
Scorpion Toxin, BmP01, Induces Pain by Targeting TRPV1 Channel

Abstract
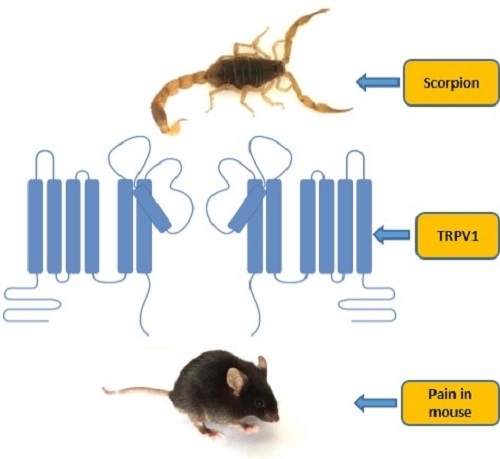
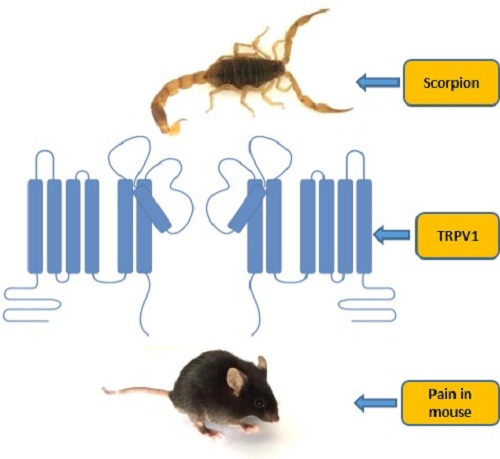
:
1. Introduction
2. Results
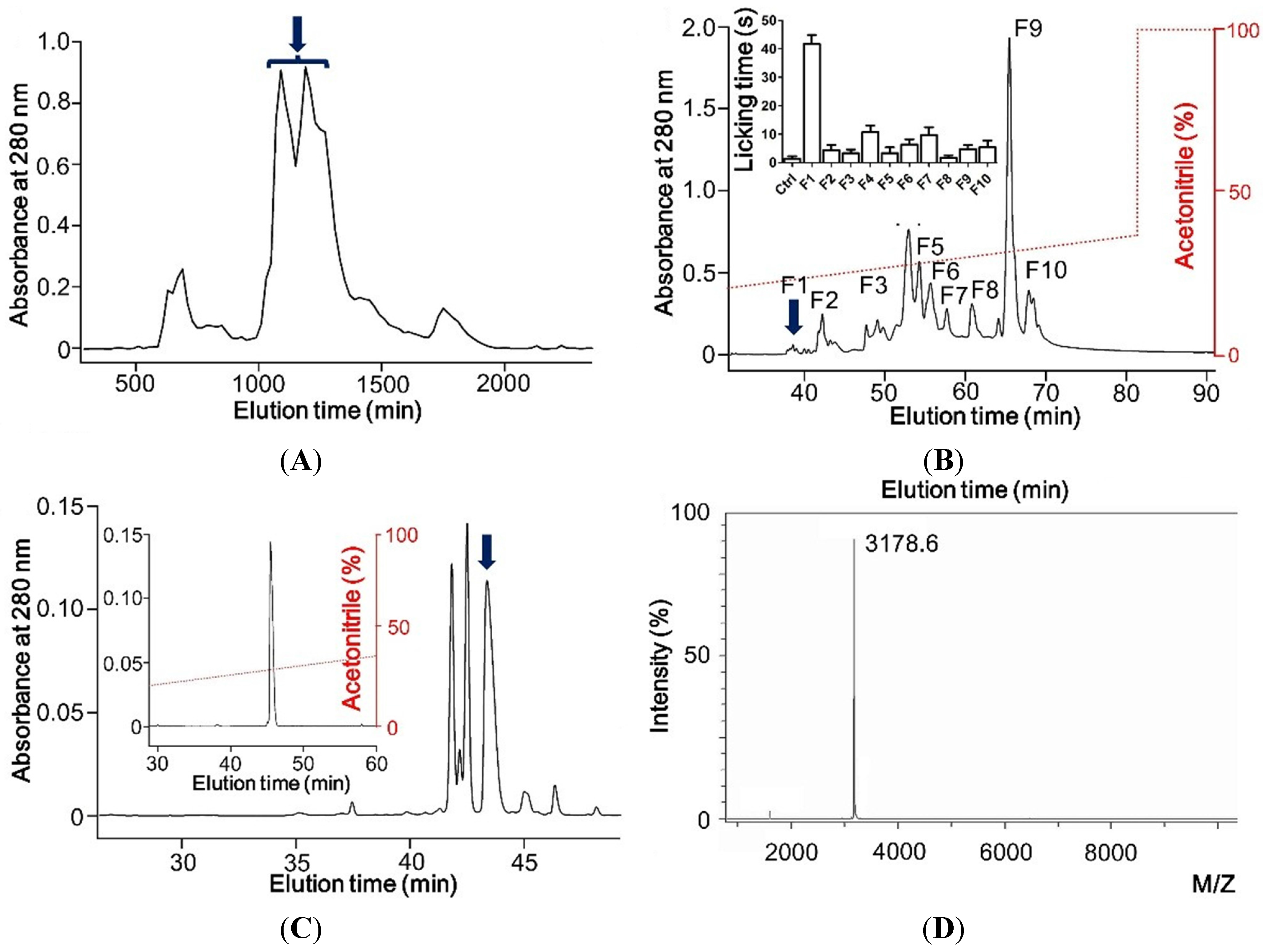
2.1. Hyrdrophobic Peptide Induces Pain in Mouse Model in Vivo

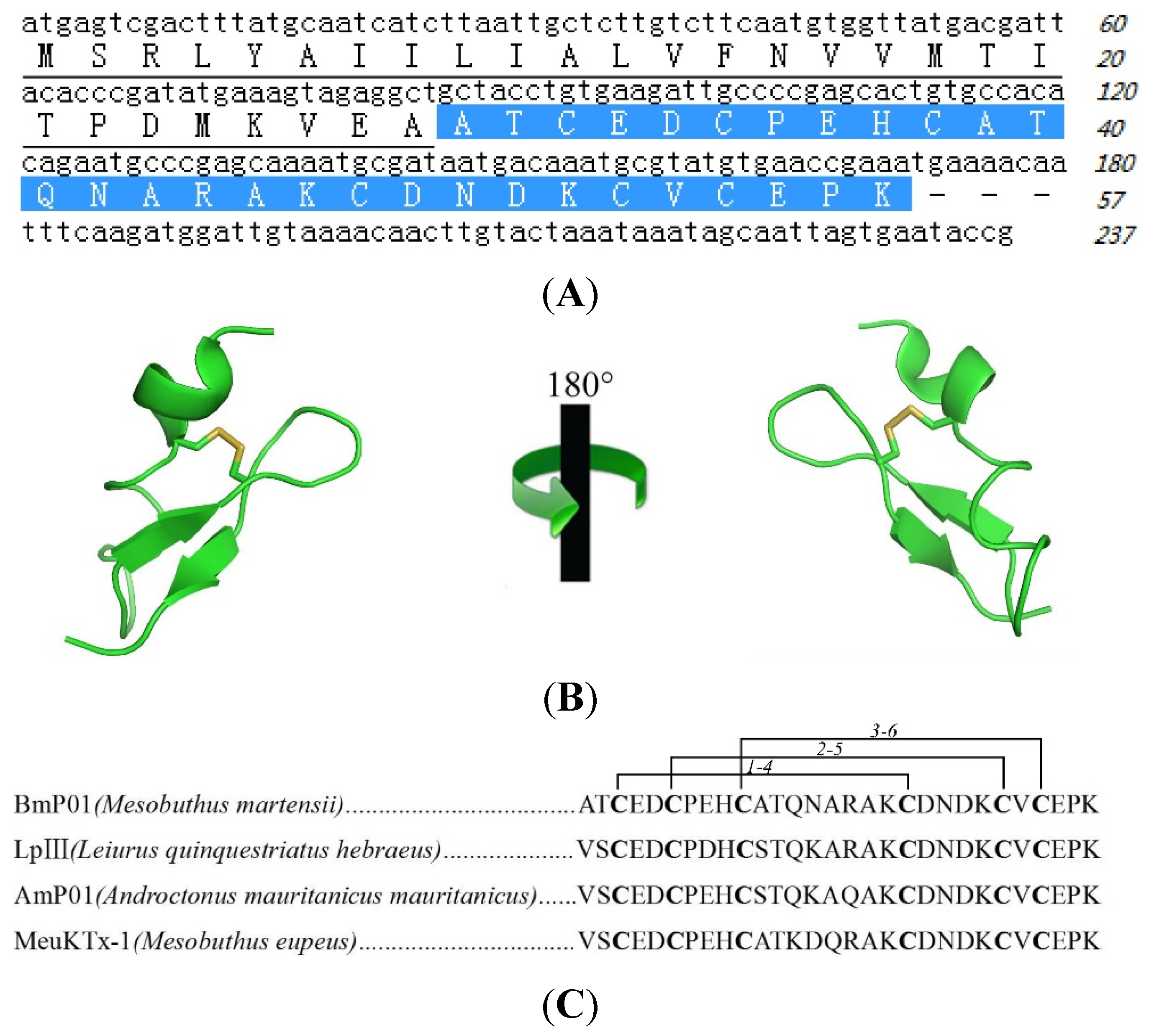
2.2. Sequence of Pain Inducing Toxin, BmP01

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| For Mature Peptide Cloning |
| Primer 1 5′-AAGCAGTGGTATCAACGCAGAGTACGCGGG-3′ |
| Primer 2 5′-GCNACNTGYG ARGAY-3′ (N = A/T/G/C; Y = C/T; R = A/G) |
| For signal peptide cloning |
| Primer 3 5-AAGCAGTGGTATCAACGCAGAGT-3 |
| Primer 4 5′-TTTCGGTTCACATAC-3′ |

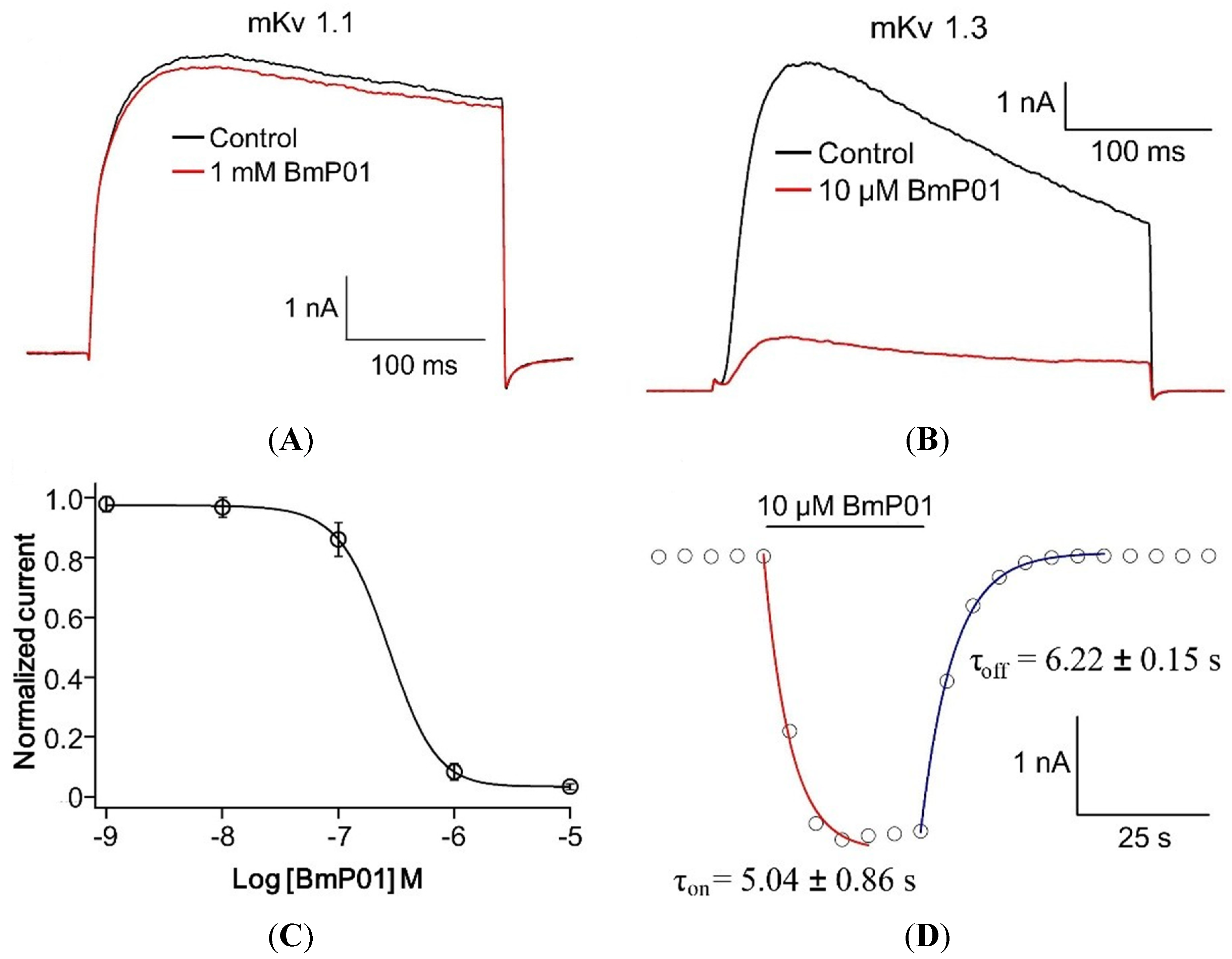
2.3. BmP01 Inhibits mKv1.3 but not mKv1.1 Channel

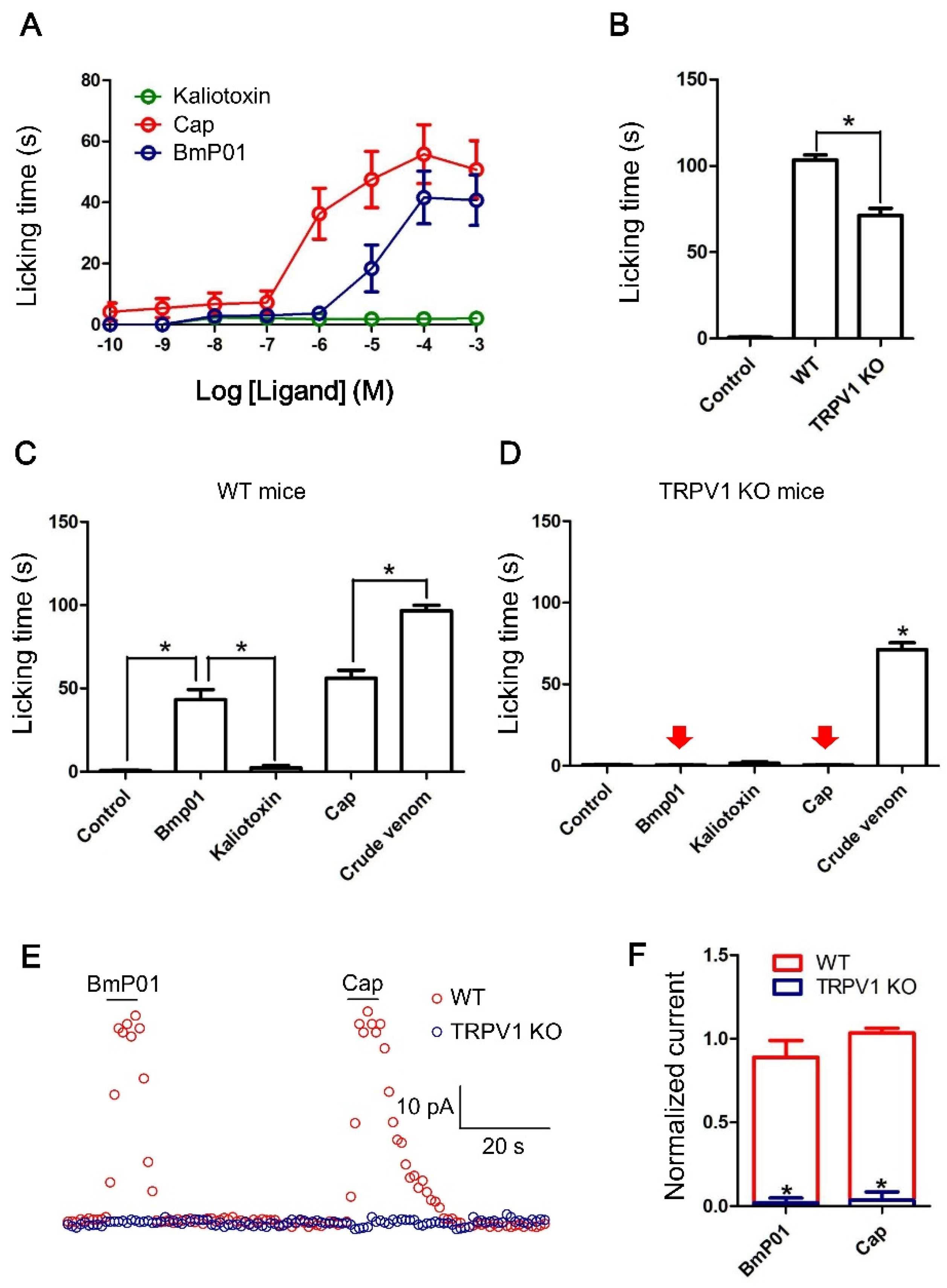
2.4. BmP01 Induces Pain in WT but not in Trpv1−/− Mice
2.5. BmP01 Evokes Currents on Sensory Neurons in WT but not in Trpv1−/− Mice
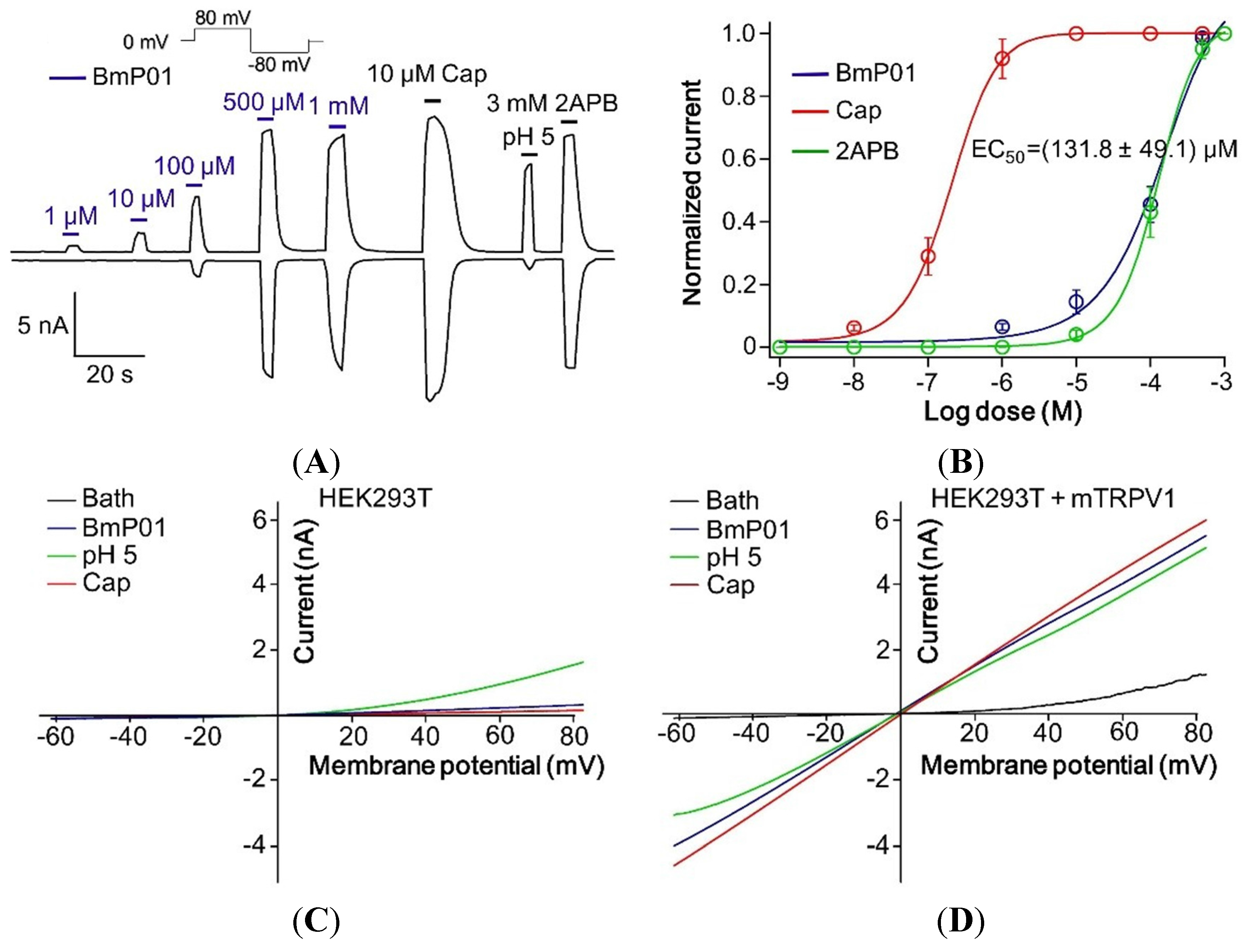
2.6. BmP01 Peptide Targets mTRPV1 Channel to Induce Pain


3. Discussion
4. Experimental Section
4.1. Venom Collection
4.2. Peptide Purification
4.3. Mass Spectrometric Analysis and Sequencing of Peptide
4.5. Cell Preparation
4.6. Electrophysiological Recordings
4.7. Animals
4.8. Paw Licking
4.9. Data Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Dunlop, J.A.; Selden, P.A. Calibrating the chelicerate clock: A paleontological reply to Jeyaprakash and Hoy. Exp. Appl. Acarol. 2009, 48, 183–197. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Yang, Y.; Dib-Hajj, S.D.; van Es, M.; Zhao, P.; Salomon, J.; Drenth, J.P.; Waxman, S.G. Depolarized inactivation overcomes impaired activation to produce DRG neuron hyperexcitability in a NaV1.7 mutation in a patient with distal limb pain. J. Neurosci. 2014, 34, 12328–12340. [Google Scholar] [CrossRef] [PubMed]
- Cao, Z.; Yu, Y.; Wu, Y.; Hao, P.; Di, Z.; He, Y.; Chen, Z.; Yang, W.; Shen, Z.; He, X.; et al. The genome of Mesobuthus martensii reveals a unique adaptation model of arthropods. Nat. Commun. 2013, 4. [Google Scholar] [CrossRef] [PubMed]
- Escoubas, P.; Diochot, S.; Corzo, G. Structure and pharmacology of spider venom neurotoxins. Biochimie 2000, 82, 893–907. [Google Scholar] [CrossRef]
- Fry, B.G.; Vidal, N.; Norman, J.A.; Vonk, F.J.; Scheib, H.; Ramjan, S.F.; Kuruppu, S.; Fung, K.; Hedges, S.B.; Richardson, M.K.; et al. Early evolution of the venom system in lizards and snakes. Nature 2006, 439, 584–588. [Google Scholar] [CrossRef] [PubMed]
- Terlau, H.; Olivera, B.M. Conus venoms: A rich source of novel ion channel-targeted peptides. Physiol. Rev. 2004, 84, 41–68. [Google Scholar] [CrossRef] [PubMed]
- Ismail, M. The scorpion envenoming syndrome. Toxicon 1995, 33, 825–858. [Google Scholar] [CrossRef]
- Almeida, D.D.; Scortecci, K.C.; Kobashi, L.S.; Agnez-Lima, L.F.; Medeiros, S.R.; Silva-Junior, A.A.; Junqueira-de-Azevedo Ide, L.; Fernandes-Pedrosa Mde, F. Profiling the resting venom gland of the scorpion tityus stigmurus through a transcriptomic survey. BMC Genomics 2012, 13. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.; Peigneur, S.; Gao, B.; Luo, L.; Jin, D.; Zhao, Y.; Tytgat, J. Molecular diversity and functional evolution of scorpion potassium channel toxins. Mol. Cell. Proteomics 2011, 10. [Google Scholar] [CrossRef] [PubMed]
- Bryson, R.W.; Riddle, B.R.; Graham, M.R.; Smith, B.T.; Prendini, L. As old as the hills: Montane scorpions in southwestern north america reveal ancient associations between biotic diversification and landscape history. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [PubMed]
- Gantenbein, B.; Largiader, C.R. The phylogeographic importance of the strait of gibraltar as a gene flow barrier in terrestrial arthropods: A case study with the scorpion Buthus occitanus as model organism. Mol. Phylogenet. Evol. 2003, 28, 119–130. [Google Scholar] [CrossRef]
- Yamashita, T.; Rhoads, D.D. Species delimitation and morphological divergence in the scorpion centruroides vittatus (say, 1821): Insights from phylogeography. PLoS ONE 2013, 8. [Google Scholar] [CrossRef]
- Zeh, D.W. The Biology of Scorpions; Polis, G.A., Ed.; Stanford University Press: Stanford, CA, USA, 1990. [Google Scholar]
- Twede, V.D.; Miljanich, G.; Olivera, B.M.; Bulaj, G. Neuroprotective and cardioprotective conopeptides: An emerging class of drug leads. Curr. Opin. Drug Discov. Dev. 2009, 12, 231–239. [Google Scholar]
- Abbas, N.; Gaudioso-Tyzra, C.; Bonnet, C.; Gabriac, M.; Amsalem, M.; Lonigro, A.; Padilla, F.; Crest, M.; Martin-Eauclaire, M.F.; Delmas, P. The scorpion toxin Amm VIII induces pain hypersensitivity through gain-of-function of TTX-sensitive Na+ channels. Pain 2013, 154, 1204–1215. [Google Scholar] [CrossRef] [PubMed]
- Bai, Z.T.; Liu, T.; Jiang, F.; Cheng, M.; Pang, X.Y.; Hua, L.M.; Shi, J.; Zhou, J.J.; Shu, X.Q.; Zhang, J.W.; et al. Phenotypes and peripheral mechanisms underlying inflammatory pain-related behaviors induced by BmK I, a modulator of sodium channels. Exp. Neurol. 2010, 226, 159–172. [Google Scholar] [CrossRef] [PubMed]
- Chippaux, J.P. Emerging options for the management of scorpion stings. Drug Des. Dev. Ther. 2012, 6, 165–173. [Google Scholar] [CrossRef] [PubMed]
- Jiang, F.; Liu, T.; Cheng, M.; Pang, X.Y.; Bai, Z.T.; Zhou, J.J.; Ji, Y.H. Spinal astrocyte and microglial activation contributes to rat pain-related behaviors induced by the venom of scorpion Buthus martensi Karch. Eur. J. Pharmacol. 2009, 623, 52–64. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Bai, Z.T.; Feng, X.H.; Li, W.P.; Yang, H.T.; Zhou, J.J.; Ji, Y.H. Hyper-excitability in low threshold mechanical a fibers is potentially involved in scorpion BmK sting pain. Brain Res. Bull. 2009, 80, 116–121. [Google Scholar] [CrossRef] [PubMed]
- Pessini, A.C.; Kanashiro, A.; Malvar Ddo, C.; Machado, R.R.; Soares, D.M.; Figueiredo, M.J.; Kalapothakis, E.; Souza, G.E. Inflammatory mediators involved in the nociceptive and oedematogenic responses induced by Tityus serrulatus scorpion venom injected into rat paws. Toxicon 2008, 52, 729–736. [Google Scholar] [CrossRef] [PubMed]
- Rowe, A.H.; Xiao, Y.; Rowe, M.P.; Cummins, T.R.; Zakon, H.H. Voltage-gated sodium channel in grasshopper mice defends against bark scorpion toxin. Science 2013, 342, 441–446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rowe, A.H.; Xiao, Y.; Scales, J.; Linse, K.D.; Rowe, M.P.; Cummins, T.R.; Zakon, H.H. Isolation and characterization of CvIV4: A pain inducing alpha-scorpion toxin. PLoS ONE 2011, 6. [Google Scholar] [CrossRef] [PubMed]
- Fry, B.G.; Roelants, K.; Champagne, D.E.; Scheib, H.; Tyndall, J.D.; King, G.F.; Nevalainen, T.J.; Norman, J.A.; Lewis, R.J.; Norton, R.S.; et al. The toxicogenomic multiverse: Convergent recruitment of proteins into animal venoms. Annu. Rev. Genomics Hum. Genet. 2009, 10, 483–511. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Liu, Z.; Xiao, Y.; Li, Y.; Rong, M.; Liang, S.; Zhang, Z.; Yu, H.; King, G.F.; Lai, R. Chemical punch packed in venoms makes centipedes excellent predators. Mol. Cell. Proteomics 2012, 11, 640–650. [Google Scholar] [CrossRef] [PubMed]
- Gangadhar, M.; Mishra, R.K.; Sriram, D.; Yogeeswari, P. Future directions in the treatment of neuropathic pain: A review on various therapeutic targets. CNS Neurol. Disord. Drug Targets 2014, 13, 63–81. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, Y.; Szallasi, A. Transient receptor potential (TRP) channels: A clinical perspective. Br. J. Pharmacol. 2014, 171, 2474–2507. [Google Scholar] [CrossRef] [PubMed]
- Kweon, H.J.; Suh, B.C. Acid-sensing ion channels (ASICs): Therapeutic targets for neurological diseases and their regulation. BMB Rep. 2013, 46, 295–304. [Google Scholar] [CrossRef] [PubMed]
- Numazaki, M.; Tominaga, M. Nociception and TRP channels. Curr. Drug Targets CNS Neurol. Disord. 2004, 3, 479–485. [Google Scholar] [CrossRef] [PubMed]
- Sekiguchi, F.; Kawabata, A. T-type calcium channels: Functional regulation and implication in pain signaling. J. Pharmacol. Sci. 2013, 122, 244–250. [Google Scholar] [CrossRef] [PubMed]
- Holzer, P. TRPV1 and the gut: From a tasty receptor for a painful vanilloid to a key player in hyperalgesia. Eur. J. Pharmacol. 2004, 500, 231–241. [Google Scholar] [CrossRef] [PubMed]
- Bohlen, C.J.; Priel, A.; Zhou, S.; King, D.; Siemens, J.; Julius, D. A bivalent tarantula toxin activates the capsaicin receptor, TRPV1, by targeting the outer pore domain. Cell 2010, 141, 834–845. [Google Scholar] [CrossRef] [PubMed]
- Siemens, J.; Zhou, S.; Piskorowski, R.; Nikai, T.; Lumpkin, E.A.; Basbaum, A.I.; King, D.; Julius, D. Spider toxins activate the capsaicin receptor to produce inflammatory pain. Nature 2006, 444, 208–212. [Google Scholar] [CrossRef] [PubMed]
- Bohlen, C.J.; Julius, D. Receptor-targeting mechanisms of pain-causing toxins: How ow? Toxicon 2012, 60, 254–264. [Google Scholar] [CrossRef] [PubMed]
- Romi-Lebrun, R.; Martin-Eauclaire, M.F.; Escoubas, P.; Wu, F.Q.; Lebrun, B.; Hisada, M.; Nakajima, T. Characterization of four toxins from Buthus martensi scorpion venom, which act on apamin-sensitive Ca2+-activated K+ channels. Eur. J. Biochem. 1997, 245, 457–464. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.J.; Dai, L.; Lan, Z.D.; Chi, C.W. Genomic organization of three neurotoxins active on small conductance Ca2+-activated potassium channels from the scorpion Buthus martensi Karsch. FEBS Lett. 1999, 452, 360–364. [Google Scholar] [CrossRef]
- Wu, G.; Li, Y.; Wei, D.; He, F.; Jiang, S.; Hu, G.; Wu, H. Solution structure of Bmp01 from the venom of scorpion Buthus martensii Karsch. Biochem. Biophys. Res. Commun. 2000, 276, 1148–1154. [Google Scholar] [CrossRef] [PubMed]
- Kourrich, S.; Mourre, C.; Soumireu-Mourat, B. Kaliotoxin, a KV1.1 and KV1.3 channel blocker, improves associative learning in rats. Behav. Brain Res. 2001, 120, 35–46. [Google Scholar] [CrossRef]
- Mourre, C.; Chernova, M.N.; Martin-Eauclaire, M.F.; Bessone, R.; Jacquet, G.; Gola, M.; Alper, S.L.; Crest, M. Distribution in rat brain of binding sites of kaliotoxin, a blocker of KV1.1 and KV1.3 alpha-subunits. J. Pharmacol. Exp. Ther. 1999, 291, 943–952. [Google Scholar] [PubMed]
- Gunthorpe, M.J.; Smith, G.D.; Davis, J.B.; Randall, A.D. Characterisation of a human acid-sensing ion channel (hASIC1a) endogenously expressed in HEK293 cells. Pflugers Arch.: Eur. J. Physiol. 2001, 442, 668–674. [Google Scholar]
- Nicholson, G.M. Insect-selective spider toxins targeting voltage-gated sodium channels. Toxicon 2007, 49, 490–512. [Google Scholar] [CrossRef] [PubMed]
- Jiang, F.; Hua, L.M.; Jiao, Y.L.; Ye, P.; Fu, J.; Cheng, Z.J.; Ding, G.; Ji, Y.H. Activation of mammalian target of rapamycin contributes to pain nociception induced in rats by BmK I, a sodium channel-specific modulator. Neurosci. Bull. 2014, 30, 21–32. [Google Scholar] [CrossRef] [PubMed]
- Jordt, S.E.; Julius, D. Molecular basis for species-specific sensitivity to “hot” chili peppers. Cell 2002, 108, 421–430. [Google Scholar] [CrossRef]
- Diochot, S.; Baron, A.; Salinas, M.; Douguet, D.; Scarzello, S.; Dabert-Gay, A.S.; Debayle, D.; Friend, V.; Alloui, A.; Lazdunski, M.; et al. Black mamba venom peptides target acid-sensing ion channels to abolish pain. Nature 2012, 490, 552–555. [Google Scholar] [CrossRef] [PubMed]
- You, D.; Hong, J.; Rong, M.; Yu, H.; Liang, S.; Ma, Y.; Yang, H.; Wu, J.; Lin, D.; Lai, R. The first gene-encoded amphibian neurotoxin. J. Biol. Chem. 2009, 284, 22079–22086. [Google Scholar] [CrossRef] [PubMed]
- Gui, J.; Liu, B.; Cao, G.; Lipchik, A.M.; Perez, M.; Dekan, Z.; Mobli, M.; Daly, N.L.; Alewood, P.F.; Parker, L.L.; et al. A tarantula-venom peptide antagonizes the TRPA1 nociceptor ion channel by binding to the S1–S4 gating domain. Curr. Biol.: CB 2014, 24, 473–483. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.; Peigneur, S.; Gao, B.; Lu, X.; Cao, C.; Tytgat, J. Evolutionary diversification of Mesobuthus alpha-scorpion toxins affecting sodium channels. Mol. Cell. Proteomics 2012, 11. [Google Scholar] [CrossRef] [PubMed]
- Giangiacomo, K.M.; Ceralde, Y.; Mullmann, T.J. Molecular basis of alpha-KTx specificity. Toxicon 2004, 43, 877–886. [Google Scholar] [CrossRef] [PubMed]
- Mouhat, S.; Andreotti, N.; Jouirou, B.; Sabatier, J.M. Animal toxins acting on voltage-gated potassium channels. Curr. Pharm. Des. 2008, 14, 2503–2518. [Google Scholar] [CrossRef] [PubMed]
- Puopolo, M.; Binshtok, A.M.; Yao, G.L.; Oh, S.B.; Woolf, C.J.; Bean, B.P. Permeation and block of TRPV1 channels by the cationic lidocaine derivative QX-314. J. Neurophysiol. 2013, 109, 1704–1712. [Google Scholar] [CrossRef] [PubMed]
- Roberson, D.P.; Binshtok, A.M.; Blasl, F.; Bean, B.P.; Woolf, C.J. Targeting of sodium channel blockers into nociceptors to produce long-duration analgesia: A systematic study and review. Br. J. Pharmacol. 2011, 164, 48–58. [Google Scholar] [CrossRef] [PubMed]
- Fedosov, A.E.; Moshkovskii, S.A.; Kuznetsova, K.G.; Olivera, B.M. conotoxins: From the biodiversity of gastropods to new drugs. Biomed. Khimiia 2013, 59, 267–294. [Google Scholar] [CrossRef]
- Yang, S.; Xiao, Y.; Kang, D.; Liu, J.; Li, Y.; Undheim, E.A.; Klint, J.K.; Rong, M.; Lai, R.; King, G.F. Discovery of a selective NaV1.7 inhibitor from centipede venom with analgesic efficacy exceeding morphine in rodent pain models. Proc. Natl. Acad. Sci. USA 2013, 110, 17534–17539. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hakim, M.A.; Jiang, W.; Luo, L.; Li, B.; Yang, S.; Song, Y.; Lai, R. Scorpion Toxin, BmP01, Induces Pain by Targeting TRPV1 Channel. Toxins 2015, 7, 3671-3687. https://doi.org/10.3390/toxins7093671
Hakim MA, Jiang W, Luo L, Li B, Yang S, Song Y, Lai R. Scorpion Toxin, BmP01, Induces Pain by Targeting TRPV1 Channel. Toxins. 2015; 7(9):3671-3687. https://doi.org/10.3390/toxins7093671
Chicago/Turabian StyleHakim, Md Abdul, Wenbin Jiang, Lei Luo, Bowen Li, Shilong Yang, Yuzhu Song, and Ren Lai. 2015. "Scorpion Toxin, BmP01, Induces Pain by Targeting TRPV1 Channel" Toxins 7, no. 9: 3671-3687. https://doi.org/10.3390/toxins7093671