordination between molecular motors of different polarities. This was confirmed by genetic inhibition of dynein (responsible for the retrograde transport from nerve terminals to cell bodies) and kinesin (responsible for the anterograde transport from cell bodies to nerve terminals), which led to a similar impairment of CAV-2 axonal transport. Ultrastructural analyses and live-cell imaging using fluorescent markers of endocytic organelles showed that CAV-2 transport was occurring inside endosomes, which matured from Rab5 to Rab7 during entry and transport (Figure 2). In epithelial cells, some serotypes of adenoviruses escape from Rab5 early endosomes due to acidification and conformational changes in the viral capsid that trigger membrane lysis [37,38]. Once in the cytoplasm, viruses may recruit dynein directly through the hexon protein to be transported to the nuclei [39]. To understand why CAV-2 is transported inside endosomes during its transport, we measured the pH of axonal endosomes using carboxyfluorescein-coupled virions. We found that CAV-2 was transported in endocytic organelles whose lumens were close to pH neutral. This vesicle likely allows long-range transport in a “protective” environment, precluding degradation or pH-induced conformational changes of the capsid and endosomal escape. Interestingly, this vesicular pathway is also used by tetanus toxin, poliovirus, and endogenous molecules such as neurotrophins and their receptors, suggesting that CAV-2 is taking advantage of an endogenous pathway crucial in neuronal differentiation and survival [32].
To gain further insight into how CAV-2 accessed this endocytic trafficking pathway, we characterized the role of Coxsackievirus and adenovirus receptor (CAR) [15,40] in the entry and transport of CAV-2. Whereas CAR’s function is partially described in tight-junctions of epithelia, no clear role in neurons has been described [41]. Notably, CAR is highly expressed during development of the nervous system and is also found in the adult nervous system [41,42,43]. Using competition experiments, we found that CAR was necessary for CAV-2 binding to the neuronal membrane and was also co-internalized with virions. Using fluorescently labeled CAV-2 fiber knob, we showed that CAR could enter and be transported bi-directionally in axons similar to CAV-2. These unsuspected vesicular dynamics of CAR opens numerous questions regarding its role in neuronal adhesion and possibly axonal transport. Together, these data suggest that CAV-2’s ability to infect neurons and be retrogradely transported could be due to the innate ability of CAR to access this trafficking pathway.
These data do not mean that CAV-2 vectors transduce exclusively neurons. Not surprisingly, CAV-2 also efficiently transduce epithelia of the upper and lower respiratory tract and alveolar cells after deep nasal instillation in mice [15,44]. In addition, one must keep in mind possible differences between “mice and men”. Although CAV-2 vectors may transduce certain murine tissues, this does not, a priori, mimic the efficiency of transduction in the clinic. We also assayed CAV-2 transduction ex vivo in temporal lobectomy specimens from epileptic patients and found preferential infection of neurons was not specific to rodents [16].
4. CAV-2 receptors
Direct observation of CAV-2 transduction was the first step to determine vector tropism in vitro and in vivo. Then, it was necessary to understand how CAV-2 interacts with cells at the molecular level.
Adenovirus entry into the cytoplasm can be functionally divided into attachment, internalization and permeabilization of the endosomal membrane. CAV-2, like all other adenoviruses, has an icosahedral capsid with the external surface composed mainly of hexon, penton base, and fiber. The fiber is an elongated thread-like molecule that projects from the penton base and initiates binding to the cellular surface. The C-terminal fiber knob domain of many Ads, such as HAd2/5, attaches to CAR [45,46,47]. Attachment of the virus is followed by internalization and permeabilization in clathrin-coated pits implicating dynamin [48], and αvβ5 and αvβ3 integrins [49]. The αv integrins recognize a conserved Arg-Gly-Asp (RGD) motif found in the adenovirus penton base and some extracellular matrix proteins. The three-dimensional structure of a recombinant soluble αvβ5 integrin bound to the penton base of HAd2 revealed a 20-Å RGD-binding cleft in the globular domain [50]. The crystal structure of the adenovirus fiber knob in complex with CAR [41] and a mutational analysis [51] identified several amino acids in the knob of some adenoviruses that are critical for CAR binding [52,53].
Figure 2.
Different steps of CAV-2 axonal transport. (A) After binding to CAR at the cell surface, CAV-2 is internalized in clathrin-coated pits. (B) During entry, maturation of CAV-2 containing vesicles occurs, with a shift from Rab5 to Rab7 endosomes that will eventually lead to CAV-2 axonal transport. Despite Rab7 maturation, the pH of axonal endosomes stays neutral, and CAV-2 remains inside vesicles during transport. (C) CAV-2 traffic involves dynein and kinesin, with a bias for the retrograde direction. CAR is still found in CAV-2 positive vesicles. (Image modified from Henaff and Salinas, Virulence [36]).
Figure 2.
Different steps of CAV-2 axonal transport. (A) After binding to CAR at the cell surface, CAV-2 is internalized in clathrin-coated pits. (B) During entry, maturation of CAV-2 containing vesicles occurs, with a shift from Rab5 to Rab7 endosomes that will eventually lead to CAV-2 axonal transport. Despite Rab7 maturation, the pH of axonal endosomes stays neutral, and CAV-2 remains inside vesicles during transport. (C) CAV-2 traffic involves dynein and kinesin, with a bias for the retrograde direction. CAR is still found in CAV-2 positive vesicles. (Image modified from Henaff and Salinas, Virulence [36]).

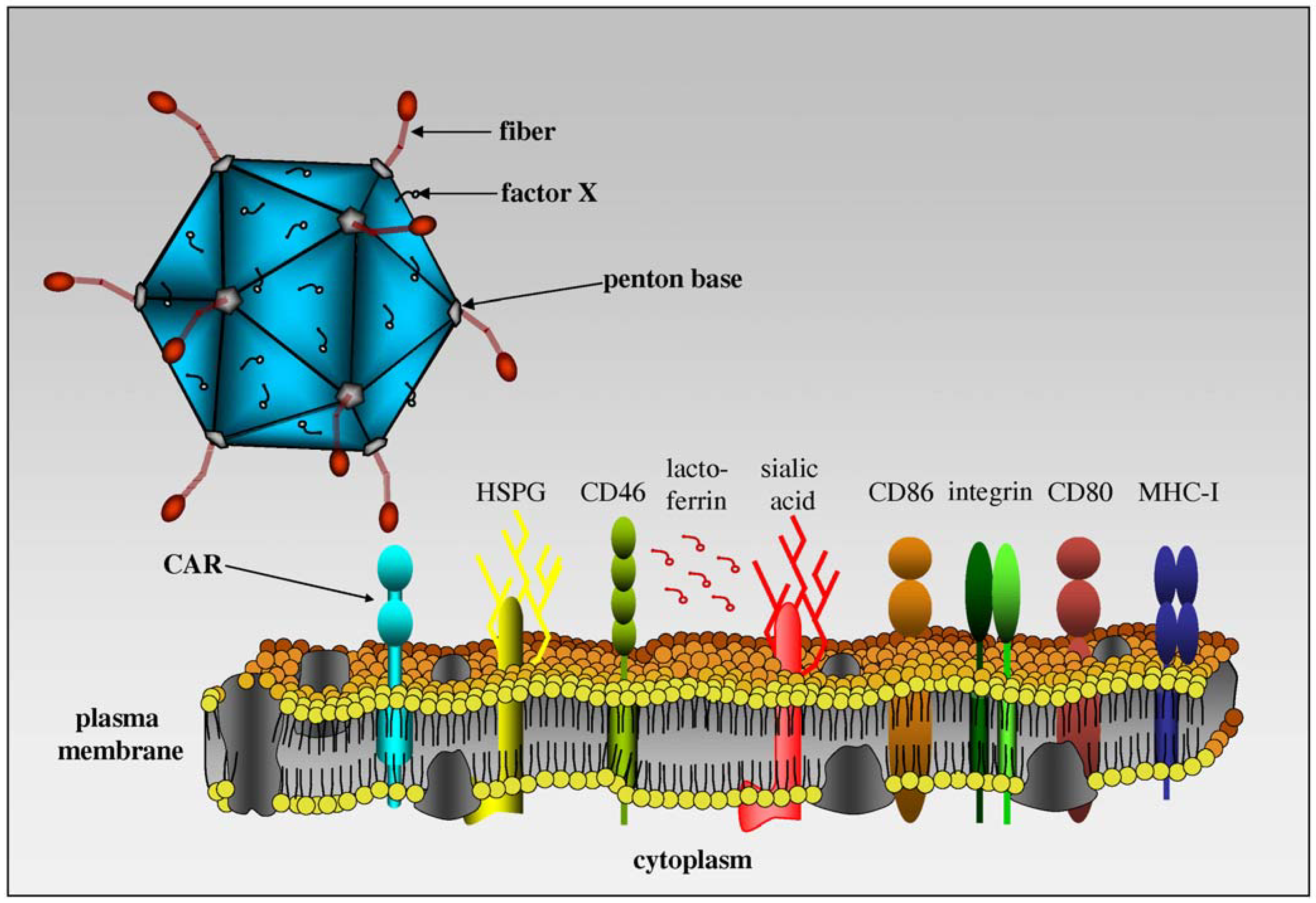
Our in vitro and in vivo data demonstrated that CAV-2 and HAd2/5 had overlapping, but distinct, tropism [15]. Essentially, we found that any cell or tissue that can be transduced by CAV-2 can also be transduced by HAd5, but not vice versa. Notably, cells expressing CAR could be transduced by CAV-2. CAR-negative cells but αMβ2 integrins and/or the heavy chain of MHC-I positive could not be transduced [40,54] (Figure 3). Interestingly, the CAV-2 capsid does not contain an integrin-interacting motif - in particular the RGD motif in the penton base loop [40,55]. Using radiolabeled CAV-2, we found that CAV-2 binds to CAR, and HAd5 fiber partially blocked attachment, suggesting that CAV-2 bound CAR at or near the same epitope. Preliminary data also suggested that CAV-2 does not interact with coagulation factor [56], lactoferrin [57] and CD46 [58,59].
A comparison of the three-dimensional structures of CAV-2 and HAd5 capsid allowed us to identify differences between these two viruses. We found that CAV-2 capsid has a smoother structure than human serotypes. Many of the external loops found in the HAd5 penton base and the hexon, against which the antibody response is directed, are shorter or absent in CAV-2, which could explain the paucity of cross-reacting human neutralizing antibodies [61,62]. On the other hand, the fiber appeared to be more complex. The CAV-2 fiber shaft is similar in length to that of HAd2, with a left-handed triple helical structure composed of β-strands interspersed with extended loops which contain 18.5 repeats with between 15 and 19 amino acids per repeat. A key difference may be the presence of two bends in the shaft [61]. Another major difference in the 3D structure of these viruses is located in protein IX. Its C-terminal part is in a different position, creating an antenna sticking out of the CAV-2 capsid. Protein IX could be a site to position additional protein domains for specific interaction with host cells.
5. Induced and crossreacting pre-existing immunity in humans
Twenty years ago, our initial reason to generate a nonhuman adenovirus vector was to avoid the inhibitory effects of the pre-existing humoral immunity readily found against multiple HAds serotypes, as well as to avoid the re-stimulation of memory CTLs. Due to multiple infections during childhood, most of us (>85%) possess relatively high circulating Abs against several HAd serotypes [8,63]. Anti-adenovirus antibodies can recognize many capsid proteins and, in particular, hexon, penton base, and fiber proteins [64,65]. Although the neutralization activity is often described as serotype specific [66] and age-related, cross-reacting nonneutralizing anti-HAd Abs also exist. Although there are ways to eliminate anti-capsid antibodies from serum, we would argue that a limited number of interventions to treat a patient should improve the chance for an encouraging outcome. In addition, transient depletion of antiviral antibodies from a patient could put them at risk for HAd disease from latent HAds [67]. Following the generation of CAV-2 vector, we quantified neutralizing Abs (NAbs) in serum from blood bank donors. We found that sera from approximately 98% of a random cohort did not contain significant titers of neutralizing anti-CAV-2 Abs [15]. These data supported the potential clinical use of CAV-2 because the donors were infected with multiple human serotypes, developed complex and varied responses and had different genetic background. Interestingly, similar observations have been made with ovine (OAd), bovine (BAd) and porcine (PAd) adenoviruses showing that anti-OAd, anti-BAd-3 and -PAd-3 NAbs were rare in the human population but that pre-existing humoral and cellular immunity could cross-react [10,68,69,70,71]. In some case, in vitro neutralization assays may not reliably predict the effect of virus-specific antibodies in vivo. Recently, assays compared the neutralization effect of human antibodies directed against wild type chimpanzee adenovirus 68 (AdC68) [72]. Human anti-HAds antibodies failed to neutralize a mutant form of AdC68 in vitro, containing a 3-amino acid mutation within the major neutralization site, but impaired the vector’s capacity to transduce cells and to stimulate a transgene product-specific CD8+ T-cells response in vivo.
NAbs are only one obstacle to efficient gene transfer. Another aspect of the pre-existing immunity includes the cellular response, in particular the long-lived proliferative CD4+ memory cells (TM). We and others have questioned the clinical potential of serotype switching of HAd vectors because a HAd5-induced cytotoxic TM can lyse autologous cells infected with other HAd species [73]. We predicted [23] that a TM response against virion proteins, which would be poorly blunted by many immunosuppressive drugs [74], would lead to deleterious side effects in some patients [14]. We assayed CAV-2-induced human TM proliferation and activation [75]. Fewer than half of the cohort harbored proliferating CD4+ TM directed against the CAV-2 virion proteins (versus >85% against HAd5 vectors). Furthermore, the CAV-2 responders had a 17-fold lower activation than the HAd5 vector responders and no CD8+ TM were detected in any donors.
In immunologically naïve rodents, CNS and respiratory tract delivery of ΔE1 CAV-2 vectors were less immunogenic than ΔE1/E3 HAd5 vectors and induced fewer infiltrating CD4+ and CD8+ cells at an equivalent number of injected particles [17,44]. Following intranasal instillation in mice, CAV-2 vectors also led to a lower level of TNF-α secretion than HAd5 vectors [44]. These data could be a result of a potential lack of capture, presentation and maturation of dendritic cells (DC) by CAV-2 [76].
Another issue that we explored was the direct interaction of the CAV-2 capsid with DC, which play a pivotal role in orchestrating and bridging innate, adaptive, and memory immunity [77,78]. Novel subtypes of DCs are also continuously being identified, ranging from Langerhans cells (myeloid) to plasmacytoïds (lymphoid) [79]. Each subtype can be artificially divided into immature or mature, which are characterized by phenotypically and functionally distinct characteristics. Immature DC sense their environment via non-specific phagocytosis and detect pathogens via evolutionarily conserved pattern recognition receptors, such as Toll-like receptors that recognize conserved microbe-associated molecules called “pathogen-associated molecular pattern” [80]. Activation promotes DC maturation, which results in the loss of their ability to take up antigens, change their morphology, and migrate towards the lymphoid compartments. Once there, matured DC are primed for antigen-specific naïve T-cell presentation and stimulation via the expression of major histocompatibility complex (MHC) class I/II and costimulatory molecules [81].
In the context of assessing the clinical relevance of CAV-2 vectors, we assayed their effect on human monocyte-derived DC (MoDC). Using a mix of functional and phenotypical assays, we found that, in contrast to the HAd5-based vectors, CAV-2 poorly transduced DC, provoked minimal upregulation of major histocompatibility complex class I/II and costimulatory molecules (CD40, CD80, and CD86), and induced negligible morphological changes indicative of DC maturation [76]. We also tested functional criteria for vector-induced MoDC maturation: reduction of antigen uptake, pertinent cytokine secretion (TNF-α, interleukin (IL)-10, IL-12, and type 1 interferon (IFN)). Again, in contrast to the three HAd5-based capsids, CAV-2 poorly induced functional characteristics of DC maturation. These results provided a partial explanation for the reductive adaptive immune response against CAV-2 vectors in rodents: poor DC transduction and maturation lead to a lower adaptive response in naïve hosts.
Together, these results showed that the CAV-2 vectors generated rare events of proliferation, activation or differentiation of T cells and also poorly transduced or induced the functional maturation of dendritic cells [10]. This may make CAV-2 vectors safer and more clinically applicable when long-term transgene expression is needed. Lack of vector-induced DC maturation may also limit an adaptive immune response following in vivo gene transfer. The lack of DC maturation was also seen when CAV-2 was incubated with human sera containing antibodies that recognize CAV-2 [82].
6. Potential uses
The preferential transduction of neurons and the high level of retrograde transport make CAV-2 vectors ideal tools to study the pathophysiology of many neurodegenerative disorders and to map complex neuronal networks in vivo. CAV-2 vectors are used in several studies to transfer genes encoding transcription factors, dominant negative mutants, constitutively activated kinases, etc., into neurons in vitro and in vivo [83,84,85,86]. Other potential uses include vaccines with overexpression of viral proteins such as rabies virus glycoprotein or cancer therapy [87,88,89].
6.1. Neurodegenerative diseases
After injection in the brain parenchyma, CAV-2 vectors preferentially target neurons and are efficiently retrograde transported into afferent brain regions. The transduction of the substantia nigra pars compacta following intrastriatal injection clearly suggests that these vectors may be well suited for the study and possible therapy of Parkinson’s disease. CAV-2 vectors could be used to modify a panel of gene expression in neurons by using either cell-specific promoters controlling a transgene of interest or encoding siRNAs that could potentially knockout a protein. For instance, one could analyze the effect of localized expression of neurotrophic factors such as ciliary or brain-derived neurotrophic factors, on Alzheimer’s, Huntington’s and many other neurodegenerative disorders.
CAV-2 vectors have been used to study the role of dopamine on fundamental behavior such as movement, feeding, reward responses and learning [83]. Dopamine-deficient (DD) mice were generated to allow a selective restoration of normal dopamine signaling to specific brain regions. These DD floxed stop mice had a non-functional tyrosine hydroxylase (TH) gene due to insertion of a NeoR gene flanked by loxP sites in the first intron of the TH gene. Injection of a CAV-2 vector expressing Cre recombinase in the central caudate putamen restored normal TH gene expression to the midbrain dopamine neurons. Transduced dopaminergic neurons expressed normal cellular proteins (TH) for >1 year post-transduction [83]. In another study, early-born mouse neurons could i) be transduced by CAV-2 vectors in organotypic slices, ii) migrate to distal regions, iii) display a voltage-gated sodium current, iv) express functional receptors, and v) show postsynaptic events. These data demonstrated that E1-deleted CAV-2 vector internalization do not significantly disrupt the normal physiology of differentiating neural progenitor cells [84].
Recently, CAV-2 vectors have been tested for their ability to improve neuropathological changes associated with the lysosomal storage disorder, mucopolysaccharidsosis (MPS) type IIIA. MPS’s are a group of lysosomal storage disorders that arise from deficiencies in the catabolism of glycosaminoglycans. At present, there are 11 known MPS, each resulting from the delivery of a different lysosomal enzyme. Of the MPS, MPS IIIA is one of the most common, and as far as treatment goes, one of the most intractable with symptoms including neurocognitive decline, hyperactivity and aggressive behavior. Direct injection of recombinant N-sulfoglucosamine sulfohydrolase (SGSH) into the brain parenchyma or cerebrospinal fluid of MPS IIIA mice improved many of the neuropathological features of the disease [90,91]. However, one of the major limitations of enzyme replacement therapy is the requirement for repeated administration. In contrast, an alternative strategy such as gene therapy is likely to generate persistent transgene expression. In MPS I, III and VII, the respective lysosomal enzymes can be secreted, then internalized by neighboring cells using the mannose-6-phosphate receptor. Because of this phenomenon, the number of cells that need to be genetically modified may be low but scattered throughout the brain. This pathophysiology is the reason CAV-2 vectors are particularly attractive. Recently, an E1-deleted CAV-2 vector expressing SGSH was intracerebrally injected in MPS IIIA mice [86]. Neonatal administration produced both dose-dependent and wide-spread transgene expression which persisted for at least 20 weeks and was sufficient to normalize memory and learning deficits in these mice. In contrast, injection of the same vector into the thalamus and ventricles of adult MPS IIIA mouse brain resulted in dose-dependent, but relatively short-lived gene expression. This loss of expression was likely due to the efficient transduction of ependymal cells in the ventricles during stereotaxic injections. Ependymal cells are efficient antigen presenting cells that, in turn, induced immune response against transduced cells. Besides the use of immunosuppressive drugs to improve the duration of transgene expression in adult mice, a more clinically relevant approach would be to use helper-dependent vectors.
6.2. Cancer therapy
Oncolytic conditionally-replicating adenoviruses (CRAds) are a class of anti-cancer agents with therapeutic potential [92]. In this approach, viruses selectively replicate in cancer cells and lead to destruction of the infected cells by virus-mediated cytolysis (CRAds are covered in the review by Toth et al. in this issue). Briefly, several oncolytic viruses have been created. Many anticancer strategies are based on the well-studied HAd5 system. To date, clinical efficacy observed in human trials has failed to reach the expectations that were based on studies in animal models [93]. This is due in part to the limited efficiency of infection caused by the absence of CAR expression in many cancer (including gastrointestinal cancers, pancreatic cancer, ovarian cancer and hormone-refractory prostate cancer) and difficulties to obtain tumor-specific replication. Moreover, the immunogenicity of HAd serotypes and the abrogation of propagation due to immune response may be other reasons for the lower than expected clinical efficacy. Finally, complete tumor eradication needs an efficient dissemination of progeny virions in the tumor. This lateral spread of virus appeared limited by both cellular and tissue barriers between neighboring cancer cells [94]. In summary, improving the oncolytic potency of these viruses has been hampered by the inability to study host-vector interactions in immune-competent systems. Notably, HAd do not productively replicate in animal tissues, with some exceptions like Syrian hamsters [95]. Therefore, approaches such as immunomodulation, which could result in sustained replication and subsequently increased oncolysis, are of particular interest.
For these purposes and to treat “man’s best friend”, a CAV-2 conditionally replicating vector was created which effectively replicates in and causes oncolysis of canine osteosarcoma cells. In this vector, the E1A expression was driven by an osteocalcin promoter that restricts its expression and therefore CAV-2 propagation to such osteosarcoma cells [89]. Canine osteosarcoma is the most common primary bone tumor of dogs and is an interesting model of human counterpart. This virus effectively replicated and killed canine osteosarcoma cells in vitro. The next step was to test the efficiency and the toxicity of such vector in normal dogs before potential clinical trials in dogs [96]. Short-term physiologic indicators of stress and shock, as well as gross and histological changes in a variety of tissues were analyzed and no major signs of virus-associated toxicity were noted. Interestingly, short-term immunosuppression, allowing increased adenoviral gene transfer and reduced T-cell mediated and neutralizing antibody formation, did not increase CRAd toxicity. These results are particularly pertinent for translation into human Phase I trials.
6.3. Vaccine vectors
Due to their aptitude to induce potent innate and adoptive immune responses, Ad vectors have been and are being explored as vaccine carriers. Until recently, the most promising vaccines were based on E1 or E1/E3 deleted HAd5 vectors. In preclinical models, these vectors induced potent transgene product-specific T- and B-cell responses [97]. However, pre-existing neutralizing antibodies to HAd5 virus drastically reduce vector efficacy in animals [98] as well as in human [99,100]. Specific CD8+ T lymphocytes eliminated vector-transduced cells and thus shortening the duration of antigen expression.
Interestingly, the detection of high frequencies of HIV-1-specific CD4 and CD8 T cells in HIV-1-infected subjects with nonprogressive disease [101] and the demonstration that CD8 T cells are key players in vivo in the control of SIV replication [102] provided the rationale for developing T-cell vaccine strategies against HIV-1. Expressing conserved antigens of HIV-1 for induction of CD8+ T-cell responses, early phase clinical trials with HAd5 vaccine yielded sufficiently promising results in volunteers at low risk for HIV-1 acquisition to allow for a large phase 2b trial in human at high risk for HIV-1 infection. This STEP trial was prematurely stopped because an interim analysis showed lack of efficiency and even more a twofold increase in the incidence of HIV acquisition among vaccinated recipients with high HAd5 neutralizing antibodies titers compared with placebo recipients [103]. In a recent study, we suggested that this increase in acquisition of HIV infection could be due to the presence of high HAd5 NAbs titers favoring the formation of HAd5 immune complexes (IC) [82]. Compared to HAd5 vector alone, HAd5 IC induced “hyperactivation” and maturation of DCs and therefore a larger expansion of both HAd5-specific memory CD4 and CD8 T cells. Expansion prevent effective generation of the primary immune response against the vector-encoded HIV Ags through notably the killing of DCs and enlarge the pool of memory CD4 T cells that effectively support HIV replication and spreading, thus facilitating susceptibility to HIV infection.
To circumvent problems caused by the pre-existing vector immunity, an alternative approach is the use of vectors derived from nonhuman Ad types. Various nonhuman Ad are currently under investigation as gene expression and vaccine vectors such as CAV-2 but also bovine Ad serotype 3 (BAd3), chimpanzee Ad serotype 1, ovine Ad serotype 7 (OAd7), porcine Ad serotype 3 and 5 (PAd3, PAd5) and fowl Ad serotype 1, 8, 9 and 10 [20,22,104,105,106,107,108].
A very effective vaccine based on an avirulent CAV-2 vector is used worldwide for the routine vaccination against both canine infectious hepatitis (CAV-1) and canine infectious laryngotracheitis (CAV-2), with an excellent safety record. Interestingly, this vaccine is usually administered subcutaneously but it is also effective when administered mucosally [109]. More recently, a replication-competent recombinant CAV-2 vaccine was constructed by homologous recombination with canine distemper virus (CDV) antigen replacing the CAV-2 genomic E3 region [110]. Like in HAd5, the E3 region was non-essential for viral replication in tissue culture. This recombinant vaccine stimulated a protective response in both dogs and minks [110].
Recently, a similar strategy was used to develop a recombinant vaccine in which the glycoprotein gene of rabies (CVS) was cloned and expressed. The effectiveness in protecting against CVS was demonstrated in dogs, cats and pigs [87,88,111].
Importantly, like most other adenoviruses, CAV-2 is also prevalent in their natural hosts and neutralizing antibodies against CAV-2 can have a negative effect on vaccine efficiency. For instance, intranasal vaccination with CAV-2 vector expressing CDV hemaglutinin induced significant levels of CDV-specific immunity in seronegative puppies but a poor immune response in the puppies pre-exposed to CAV-2 [110]. This difference was attributed to the presence of an anti-CAV-2 mucosal immunity in these pre-exposed animals. As mentioned above, neutralizing antibodies against CAV-2 are not prevalent in humans and are not cross-neutralized by HAd neutralizing antibodies. Therefore, CAV-2 vectors may be an alternative for human as well as veterinary use.
Conclusion
Significant progress has been made in the understanding of the biology of CAV-2 and in particular CAV-2 vectors. The paucity of pre-existing immunity in human, its preferential interaction with CAR and therefore a neuronal tropism in the CNS, and its axonal transport are characteristics favorable to address fundamental neurobiological questions and to develop potential treatment of neurodegenerative disorders. Currently, CAV-2 vectors have found a niche in gene transfer to the CNS, which limits it to labs with expertise in stereotactic brain surgery and interest in gene transfer. In addition to the native tropism of CAV-2 vectors, mutant vectors harboring a point mutation that abolishes CAR interaction, would be an ideal platform for targeted gene delivery for numerous tissues.
Acknowledgments
We thank the members of the Kremer lab for helpful discussions and illustrations. We thank Natalia Paz Mena for Figure 1. The work in the Kremer lab is supported by Agence Nationale pour la recherche, the European Community’s Seventh Framework programme (FP7/2007-2013) under grant agreement n° 222992 – BrainCAV, the Fondation de France, the Region Languedoc Roussillon, Association Francaise contre les Myopathies, Vaincre les Maladies Lysosomales, Association pour la Recherche sur la Sclerose Lateral Amytrophique et autres maladies du motoneurone, the Motor Neuron Disease Association (MNDA), European Research Projects on Rare Diseases, Foundation Beatriu de Pinós, Cystinosis Research Foundation, Cystinosis Research Network.
References and Notes
- Davison, A.J.; Benko, M.; Harrach, B. Genetic content and evolution of adenoviruses. J. Gen. Virol. 2003, 84, 2895–2908. [Google Scholar] [CrossRef]
- Hierholzer, J.C. Adenoviruses in the immunocompromised host. Clin. Microbiol. Rev. 1992, 5, 262–274. [Google Scholar] [CrossRef] [PubMed]
- Regagnon, C.; Souweine, B.; Archimbaud, C.; Duperron, F.; Thouvenot, D.; Peigue-Lafeuille, H. [A fatal case of adenovirus type 3 pneumonia in an immunocompetent adult]. Med. Mal. Infect. 2004, 34, 102–104. [Google Scholar] [CrossRef] [PubMed]
- Wadell, G.; Allard, A.; Johansson, M.; Svensson, L.; Uhnoo, I. Enteric adenoviruses. Ciba Found Symp. 1987, 128, 63–91. [Google Scholar] [PubMed]
- Chakrabarti, S.; Collingham, K.E.; Fegan, C.D.; Milligan, D.W. Fulminant adenovirus hepatitis following unrelated bone marrow transplantation: Failure of intravenous ribavirin therapy. Bone Marrow Transplant 1999, 23, 1209–1211. [Google Scholar] [CrossRef] [PubMed]
- Chakrabarti, S.; Mautner, V.; Osman, H.; Collingham, K.E.; Fegan, C.D.; Klapper, P.E.; Moss, P.A.; Milligan, D.W. Adenovirus infections following allogeneic stem cell transplantation: Incidence and outcome in relation to graft manipulation, immunosuppression, and immune recovery. Blood 2002, 100, 1619–1627. [Google Scholar] [CrossRef]
- D’Ambrosio, E.; Del Grosso, N.; Chicca, A.; Midulla, M. Neutralizing antibodies against 33 human adenoviruses in normal children in Rome. J. Hyg. (Lond) 1982, 89, 155–161. [Google Scholar] [CrossRef]
- Piedra, P.A.; Poveda, G.A.; Ramsey, B.; McCoy, K.; Hiatt, P.W. Incidence and prevalence of neutralizing antibodies to the common adenoviruses in children with cystic fibrosis: Implication for gene therapy with adenovirus vectors. Pediatrics 1998, 101, 1013–1019. [Google Scholar] [CrossRef]
- Wadell, G. Sensitization and neutralization of adenovirus by specific sera against capsid subunits. J. Immunol. 1972, 108, 622–632. [Google Scholar] [CrossRef]
- Perreau, M.; Kremer, E.J. The conundrum between immunological memory to adenovirus and their use as vectors in clinical gene therapy. Mol. Biotechnol. 2006, 34, 247–256. [Google Scholar] [CrossRef]
- Flomenberg, P.; Piaskowski, V.; Truitt, R.L.; Casper, J.T. Characterization of human proliferative T cell responses to adenovirus. J. Infect. Dis. 1995, 171, 1090–1096. [Google Scholar] [CrossRef] [PubMed]
- Olive, M.; Eisenlohr, L.C.; Flomenberg, P. Quantitative analysis of adenovirus-specific CD4+ T-cell responses from healthy adults. Viral Immunol. 2001, 14, 403–413. [Google Scholar] [CrossRef] [PubMed]
- Klonjkowski, B.; Gilardi-Hebenstreit, P.; Hadchouel, J.; Randrianarison, V.; Boutin, S.; Yeh, P.; Perricaudet, M.; Kremer, E.J. A recombinant E1-deleted canine adenoviral vector capable of transduction and expression of a transgene in human-derived cells and in vivo. Hum. Gene Ther. 1997, 8, 2103–2115. [Google Scholar] [CrossRef] [PubMed]
- Kremer, E.J. CAR chasing: Canine adenovirus vectors-all bite and no bark? J. Gene Med. 2004, 6 (Suppl. 1), S139–151. [Google Scholar] [CrossRef]
- Kremer, E.J.; Boutin, S.; Chillon, M.; Danos, O. Canine adenovirus vectors: An alternative for adenovirus-mediated gene transfer. J. Virol. 2000, 74, 505–512. [Google Scholar] [CrossRef]
- Soudais, C.; Laplace-Builhe, C.; Kissa, K.; Kremer, E.J. Preferential transduction of neurons by canine adenovirus vectors and their efficient retrograde transport in vivo. Faseb J. 2001, 15, 2283–2285. [Google Scholar] [CrossRef]
- Soudais, C.; Skander, N.; Kremer, E.J. Long-term in vivo transduction of neurons throughout the rat CNS using novel helper-dependent CAV-2 vectors. Faseb J. 2004, 18, 391–393. [Google Scholar] [CrossRef]
- Both, G.W. Ovine atadenovirus: A review of its biology, biosafety profile and application as a gene delivery vector. Immunol. Cell. Biol. 2004, 82, 189–195. [Google Scholar] [CrossRef]
- Reddy, P.S.; Idamakanti, N.; Chen, Y.; Whale, T.; Babiuk, L.A.; Mehtali, M.; Tikoo, S.K. Replication-defective bovine adenovirus type 3 as an expression vector. J. Virol. 1999, 73, 9137–9144. [Google Scholar] [CrossRef]
- Reddy, P.S.; Idamakanti, N.; Hyun, B.H.; Tikoo, S.K.; Babiuk, L.A. Development of porcine adenovirus-3 as an expression vector. J. Gen. Virol. 1999, 80 (Pt 3), 563–570. [Google Scholar] [CrossRef]
- Farina, S.F.; Gao, G.P.; Xiang, Z.Q.; Rux, J.J.; Burnett, R.M.; Alvira, M.R.; Marsh, J.; Ertl, H.C.; Wilson, J.M. Replication-defective vector based on a chimpanzee adenovirus. J. Virol. 2001, 75, 11603–11613. [Google Scholar] [CrossRef]
- Xiang, Z.; Gao, G.; Reyes-Sandoval, A.; Cohen, C.J.; Li, Y.; Bergelson, J.M.; Wilson, J.M.; Ertl, H.C. Novel, chimpanzee serotype 68-based adenoviral vaccine carrier for induction of antibodies to a transgene product. J. Virol. 2002, 76, 2667–2675. [Google Scholar] [CrossRef] [PubMed]
- Paillard, F. Advantages of non-human adenoviruses versus human adenoviruses. Hum. Gene Ther. 1997, 8, 2007–2009. [Google Scholar] [CrossRef] [PubMed]
- Chartier, C.; Degryse, E.; Gantzer, M.; Dieterle, A.; Pavirani, A.; Mehtali, M. Efficient generation of recombinant adenovirus vectors by homologous recombination in Escherichia coli. J. Virol. 1996, 70, 4805–4810. [Google Scholar] [CrossRef]
- Soudais, C.; Boutin, S.; Kremer, E.J. Characterization of cis-acting sequences involved in canine adenovirus packaging. Mol. Ther. 2001, 3, 631–640. [Google Scholar] [CrossRef]
- Parks, R.J.; Chen, L.; Anton, M.; Sankar, U.; Rudnicki, M.A.; Graham, F.L. A helper-dependent adenovirus vector system: Removal of helper virus by Cre-mediated excision of the viral packaging signal. Proc. Natl. Acad. Sci. U. S. A. 1996, 93, 13565–13570. [Google Scholar] [CrossRef] [PubMed]
- Kochanek, S.; Clemens, P.R.; Mitani, K.; Chen, H.H.; Chan, S.; Caskey, C.T. A new adenoviral vector: Replacement of all viral coding sequences with 28 kb of DNA independently expressing both full-length dystrophin and beta-galactosidase. Proc. Natl. Acad. Sci. U. S. A. 1996, 93, 5731–5736. [Google Scholar] [CrossRef]
- Amalfitano, A.; Parks, R.J. Separating fact from fiction: Assessing the potential of modified adenovirus vectors for use in human gene therapy. Curr. Gene Ther. 2002, 2, 111–133. [Google Scholar] [CrossRef]
- Alba, R.; Bosch, A.; Chillon, M. Gutless adenovirus: Last-generation adenovirus for gene therapy. Gene Ther. 2005, 12 (Suppl. 1), S18-27. [Google Scholar] [CrossRef]
- Cornwell, H.J.; Koptopoulos, G.; Thompson, H.; McCandlish, I.A.; Wright, N.G. Immunity to canine adenovirus respiratory disease: A comparison of attenuated CAV-1 and CAV-2 vaccines. Vet. Rec. 1982, 110, 27–32. [Google Scholar] [CrossRef] [PubMed]
- Philippa, J.D.; Leighton, F.A.; Daoust, P.Y.; Nielsen, O.; Pagliarulo, M.; Schwantje, H.; Shury, T.; Van Herwijnen, R.; Martina, B.E.; Kuiken, T.; Van de Bildt, M.W.; Osterhaus, A.D. Antibodies to selected pathogens in free-ranging terrestrial carnivores and marine mammals in Canada. Vet. Rec. 2004, 155, 135–140. [Google Scholar] [CrossRef] [PubMed]
- Salinas, S.; Schiavo, G.; Kremer, E.J. A hitchhiker’s guide to the nervous system: The complex journey of viruses and toxins. Nat. Rev. Microbiol. 8, 645–655. [CrossRef] [PubMed]
- Smith, G.A.; Pomeranz, L.; Gross, S.P.; Enquist, L.W. Local modulation of plus-end transport targets herpesvirus entry and egress in sensory axons. Proc. Natl. Acad. Sci. U. S. A. 2004, 101, 16034–16039. [Google Scholar] [CrossRef]
- Mueller, S.; Wimmer, E.; Cello, J. Poliovirus and poliomyelitis: A tale of guts, brains, and an accidental event. Virus Res. 2005, 111, 175–193. [Google Scholar] [CrossRef] [PubMed]
- Salinas, S.; Bilsland, L.G.; Henaff, D.; Weston, A.E.; Keriel, A.; Schiavo, G.; Kremer, E.J. CAR-associated vesicular transport of an adenovirus in motor neuron axons. PLoS Pathog. 2009, 5, e1000442. [Google Scholar] [CrossRef] [PubMed]
- Henaff, D.; Salinas, S. An endocytic CARriage tale: Adenoviruses internalization and trafficking in neurons. Virulence 2010, 1, 188–191. [Google Scholar] [CrossRef] [PubMed]
- Leopold, P.L.; Crystal, R.G. Intracellular trafficking of adenovirus: Many means to many ends. Adv. Drug Deliv. Rev. 2007, 59, 810–821. [Google Scholar] [CrossRef]
- Meier, O.; Greber, U.F. Adenovirus endocytosis. J. Gene Med. 2003, 5, 451–462. [Google Scholar] [CrossRef]
- Bremner, K.H.; Scherer, J.; Yi, J.; Vershinin, M.; Gross, S.P.; Vallee, R.B. Adenovirus transport via direct interaction of cytoplasmic dynein with the viral capsid hexon subunit. Cell Host Microbe 2009, 6, 523–535. [Google Scholar] [CrossRef]
- Soudais, C.; Boutin, S.; Hong, S.S.; Chillon, M.; Danos, O.; Bergelson, J.M.; Boulanger, P.; Kremer, E.J. Canine adenovirus type 2 attachment and internalization: Coxsackievirus-adenovirus receptor, alternative receptors, and an RGD-independent pathway. J. Virol. 2000, 74, 10639–10649. [Google Scholar] [CrossRef]
- Freimuth, P.; Philipson, L.; Carson, S.D. The coxsackievirus and adenovirus receptor. Curr. Top. Microbiol. Immunol. 2008, 323, 67–87. [Google Scholar] [PubMed]
- Honda, T.; Saitoh, H.; Masuko, M.; Katagiri-Abe, T.; Tominaga, K.; Kozakai, I.; Kobayashi, K.; Kumanishi, T.; Watanabe, Y.G.; Odani, S.; Kuwano, R. The coxsackievirus-adenovirus receptor protein as a cell adhesion molecule in the developing mouse brain. Brain Res. Mol. Brain Res. 2000, 77, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Lim, B.K.; Xiong, D.; Dorner, A.; Youn, T.J.; Yung, A.; Liu, T.I.; Gu, Y.; Dalton, N.D.; Wright, A.T.; Evans, S.M.; Chen, J.; Peterson, K.L.; McCulloch, A.D.; Yajima, T.; Knowlton, K.U. Coxsackievirus and adenovirus receptor (CAR) mediates atrioventricular-node function and connexin 45 localization in the murine heart. J. Clin. Invest. 2008, 118, 2758–2770. [Google Scholar] [CrossRef] [PubMed]
- Keriel, A.; Rene, C.; Galer, C.; Zabner, J.; Kremer, E.J. Canine adenovirus vectors for lung-directed gene transfer: Efficacy, immune response, and duration of transgene expression using helper-dependent vectors. J. Virol. 2006, 80, 1487–1496. [Google Scholar] [CrossRef]
- Bergelson, J.M.; Cunningham, J.A.; Droguett, G.; Kurt-Jones, E.A.; Krithivas, A.; Hong, J.S.; Horwitz, M.S.; Crowell, R.L.; Finberg, R.W. Isolation of a common receptor for Coxsackie B viruses and adenoviruses 2 and 5. Science 1997, 275, 1320–1323. [Google Scholar] [CrossRef]
- Roelvink, P.W.; Lizonova, A.; Lee, J.G.; Li, Y.; Bergelson, J.M.; Finberg, R.W.; Brough, D.E.; Kovesdi, I.; Wickham, T.J. The coxsackievirus-adenovirus receptor protein can function as a cellular attachment protein for adenovirus serotypes from subgroups A, C, D, E, and F. J. Virol. 1998, 72, 7909–7915. [Google Scholar] [CrossRef]
- Carson, S.D. Receptor for the group B coxsackieviruses and adenoviruses: CAR. Rev. Med. Virol. 2001, 11, 219–226. [Google Scholar] [CrossRef]
- Wang, K.; Huang, S.; Kapoor-Munshi, A.; Nemerow, G. Adenovirus internalization and infection require dynamin. J. Virol. 1998, 72, 3455–3458. [Google Scholar] [CrossRef]
- Wickham, T.J.; Mathias, P.; Cheresh, D.A.; Nemerow, G.R. Integrins alpha v beta 3 and alpha v beta 5 promote adenovirus internalization but not virus attachment. Cell 1993, 73, 309–319. [Google Scholar] [CrossRef]
- Chiu, C.Y.; Mathias, P.; Nemerow, G.R.; Stewart, P.L. Structure of adenovirus complexed with its internalization receptor, alphavbeta5 integrin. J. Virol. 1999, 73, 6759–6768. [Google Scholar] [CrossRef]
- Bewley, M.C.; Springer, K.; Zhang, Y.B.; Freimuth, P.; Flanagan, J.M. Structural analysis of the mechanism of adenovirus binding to its human cellular receptor, CAR. Science 1999, 286, 1579–1583. [Google Scholar] [CrossRef] [PubMed]
- Kirby, I.; Davison, E.; Beavil, A.J.; Soh, C.P.; Wickham, T.J.; Roelvink, P.W.; Kovesdi, I.; Sutton, B.J.; Santis, G. Identification of contact residues and definition of the CAR-binding site of adenovirus type 5 fiber protein. J. Virol. 2000, 74, 2804–2813. [Google Scholar] [CrossRef] [PubMed]
- Roelvink, P.W.; Mi Lee, G.; Einfeld, D.A.; Kovesdi, I.; Wickham, T.J. Identification of a conserved receptor-binding site on the fiber proteins of CAR-recognizing adenoviridae. Science 1999, 286, 1568–1571. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.S.; Karayan, L.; Tournier, J.; Curiel, D.T.; Boulanger, P.A. Adenovirus type 5 fiber knob binds to MHC class I alpha2 domain at the surface of human epithelial and B lymphoblastoid cells. Embo J. 1997, 16, 2294–2306. [Google Scholar] [CrossRef]
- Fabry, C.M.; Rosa-Calatrava, M.; Conway, J.F.; Zubieta, C.; Cusack, S.; Ruigrok, R.W.; Schoehn, G. A quasi-atomic model of human adenovirus type 5 capsid. Embo J. 2005, 24, 1645–1654. [Google Scholar] [CrossRef]
- Baker, A.H.; McVey, J.H.; Waddington, S.N.; Di Paolo, N.C.; Shayakhmetov, D.M. The influence of blood on in vivo adenovirus bio-distribution and transduction. Mol. Ther. 2007, 15, 1410–1416. [Google Scholar] [CrossRef]
- Johansson, C.; Jonsson, M.; Marttila, M.; Persson, D.; Fan, X.L.; Skog, J.; Frangsmyr, L.; Wadell, G.; Arnberg, N. Adenoviruses use lactoferrin as a bridge for CAR-independent binding to and infection of epithelial cells. J. Virol. 2007, 81, 954–963. [Google Scholar] [CrossRef]
- Gaggar, A.; Shayakhmetov, D.M.; Lieber, A. CD46 is a cellular receptor for group B adenoviruses. Nat. Med. 2003, 9, 1408–1412. [Google Scholar] [CrossRef]
- Marttila, M.; Persson, D.; Gustafsson, D.; Liszewski, M.K.; Atkinson, J.P.; Wadell, G.; Arnberg, N. CD46 is a cellular receptor for all species B adenoviruses except types 3 and 7. J. Virol. 2005, 79, 14429–14436. [Google Scholar] [CrossRef]
- Kremer, E.J. Mutagenesis of hexon "FX" hepatic tropism. Blood 2009, 114, 929–930. [Google Scholar] [CrossRef]
- Schoehn, G.; El Bakkouri, M.; Fabry, C.M.; Billet, O.; Estrozi, L.F.; Le, L.; Curiel, D.T.; Kajava, A.V.; Ruigrok, R.W.; Kremer, E.J. Three-dimensional structure of canine adenovirus serotype 2 capsid. J. Virol. 2008, 82, 3192–3203. [Google Scholar] [CrossRef] [PubMed]
- Perreau, M.; Guerin, M.C.; Drouet, C.; Kremer, E.J. Interactions between human plasma components and a xenogenic adenovirus vector: Reduced immunogenicity during gene transfer. Mol. Ther. 2007, 15, 1998–2007. [Google Scholar] [CrossRef] [PubMed]
- Schmitz, H.; Wigand, R.; Heinrich, W. Worldwide epidemiology of human adenovirus infections. Am. J. Epidemiol. 1983, 117, 455–466. [Google Scholar] [CrossRef] [PubMed]
- Sumida, S.M.; Truitt, D.M.; Kishko, M.G.; Arthur, J.C.; Jackson, S.S.; Gorgone, D.A.; Lifton, M.A.; Koudstaal, W.; Pau, M.G.; Kostense, S.; Havenga, M.J.; Goudsmit, J.; Letvin, N.L.; Barouch, D.H. Neutralizing antibodies and CD8+ T lymphocytes both contribute to immunity to adenovirus serotype 5 vaccine vectors. J. Virol. 2004, 78, 2666–2673. [Google Scholar] [CrossRef] [PubMed]
- Gahery-Segard, H.; Juillard, V.; Gaston, J.; Lengagne, R.; Pavirani, A.; Boulanger, P.; Guillet, J.G. Humoral immune response to the capsid components of recombinant adenoviruses: Routes of immunization modulate virus-induced Ig subclass shifts. Eur. J. Immunol. 1997, 27, 653–659. [Google Scholar] [CrossRef]
- Bjorklund, A.; Kirik, D.; Rosenblad, C.; Georgievska, B.; Lundberg, C.; Mandel, R.J. Towards a neuroprotective gene therapy for Parkinson’s disease: Use of adenovirus, AAV and lentivirus vectors for gene transfer of GDNF to the nigrostriatal system in the rat Parkinson model. Brain Res. 2000, 886, 82–98. [Google Scholar] [CrossRef]
- Echavarria, M. Adenoviruses in immunocompromised hosts. Clin. Microbiol. Rev. 2008, 21, 704–715. [Google Scholar] [CrossRef]
- Bangari, D.S.; Mittal, S.K. Porcine adenoviral vectors evade preexisting humoral immunity to adenoviruses and efficiently infect both human and murine cells in culture. Virus Res. 2004, 105, 127–136. [Google Scholar] [CrossRef]
- Bangari, D.S.; Shukla, S.; Mittal, S.K. Comparative transduction efficiencies of human and nonhuman adenoviral vectors in human, murine, bovine, and porcine cells in culture. Biochem. Biophys. Res. Commun. 2005, 327, 960–966. [Google Scholar] [CrossRef]
- Sharma, A.; Tandon, M.; Ahi, Y.S.; Bangari, D.S.; Vemulapalli, R.; Mittal, S.K. Evaluation of cross-reactive cell-mediated immune responses among human, bovine and porcine adenoviruses. Gene Ther. 17, 634–642. [CrossRef]
- Hofmann, C.; Loser, P.; Cichon, G.; Arnold, W.; Both, G.W.; Strauss, M. Ovine adenovirus vectors overcome preexisting humoral immunity against human adenoviruses in vivo. J. Virol. 1999, 73, 6930–6936. [Google Scholar] [CrossRef] [PubMed]
- Pichla-Gollon, S.L.; Lin, S.W.; Hensley, S.E.; Lasaro, M.O.; Herkenhoff-Haut, L.; Drinker, M.; Tatsis, N.; Gao, G.P.; Wilson, J.M.; Ertl, H.C.; Bergelson, J.M. Effect of preexisting immunity on an adenovirus vaccine vector: In vitro neutralization assays fail to predict inhibition by antiviral antibody in vivo. J. Virol. 2009, 83, 5567–5573. [Google Scholar] [CrossRef]
- Mastrangeli, A.; Harvey, B.G.; Yao, J.; Wolff, G.; Kovesdi, I.; Crystal, R.G.; Falck-Pedersen, E. "Sero-switch" adenovirus-mediated in vivo gene transfer: Circumvention of anti-adenovirus humoral immune defenses against repeat adenovirus vector administration by changing the adenovirus serotype. Hum. Gene Ther. 1996, 7, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Ho, S.; Clipstone, N.; Timmermann, L.; Northrop, J.; Graef, I.; Fiorentino, D.; Nourse, J.; Crabtree, G.R. The mechanism of action of cyclosporin A and FK506. Clin. Immunol. Immunopathol. 1996, 80, S40–S45. [Google Scholar] [CrossRef] [PubMed]
- Perreau, M.; Kremer, E.J. Frequency, proliferation, and activation of human memory T cells induced by a nonhuman adenovirus. J. Virol. 2005, 79, 14595–14605. [Google Scholar] [CrossRef]
- Perreau, M.; Mennechet, F.; Serratrice, N.; Glasgow, J.N.; Curiel, D.T.; Wodrich, H.; Kremer, E.J. Contrasting effects of human, canine, and hybrid adenovirus vectors on the phenotypical and functional maturation of human dendritic cells: Implications for clinical efficacy. J. Virol. 2007, 81, 3272–3284. [Google Scholar] [CrossRef]
- Foti, M.; Granucci, F.; Ricciardi-Castagnoli, P. A central role for tissue-resident dendritic cells in innate responses. Trends Immunol. 2004, 25, 650–654. [Google Scholar] [CrossRef]
- Le Bon, A.; Tough, D.F. Links between innate and adaptive immunity via type I interferon. Curr. Opin. Immunol. 2002, 14, 432–436. [Google Scholar] [CrossRef]
- Colonna, M.; Pulendran, B.; Iwasaki, A. Dendritic cells at the host-pathogen interface. Nat. Immunol. 2006, 7, 117–120. [Google Scholar] [CrossRef]
- Underhill, D.M.; Ozinsky, A.; Hajjar, A.M.; Stevens, A.; Wilson, C.B.; Bassetti, M.; Aderem, A. The Toll-like receptor 2 is recruited to macrophage phagosomes and discriminates between pathogens. Nature 1999, 401, 811–815. [Google Scholar] [CrossRef]
- Banchereau, J.; Briere, F.; Caux, C.; Davoust, J.; Lebecque, S.; Liu, Y.J.; Pulendran, B.; Palucka, K. Immunobiology of dendritic cells. Annu. Rev. Immunol. 2000, 18, 767–811. [Google Scholar] [CrossRef] [PubMed]
- Perreau, M.; Pantaleo, G.; Kremer, E.J. Activation of a dendritic cell-T cell axis by Ad5 immune complexes creates an improved environment for replication of HIV in T cells. J. Exp. Med. 2008, 205, 2717–2725. [Google Scholar] [CrossRef] [PubMed]
- Hnasko, T.S.; Perez, F.A.; Scouras, A.D.; Stoll, E.A.; Gale, S.D.; Luquet, S.; Phillips, P.E.; Kremer, E.J.; Palmiter, R.D. Cre recombinase-mediated restoration of nigrostriatal dopamine in dopamine-deficient mice reverses hypophagia and bradykinesia. Proc. Natl. Acad. Sci. U. S. A. 2006, 103, 8858–8863. [Google Scholar] [CrossRef] [PubMed]
- Morante-Oria, J.; Carleton, A.; Ortino, B.; Kremer, E.J.; Fairen, A.; Lledo, P.M. Subpallial origin of a population of projecting pioneer neurons during corticogenesis. Proc. Natl. Acad. Sci. U. S. A. 2003, 100, 12468–12473. [Google Scholar] [CrossRef]
- Kissa, K.; Mordelet, E.; Soudais, C.; Kremer, E.J.; Demeneix, B.A.; Brulet, P.; Coen, L. In vivo neuronal tracing with GFP-TTC gene delivery. Mol. Cell. Neurosci. 2002, 20, 627–637. [Google Scholar] [CrossRef]
- Lau, A.A.; Hopwood, J.J.; Kremer, E.J.; Hemsley, K.M. SGSH gene transfer in mucopolysaccharidosis type IIIA mice using canine adenovirus vectors. Mol. Genet. Metab. 2010, 100, 168–175. [Google Scholar] [CrossRef]
- Hu, R.; Zhang, S.; Fooks, A.R.; Yuan, H.; Liu, Y.; Li, H.; Tu, C.; Xia, X.; Xiao, Y. Prevention of rabies virus infection in dogs by a recombinant canine adenovirus type-2 encoding the rabies virus glycoprotein. Microbes Infect. 2006, 8, 1090–1097. [Google Scholar] [CrossRef]
- Hu, R.L.; Liu, Y.; Zhang, S.F.; Zhang, F.; Fooks, A.R. Experimental immunization of cats with a recombinant rabies-canine adenovirus vaccine elicits a long-lasting neutralizing antibody response against rabies. Vaccine 2007, 25, 5301–5307. [Google Scholar] [CrossRef]
- Hemminki, A.; Kanerva, A.; Kremer, E.J.; Bauerschmitz, G.J.; Smith, B.F.; Liu, B.; Wang, M.; Desmond, R.A.; Keriel, A.; Barnett, B.; Baker, H.J.; Siegal, G.P.; Curiel, D.T. A canine conditionally replicating adenovirus for evaluating oncolytic virotherapy in a syngeneic animal model. Mol. Ther. 2003, 7, 163–173. [Google Scholar] [CrossRef]
- Hemsley, K.M.; Luck, A.J.; Crawley, A.C.; Hassiotis, S.; Beard, H.; King, B.; Rozek, T.; Rozaklis, T.; Fuller, M.; Hopwood, J.J. Examination of intravenous and intra-CSF protein delivery for treatment of neurological disease. Eur. J. Neurosci. 2009, 29, 1197–1214. [Google Scholar] [CrossRef]
- Hemsley, K.M.; Beard, H.; King, B.M.; Hopwood, J.J. Effect of high dose, repeated intra-CSF injection of sulphamidase on neuropathology in MPS IIIA mice. Genes Brain Behav. 2008, 7, 740–753. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, M.; Curiel, D.T. Current issues and future directions of oncolytic adenoviruses. Mol. Ther. 2010, 18, 243–250. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, M.; Curiel, D.T. Cancer gene therapy. Technol. Cancer Res. Treat. 2005, 4, 315–330. [Google Scholar] [CrossRef] [PubMed]
- Dobbelstein, M. Replicating adenoviruses in cancer therapy. Curr. Top. Microbiol. Immunol. 2004, 273, 291–334. [Google Scholar]
- Thomas, M.A.; Spencer, J.F.; La Regina, M.C.; Dhar, D.; Tollefson, A.E.; Toth, K.; Wold, W.S. Syrian hamster as a permissive immunocompetent animal model for the study of oncolytic adenovirus vectors. Cancer Res. 2006, 66, 1270–1276. [Google Scholar] [CrossRef]
- Smith, B.F.; Curiel, D.T.; Ternovoi, V.V.; Borovjagin, A.V.; Baker, H.J.; Cox, N.; Siegal, G.P. Administration of a conditionally replicative oncolytic canine adenovirus in normal dogs. Cancer Biother. Radiopharm. 2006, 21, 601–606. [Google Scholar] [CrossRef]
- Xiang, Z.Q.; Yang, Y.; Wilson, J.M.; Ertl, H.C. A replication-defective human adenovirus recombinant serves as a highly efficacious vaccine carrier. Virology 1996, 219, 220–227. [Google Scholar] [CrossRef]
- Yang, Y.; Li, Q.; Ertl, H.C.; Wilson, J.M. Cellular and humoral immune responses to viral antigens create barriers to lung-directed gene therapy with recombinant adenoviruses. J. Virol. 1995, 69, 2004–2015. [Google Scholar] [CrossRef]
- Shiver, J.W.; Emini, E.A. Recent advances in the development of HIV-1 vaccines using replication-incompetent adenovirus vectors. Annu. Rev. Med. 2004, 55, 355–372. [Google Scholar] [CrossRef]
- Lasaro, M.O.; Ertl, H.C. New insights on adenovirus as vaccine vectors. Mol. Ther. 2009, 17, 1333–1339. [Google Scholar] [CrossRef]
- Pantaleo, G.; Koup, R.A. Correlates of immune protection in HIV-1 infection: What we know, what we don’t know, what we should know. Nat. Med. 2004, 10, 806–810. [Google Scholar] [CrossRef] [PubMed]
- Schmitz, J.E.; Kuroda, M.J.; Santra, S.; Sasseville, V.G.; Simon, M.A.; Lifton, M.A.; Racz, P.; Tenner-Racz, K.; Dalesandro, M.; Scallon, B.J.; Ghrayeb, J.; Forman, M.A.; Montefiori, D.C.; Rieber, E.P.; Letvin, N.L.; Reimann, K.A. Control of viremia in simian immunodeficiency virus infection by CD8+ lymphocytes. Science 1999, 283, 857–860. [Google Scholar] [CrossRef]
- Buchbinder, S.P.; Mehrotra, D.V.; Duerr, A.; Fitzgerald, D.W.; Mogg, R.; Li, D.; Gilbert, P.B.; Lama, J.R.; Marmor, M.; Del Rio, C.; McElrath, M.J.; Casimiro, D.R.; Gottesdiener, K.M.; Chodakewitz, J.A.; Corey, L.; Robertson, M.N. Efficacy assessment of a cell-mediated immunity HIV-1 vaccine (the Step Study): A double-blind, randomised, placebo-controlled, test-of-concept trial. Lancet 2008, 372, 1881–1893. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Pandey, A.; Jayashankar, L.; Mittal, S.K. Bovine adenoviral vector-based H5N1 influenza vaccine overcomes exceptionally high levels of pre-existing immunity against human adenovirus. Mol. Ther. 2008, 16, 965–971. [Google Scholar] [CrossRef] [PubMed]
- Wuest, T.; Both, G.W.; Prince, A.M.; Hofmann, C.; Loser, P. Recombinant ovine atadenovirus induces a strong and sustained T cell response against the hepatitis C virus NS3 antigen in mice. Vaccine 2004, 22, 2717–2721. [Google Scholar] [CrossRef]
- Hammond, J.M.; McCoy, R.J.; Jansen, E.S.; Morrissy, C.J.; Hodgson, A.L.; Johnson, M.A. Vaccination with a single dose of a recombinant porcine adenovirus expressing the classical swine fever virus gp55 (E2) gene protects pigs against classical swine fever. Vaccine 2000, 18, 1040–1050. [Google Scholar] [CrossRef]
- Francois, A.; Chevalier, C.; Delmas, B.; Eterradossi, N.; Toquin, D.; Rivallan, G.; Langlois, P. Avian adenovirus CELO recombinants expressing VP2 of infectious bursal disease virus induce protection against bursal disease in chickens. Vaccine 2004, 22, 2351–2360. [Google Scholar] [CrossRef]
- Bridgeman, A.; Roshorm, Y.; Lockett, L.J.; Xu, Z.Z.; Hopkins, R.; Shaw, J.; Both, G.W.; Hanke, T. Ovine atadenovirus, a novel and highly immunogenic vector in prime-boost studies of a candidate HIV-1 vaccine. Vaccine 2009, 28, 474–483. [Google Scholar] [CrossRef]
- Baer, G.M.; Brooks, R.C.; Foggin, C.M. Oral vaccination of dogs fed canine adenovirus in baits. Am. J. Vet. Res. 1989, 50, 836–837. [Google Scholar]
- Fischer, L.; Tronel, J.P.; Pardo-David, C.; Tanner, P.; Colombet, G.; Minke, J.; Audonnet, J.C. Vaccination of puppies born to immune dams with a canine adenovirus-based vaccine protects against a canine distemper virus challenge. Vaccine 2002, 20, 3485–3497. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, S.; Ma, G.; Zhang, F.; Hu, R. Efficacy and safety of a live canine adenovirus-vectored rabies virus vaccine in swine. Vaccine 2008, 26, 5368–5372. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Stereotactic injection of a CAV-2 vector expressing GFP in the rat striatum. (A-B) Schematic inserts show respectively midsagittal and frontal cross sections of the rat brain and the location of the injection site (striatum) relative to the location of the transduced cells (substantia nigra (denoted SN)). (C) Immunolabeling for tyrosine hydroxylase (TH) expressed in the substantia nigra (red). (D) Retrograde axonal transport and expression in the vector-transduced neurons (green) in the substantia nigra. (E) Colocalization of GFP with TH.
Figure 1.
Stereotactic injection of a CAV-2 vector expressing GFP in the rat striatum. (A-B) Schematic inserts show respectively midsagittal and frontal cross sections of the rat brain and the location of the injection site (striatum) relative to the location of the transduced cells (substantia nigra (denoted SN)). (C) Immunolabeling for tyrosine hydroxylase (TH) expressed in the substantia nigra (red). (D) Retrograde axonal transport and expression in the vector-transduced neurons (green) in the substantia nigra. (E) Colocalization of GFP with TH.

Figure 3.
An overview of molecules involved in adenovirus binding. Adenoviruses can attach to CAR, sialic acid, CD46, CD86, CD80, MHC-I and heparin sulfate proteoglycan (HSPG) depending on the serotype. Some HAds also use coagulation factors and lactoferrin as bridges to attach to cells. To the best of our knowledge, CAR is the only receptor for CAV-2. (Image modified from Kremer, Blood [60]).
Figure 3.
An overview of molecules involved in adenovirus binding. Adenoviruses can attach to CAR, sialic acid, CD46, CD86, CD80, MHC-I and heparin sulfate proteoglycan (HSPG) depending on the serotype. Some HAds also use coagulation factors and lactoferrin as bridges to attach to cells. To the best of our knowledge, CAR is the only receptor for CAV-2. (Image modified from Kremer, Blood [60]).

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2010 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/3.0/).
Share and Cite
MDPI and ACS Style
Bru, T.; Salinas, S.; Kremer, E.J. An Update on Canine Adenovirus Type 2 and Its Vectors. Viruses 2010, 2, 2134-2153. https://doi.org/10.3390/v2092134
AMA Style
Bru T, Salinas S, Kremer EJ. An Update on Canine Adenovirus Type 2 and Its Vectors. Viruses. 2010; 2(9):2134-2153. https://doi.org/10.3390/v2092134
Chicago/Turabian StyleBru, Thierry, Sara Salinas, and Eric J. Kremer. 2010. "An Update on Canine Adenovirus Type 2 and Its Vectors" Viruses 2, no. 9: 2134-2153. https://doi.org/10.3390/v2092134