
Maternal and Neonatal Polyunsaturated Fatty Acid Intake and Risk of Neurodevelopmental Impairment in Premature Infants


{kind=link}
Abstract
:1. Introduction
2. Why Are DHA and AA Important in the Developing Brain?
2.1. LCPUFA and Neurodevelopment
2.2. LCPUFA Influence Inflammatory Signaling
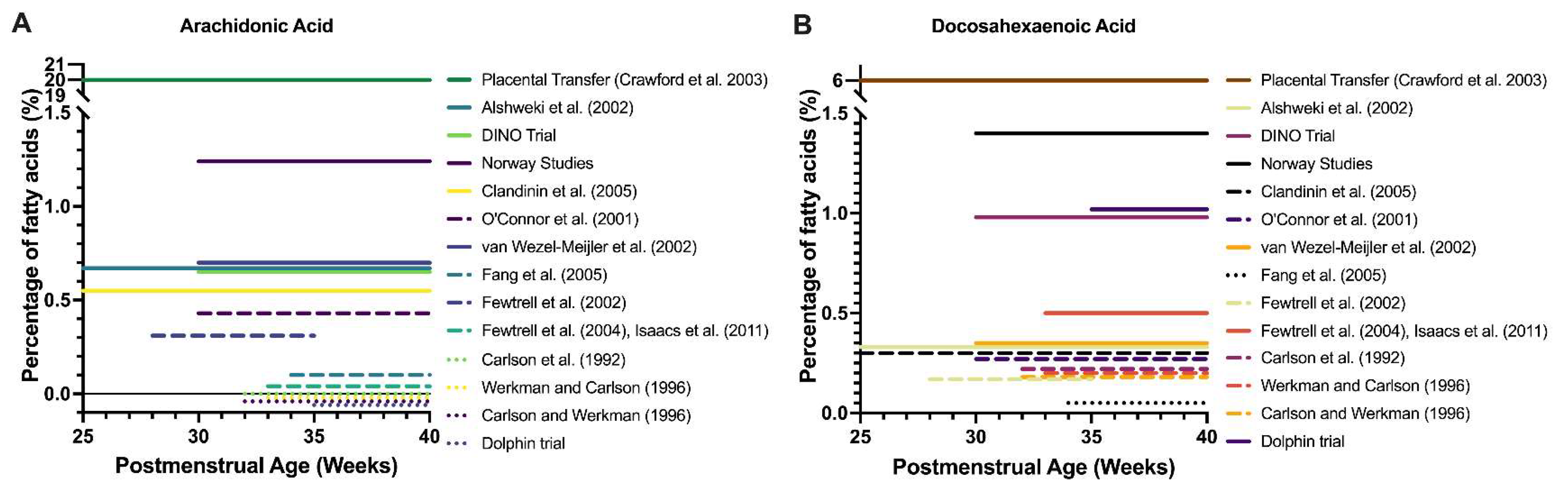
3. LCPUFA Accretion into Fetal Tissues
3.1. In Utero LCPUFA Accretion
3.2. Post-Natal LCPUFA Accretion
3.3. LCPUFA Accretion into Adipose
4. What Is the Evidence Linking Maternal and Neonatal PUFA Intake with Neurodevelopmental Outcomes?
4.1. Randomized Controlled Trials
4.2. Reviews and Systematic Reviews
5. Can the Evidence Inform Future LCPUFA Supplementation Strategies?
5.1. Is Supplemental LCPUFA Better than None at All?
5.2. Would Higher Doses of Supplemental LCPUFA Provide Greater Benefits to Neurodevelopment?
5.3. Which Preterm Infant Population May Benefit Most from LCPUFA Supplementation?
5.4. Tailoring Supplemental DHA and AA to Mimic Dynamic Physiological Placental Supply and Developmental Demand
5.5. Organ-Specific Effects of LCPUFA
5.6. Challenges for Appropriate LCPUFA Supplementation
6. The Wider Contexts of LCPUFA Supplementation
Dietary LA Affects Infant AA and DHA Availability
7. The Wider Factors Affecting LCPUFA Status
7.1. Maternal LCPUFA Status
7.2. Genetic Polymorphisms
7.3. Socioeconomic Status
8. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Xie, L.; Innis, S.M. Genetic Variants of the FADS1 FADS2 Gene Cluster Are Associated with Altered (n-6) and (n-3) Essential Fatty Acids in Plasma and Erythrocyte Phospholipids in Women during Pregnancy and in Breast Milk during Lactation. J. Nutr. 2008, 138, 2222–2228. [Google Scholar] [CrossRef] [Green Version]
- Carnielli, V.P.; Wattimena, D.J.L.; Luijendijk, I.H.T.; Boerlage, A.; Degenhart, H.J.; Sauer, P.J.J. The Very Low Birth Weight Premature Infant Is Capable of Synthesizing Arachidonic and Docosahexaenoic Acids from Linoleic and Linolenic Acids. Pediatr. Res. 1996, 40, 169–174. [Google Scholar] [CrossRef] [Green Version]
- Carnielli, V.P.; Simonato, M.; Verlato, G.; Luijendijk, I.; de Curtis, M.; Sauer, P.J.J.; Cogo, P.E. Synthesis of Long-Chain Polyunsaturated Fatty Acids in Preterm Newborns Fed Formula with Long-Chain Polyunsaturated Fatty Acids. Am. J. Clin. Nutr. 2007, 86, 1323–1330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agostoni, C.; Buonocore, G.; Carnielli, V.P.; de Curtis, M.; Darmaun, D.; Decsi, T.; Domellöf, M.; Embleton, N.D.; Fusch, C.; Genzel-Boroviczeny, O.; et al. Enteral Nutrient Supply for Preterm Infants: Commentary from the European Society of Paediatric Gastroenterology, Hepatology and Nutrition Committee on Nutrition. J. Pediatr. Gastroenterol. Nutr. 2010, 50, 85–91. [Google Scholar] [CrossRef]
- Salem, N.; Wegher, B.; Mena, P.; Uauy, R. Arachidonic and Docosahexaenoic Acids Are Biosynthesized from Their 18-Carbon Precursors in Human Infants. Proc. Natl. Acad. Sci. USA 1996, 93, 49–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moon, K.; Rao, S.C.; Schulzke, S.M.; Patole, S.K.; Simmer, K. Longchain Polyunsaturated Fatty Acid Supplementation in Preterm Infants. Cochrane Database Syst. Rev. 2016, 2017. [Google Scholar] [CrossRef]
- Almaas, A.N.; Tamnes, C.K.; Nakstad, B.; Henriksen, C.; Walhovd, K.B.; Fjell, A.M.; Due-Tonnessen, P.; Drevon, C.A.; Iversen, P.O. Long-Chain Polyunsaturated Fatty Acids and Cognition in VLBW Infants at 8 Years: An RCT. Pediatrics 2015, 135, 972–980. [Google Scholar] [CrossRef] [Green Version]
- Alshweki, A.; Muñuzuri, A.P.; Baña, A.M.; José De Castro, M.; Andrade, F.; Aldamiz-Echevarría, L.; Sáenz De Pipaón, M.; Fraga, J.M.; Couce, M.L. Effects of Different Arachidonic Acid Supplementation on Psychomotor Development in Very Preterm Infants; a Randomized Controlled Trial. Nutr. J. 2015, 14, 101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collins, C.T.; Gibson, R.A.; Anderson, P.J.; McPhee, A.J.; Sullivan, T.R.; Gould, J.F.; Ryan, P.; Doyle, L.W.; Davis, P.G.; McMichael, J.E.; et al. Neurodevelopmental Outcomes at 7 Years’ Corrected Age in Preterm Infants Who Were Fed High-Dose Docosahexaenoic Acid to Term Equivalent: A Follow-up of a Randomised Controlled Trial. BMJ Open 2015, 5, e007314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makrides, M.; Gibson, R.A.; McPhee, A.J.; Collins, C.T.; Davis, P.G.; Doyle, L.W.; Simmer, K.; Colditz, P.B.; Morris, S.; Smithers, L.G.; et al. Neurodevelopmental Outcomes of Preterm Infants Fed High-Dose Docosahexaenoic Acid. JAMA 2009, 301, 175. [Google Scholar] [CrossRef] [Green Version]
- Smithers, L.G.; Collins, C.T.; Simmonds, L.A.; Gibson, R.A.; McPhee, A.; Makrides, M. Feeding Preterm Infants Milk with a Higher Dose of Docosahexaenoic Acid than That Used in Current Practice Does Not Influence Language or Behavior in Early Childhood: A Follow-up Study of a Randomized Controlled Trial. Am. J. Clin. Nutr. 2010, 91, 628–634. [Google Scholar] [CrossRef] [PubMed]
- Hadley, K.B.; Ryan, A.S.; Forsyth, S.; Gautier, S.; Salem, N. The Essentiality of Arachidonic Acid in Infant Development. Nutrients 2016, 8, 216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashimoto, M.; Hossain, S.; Shimada, T.; Shido, O. Docosahexaenoic Acid-Induced Protective Effect against Impaired Learning in Amyloid β-Infused Rats Is Associated with Increased Synaptosomal Membrane Fluidity. Clin. Exp. Pharmacol. Physiol. 2006, 33, 934–939. [Google Scholar] [CrossRef]
- Onuki, Y.; Morishita, M.; Chiba, Y.; Tokiwa, S.; Takayama, K. Docosahexaenoic Acid and Eicosapentaenoic Acid Induce Changes in the Physical Properties of a Lipid Bilayer Model Membrane. Chem. Pharm. Bull. 2006, 54, 68–71. [Google Scholar] [CrossRef] [Green Version]
- Berman, D.R.; Liu, Y.; Barks, J.; Mozurkewich, E. Treatment with Docosahexaenoic Acid after Hypoxia-Ischemia Improves Forepaw Placing in a Rat Model of Perinatal Hypoxia-Ischemia. Am. J. Obstet. Gynecol. 2010, 203, 385-e1. [Google Scholar] [CrossRef] [Green Version]
- Buddington, R.K.; Chizhikov, V.V.; Iskusnykh, I.Y.; Sable, H.J.; Sable, J.J.; Holloway, Z.R.; Katzir, T.B.; van der Merwe, M.; Yakimkova, T.; Buddington, K.K.; et al. A Phosphatidylserine Source of Docosahexanoic Acid Improves Neurodevelopment and Survival of Preterm Pigs. Nutrients 2018, 10, 637. [Google Scholar] [CrossRef] [Green Version]
- Suganuma, H.; Arai, Y.; Kitamura, Y.; Hayashi, M.; Okumura, A.; Shimizu, T. Maternal Docosahexaenoic Acid-Enriched Diet Prevents Neonatal Brain Injury. Neuropathology 2010, 30, 597–605. [Google Scholar] [CrossRef] [PubMed]
- Farquharson, J.; Jamieson, E.C.; Logan, R.W.; Cockburn, F.; Ainslie Patrick, W. Infant Cerebral Cortex Phospholipid Fatty-Acid Composition and Diet. Lancet 1992, 340, 810–813. [Google Scholar] [CrossRef]
- Farquharson, J.; Jamieson, E.C.; Abbasi, K.A.; Patrick, W.J.A.; Logan, R.W.; Cockbum, F. Effect of Diet on the Fatty Acid Composition of the Major Phospholipids of Infant Cerebral Cortex. Arch. Dis. Child. 1995, 72, 198–203. [Google Scholar] [CrossRef]
- Uauy, R.D.; Birch, D.G.; Birch, E.E.; Tyson, J.E.; Hoffman, D.R. Effect of Dietary Omega-3 Fatty Acids on Retinal Function of Very-Low-Birth-Weight Neonates. Pediatr. Res. 1990, 28, 485–492. [Google Scholar] [CrossRef] [PubMed]
- Hellström, A.; Nilsson, A.K.; Wackernagel, D.; Pivodic, A.; Vanpee, M.; Sjöbom, U.; Hellgren, G.; Hallberg, B.; Domellöf, M.; Klevebro, S.; et al. Effect of Enteral Lipid Supplement on Severe Retinopathy of Prematurity: A Randomized Clinical Trial. JAMA Pediatr. 2021, 175, 359–367. [Google Scholar] [CrossRef]
- Bernabe-García, M.; Villegas-Silva, R.; Villavicencio-Torres, A.; Calder, P.C.; Rodríguez-Cruz, M.; Maldonado-Hernández, J.; Macías-Loaiza, D.; López-Alarcón, M.; Inda-Icaza, P.; Cruz-Reynoso, L. Enteral Docosahexaenoic Acid and Retinopathy of Prematurity: A Randomized Clinical Trial Mariela Bernabe-García. J. Parenter. Enter. Nutr. 2019, 43, 874–882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sveinsdóttir, K.; Ley, D.; Hövel, H.; Fellman, V.; Hüppi, P.S.; Smith, L.E.H.; Hellström, A.; Hansen Pupp, I. Relation of Retinopathy of Prematurity to Brain Volumes at Term Equivalent Age and Developmental Outcome at 2 Years of Corrected Age in Very Preterm Infants. Neonatology 2018, 114, 46–52. [Google Scholar] [CrossRef]
- Tam, E.W.Y.; Chau, V.; Barkovich, A.J.; Ferriero, D.M.; Miller, S.P.; Rogers, E.E.; Grunau, R.E.; Synnes, A.R.; Xu, D.; Foong, J.; et al. Early Postnatal Docosahexaenoic Acid Levels and Improved Preterm Brain Development. Pediatr. Res. 2016, 79, 723–730. [Google Scholar] [CrossRef] [Green Version]
- Carlson, S.E.; Cooke, R.J.; Werkman, S.H.; Tolley, E.A. First Year Growth of Preterm Infants Fed Standard Compared to Marine Oil N-3 Supplemented Formula. Lipids 1992, 27, 901–907. [Google Scholar] [CrossRef]
- Carlson, S.E.; Werkman, S.H.; Peeples, J.M.; Wilson, W.M. Growth and Development of Premature Infants in Relation to Omega 3 and Omega 6 Fatty Acid Status. World Rev. Nutr. Diet. 1994, 75, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Hack, M.; Flannery, D.J.; Schluchter, M.; Cartar, L.; Borawski, E.; Klein, N. Outcomes in Young Adulthood for Very-Low-Birth-Weight Infants. N. Engl. J. Med. 2002, 346, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Calder, P.C. Polyunsaturated Fatty Acids and Inflammation: From Molecular Biology to the Clinic. Lipids 2003, 38, 343–352. [Google Scholar] [CrossRef] [PubMed]
- Levy, B.D.; Clish, C.B.; Schmidt, B.; Gronert, K.; Serhan, C.N. Lipid Mediator Class Switching during Acute Inflammation: Signals in Resolution. Nat. Immunol. 2001, 2, 612–619. [Google Scholar] [CrossRef] [PubMed]
- Serhan, C.N.; Chiang, N.; Dalli, J.; Levy, B.D. Lipid Mediators in the Resolution of Inflammation. Cold Spring Harb. Perspect. Biol. 2015, 7, a016311. [Google Scholar] [CrossRef] [Green Version]
- Serhan, C.N.; Chiang, N.; van Dyke, T.E. Resolving Inflammation: Dual Anti-Inflammatory and pro-Resolution Lipid Mediators. Nat. Rev. Immunol. 2008, 8, 349–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bazan, N.G.; Calandria, J.M.; Gordon, W.C. Docosahexaenoic Acid and Its Derivative Neuroprotectin D1 Display Neuroprotective Properties in the Retina, Brain and Central Nervous System. Nestle Nutr. Inst. Workshop Ser. 2013, 77, 121–131. [Google Scholar] [CrossRef] [PubMed]
- Humberg, A.; Fortmann, I.; Siller, B.; Kopp, M.V.; Herting, E.; Göpel, W.; Härtel, C. Preterm Birth and Sustained Inflammation: Consequences for the Neonate. Semin. Immunopathol. 2020, 42, 451–468. [Google Scholar] [CrossRef]
- Martin, C.R.; Dasilva, D.A.; Cluette-Brown, J.E.; Dimonda, C.; Hamill, A.; Bhutta, A.Q.; Coronel, E.; Wilschanski, M.; Stephens, A.J.; Driscoll, D.F.; et al. Decreased Postnatal Docosahexaenoic and Arachidonic Acid Blood Levels in Premature Infants Are Associated with Neonatal Morbidities. J. Pediatr. 2011, 159, 743–749.e2. [Google Scholar] [CrossRef] [Green Version]
- Collins, C.T.; Makrides, M.; McPhee, A.J.; Sullivan, T.R.; Davis, P.G.; Thio, M.; Simmer, K.; Rajadurai, V.S.; Travadi, J.; Berry, M.J.; et al. Docosahexaenoic Acid and Bronchopulmonary Dysplasia in Preterm Infants. N. Engl. J. Med. 2017, 376, 1245–1255. [Google Scholar] [CrossRef]
- Marc, I.; Piedboeuf, B.; Lacaze-Masmonteil, T.; Fraser, W.; Mâsse, B.; Mohamed, I.; Qureshi, M.; Afifi, J.; Lemyre, B.; Caouette, G.; et al. Effect of Maternal Docosahexaenoic Acid Supplementation on Bronchopulmonary Dysplasia-Free Survival in Breastfed Preterm Infants: A Randomized Clinical Trial. JAMA—J. Am. Med. Assoc. 2020, 324, 157–167. [Google Scholar] [CrossRef] [PubMed]
- Hellström, A.; Pivodic, A.; Gränse, L.; Lundgren, P.; Sjöbom, U.; Nilsson, A.K.; Söderling, H.; Hård, A.-L.; Smith, L.E.H.; Löfqvist, C.A. Association of Docosahexaenoic Acid and Arachidonic Acid Serum Levels With Retinopathy of Prematurity in Preterm Infants. JAMA Netw. Open 2021, 4, e2128771. [Google Scholar] [CrossRef]
- Wu, W.X.; Ma, X.H.; Coksaygan, T.; Chakrabarty, K.; Collins, V.; Rose, J.; Nathanielsz, P.W. Prostaglandin Mediates Premature Delivery in Pregnant Sheep Induced by Estradiol at 121 Days of Gestational Age. Endocrinology 2004, 145, 1444–1452. [Google Scholar] [CrossRef] [Green Version]
- Olsen, S.F.; Halldorsson, T.I.; Thorne-Lyman, A.L.; Strøm, M.; Gørtz, S.; Granstrøm, C.; Nielsen, P.H.; Wohlfahrt, J.; Lykke, J.A.; Langhoff-Roos, J.; et al. Plasma Concentrations of Long Chain N-3 Fatty Acids in Early and Mid-Pregnancy and Risk of Early Preterm Birth. EBioMedicine 2018, 35, 325–333. [Google Scholar] [CrossRef] [Green Version]
- Hellström, A.; Hellström, W.; Hellgren, G.; Smith, L.E.H.; Puttonen, H.; Fyhr, I.M.; Sävman, K.; Nilsson, A.K.; Klevebro, S. Docosahexaenoic Acid and Arachidonic Acid Levels Are Associated with Early Systemic Inflammation in Extremely Preterm Infants. Nutrients 2020, 12, 1996. [Google Scholar] [CrossRef]
- Hofer, N.; Kothari, R.; Morris, N.; Müller, W.; Resch, B. The Fetal Inflammatory Response Syndrome Is a Risk Factor for Morbidity in Preterm Neonates. Am. J. Obstet. Gynecol. 2013, 209, 542.e1–542.e11. [Google Scholar] [CrossRef]
- Kuban, K.C.K.; Joseph, R.M.; O’Shea, T.M.; Heeren, T.; Fichorova, R.N.; Douglass, L.; Jara, H.; Frazier, J.A.; Hirtz, D.; Rollins, J.V.; et al. Circulating Inflammatory-Associated Proteins in the First Month of Life and Cognitive Impairment at Age 10 Years in Children Born Extremely Preterm. J. Pediatr. 2017, 180, 116–123.e1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crawford, M.A.; Williams, G.; Hassam, A.G.; Whitehouse, W.L. Essential Fatty Acids and Fetal Brain Growth. Lancet 1976, 307, 452–453. [Google Scholar] [CrossRef]
- Hüppi, P.S.; Warfield, S.; Kikinis, R.; Barnes, P.D.; Zientara, G.P.; Jolesz, F.A.; Tsuji, M.K.; Volpe, J.J. Quantitative Magnetic Resonance Imaging of Brain Development in Premature and Mature Newborns. Ann. Neurol. 1998, 43, 224–235. [Google Scholar] [CrossRef]
- Clandinin, M.T.; Chappell, J.E.; Leong, S.; Heim, T.; Swyer, P.R.; Chance, G.W. Intrauterine Fatty Acid Accretion Rates in Human Brain: Implications for Fatty Acid Requirements. Early Hum. Dev. 1980, 4, 121–129. [Google Scholar] [CrossRef]
- Bernhard, W.; Raith, M.; Koch, V.; Maas, C.; Abele, H.; Poets, C.F.; Franz, A.R. Developmental Changes in Polyunsaturated Fetal Plasma Phospholipids and Feto-Maternal Plasma Phospholipid Ratios and Their Association with Bronchopulmonary Dysplasia. Eur. J. Nutr. 2016, 55, 2265–2274. [Google Scholar] [CrossRef] [PubMed]
- Kuipers, R.S.; Luxwolda, M.F.; Offringa, P.J.; Rudi Boersma, E.; Dijck-Brouwer, D.A.J.; Muskiet, F.A.J. Fetal Intrauterine Whole Body Linoleic, Arachidonic and Docosahexaenoic Acid Contents and Accretion Rates. Prostaglandins Leukot. Essent. Fat. Acids 2012, 86, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Kuipers, R.S.; Luxwolda, M.F.; Offringa, P.J.; Rudy Boersma, E.; Janneke Dijck-Brouwer, D.A.; Muskiet, F.A.J. Gestational Age Dependent Changes of the Fetal Brain, Liver and Adipose Tissue Fatty Acid Compositions in a Population with High Fish Intakes. Prostaglandins Leukot. Essent. Fat. Acids 2012, 86, 189–199. [Google Scholar] [CrossRef]
- Martínez, M.; Mougan, I. Fatty Acid Composition of Human Brain Phospholipids during Normal Development. J. Neurochem. 1998, 71, 2528–2533. [Google Scholar] [CrossRef] [Green Version]
- Innis, S.M.; Kuhnlein, H.V. Long-Chain n-3 Fatty Acids in Breast Milk of Inuit Women Consuming Traditional Foods. Early Hum. Dev. 1988, 18, 185–189. [Google Scholar] [CrossRef]
- Cunnane, S.C.; Francescutti, V.; Brenna, J.T.; Crawford, M.A. Breast-Fed Infants Achieve a Higher Rate of Brain and Whole Body Docosahexaenoate Accumulation than Formula-Fed Infants Not Consuming Dietary Docosahexaenoate. Lipids 2000, 35, 105–111. [Google Scholar] [CrossRef]
- Clandinin, M.T.; Chappell, J.E.; Leong, S.; Heim, T.; Swyer, P.R.; Chance, G.W. Extrauterine Fatty Acid Accretion in Infant Brain: Implications for Fatty Acid Requirements. Early Hum. Dev. 1980, 4, 131–138. [Google Scholar] [CrossRef]
- Clandinin, M.T.; Chappell, J.E.; Heim, T.; Swyer, P.R.; Chance, G.W. Fatty Acid Utilization in Perinatal de Novo Synthesis of Tissues. Early Hum. Dev. 1981, 5, 355–366. [Google Scholar] [CrossRef]
- Cunnane, S.C.; Crawford, M.A. Survival of the Fattest: Fat Babies Were the Key to Evolution of the Large Human Brain. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2003, 136, 17–26. [Google Scholar] [CrossRef]
- Innis, S.M. Essential Fatty Acid Transfer and Fetal Development. Placenta 2005, 26, 4–9. [Google Scholar] [CrossRef]
- Henriksen, C.; Haugholt, K.; Lindgren, M.; Aurvåg, A.K.; Rønnestad, A.; Grønn, M.; Solberg, R.; Moen, A.; Nakstad, B.; Berge, R.K.; et al. Improved Cognitive Development among Preterm Infants Attributable to Early Supplementation of Human Milk with Docosahexaenoic Acid and Arachidonic Acid. Pediatrics 2008, 121, 1137–1145. [Google Scholar] [CrossRef] [PubMed]
- Westerberg, A.C.; Schei, R.; Henriksen, C.; Smith, L.; Veierød, M.B.; Drevon, C.A.; Iversen, P.O. Attention among Very Low Birth Weight Infants Following Early Supplementation with Docosahexaenoic and Arachidonic Acid. Acta Paediatr. Int. J. Paediatr. 2011, 100, 47–52. [Google Scholar] [CrossRef]
- Almaas, A.N.; Tamnes, C.K.; Nakstad, B.; Henriksen, C.; Grydeland, H.; Walhovd, K.B.; Fjell, A.M.; Iversen, P.O.; Drevon, C.A. Diffusion Tensor Imaging and Behavior in Premature Infants at 8 Years of Age, a Randomized Controlled Trial with Long-Chain Polyunsaturated Fatty Acids. Early Hum. Dev. 2016, 95, 41–46. [Google Scholar] [CrossRef]
- Henriksen, C.; Almaas, A.N.; Westerberg, A.C.; Drevon, C.A.; Iversen, P.O.; Nakstad, B. Growth, Metabolic Markers, and Cognition in 8-Year Old Children Born Prematurely, Follow-up of a Randomized Controlled Trial with Essential Fatty Acids. Eur. J. Pediatr. 2016, 175, 1165–1174. [Google Scholar] [CrossRef]
- Clandinin, M.T.; van Aerde, J.E.; Merkel, K.L.; Harris, C.L.; Springer, M.A.; Hansen, J.W.; Diersen-Schade, D.A. Growth and Development of Preterm Infants Fed Infant Formulas Containing Docosahexaenoic Acid and Arachidonic Acid. J. Pediatr. 2005, 146, 461–468. [Google Scholar] [CrossRef]
- O’Connor, D.L.; Hall, R.; Adamkin, D.; Auestad, N.; Castillo, M.; Connor, W.E.; Connor, S.L.; Fitzgerald, K.; Groh-Wargo, S.; Hartmann, E.E.; et al. Growth and Development in Preterm Infants Fed Long-Chain Polyunsaturated Fatty Acids: A Prospective, Randomized Controlled Trial. Pediatrics 2001, 108, 359–371. [Google Scholar] [CrossRef] [Green Version]
- van Wezel-Meijler, G.; van der Knaap, M.S.; Huisman, J.; Jonkman, E.J.; Valk, J.; Lafeber, H.N. Dietary Supplementation of Long-Chain Polyunsaturated Fatty Acids in Preterm Infants: Effects on Cerebral Maturation. Acta Paediatr. 2002, 91, 942–950. [Google Scholar] [CrossRef] [PubMed]
- Fang, P.C.; Kuo, H.K.; Huang, C.; Ko, T.Y.; Chen, C.C.; Chung, M.Y. The Effect of Supplementation of Docosahexaenoic Acid and Arachidonic Acid on Visual Acuity and Neurodevelopment in Larger Preterm Infants. Chang Gung Med. J. 2005, 28, 708–715. [Google Scholar]
- Fewtrell, M.S.; Morley, R.; Abbott, R.A.; Singhal, A.; Isaacs, E.B.; Stephenson, T.; MacFadyen, U.; Lucas, A. Double-Blind, Randomized Trial of Long-Chain Polyunsaturated Fatty Acid Supplementation in Formula Fed to Preterm Infants. Pediatrics 2002, 110, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Fewtrell, M.S.; Abbott, R.A.; Kennedy, K.; Singhal, A.; Morley, R.; Caine, E.; Jamieson, C.; Cockburn, F.; Lucas, A. Randomized, Double-Blind Trial of Long-Chain Polyunsaturated Fatty Acid Supplementation with Fish Oil and Borage Oil in Preterm Infants. J. Pediatr. 2004, 144, 471–479. [Google Scholar] [CrossRef]
- Isaacs, E.B.; Ross, S.; Kennedy, K.; Weaver, L.T.; Lucas, A.; Fewtrell, M.S. 10-Year Cognition in Preterms after Random Assignment To Fatty Acid Supplementation in Infancy. Pediatrics 2011, 128, e890–e898. [Google Scholar] [CrossRef] [PubMed]
- Carlson, S.E.; Werkman, S.H. A Randomized Trial of Visual Attention of Preterm Infants Fed Docosahexaenoic Acid until Two Months. Lipids 1996, 31, 91–97. [Google Scholar] [CrossRef]
- Werkman, S.H.; Carlson, S.E. A Randomized Trial of Visual Attention of Preterm Infants Fed Docosahexaenoic Acid until Nine Months. Lipids 1996, 31, 91–97. [Google Scholar] [CrossRef]
- Andrew, M.J.; Parr, J.R.; Montague-Johnson, C.; Laler, K.; Holmes, J.; Baker, B.; Sullivan, P.B. Neurodevelopmental Outcome of Nutritional Intervention in Newborn Infants at Risk of Neurodevelopmental Impairment: The Dolphin Neonatal Double-Blind Randomized Controlled Trial. Dev. Med. Child Neurol. 2018, 60, 897–905. [Google Scholar] [CrossRef]
- Keim, S.A.; Gracious, B.; Boone, K.M.; Klebanoff, M.A.; Rogers, L.K.; Rausch, J.; Coury, D.L.; Sheppard, K.W.; Husk, J.; Rhoda, D.A. Ω-3 and Ω-6 Fatty Acid Supplementation May Reduce Autism Symptoms Based on Parent Report in Preterm Toddlers. J. Nutr. 2018, 148, 227–235. [Google Scholar] [CrossRef]
- Wang, Q.; Cui, Q.; Yan, C. The Effect of Supplementation of Long-Chain Polyunsaturated Fatty Acids during Lactation on Neurodevelopmental Outcomes of Preterm Infant from Infancy to School Age: A Systematic Review and Meta-Analysis. Pediatr. Neurol. 2016, 59, 54–61.e1. [Google Scholar] [CrossRef]
- Shulkin, M.; Pimpin, L.; Bellinger, D.; Kranz, S.; Fawzi, W.; Duggan, C.; Mozaffarian, D. N-3 Fatty Acid Supplementation in Mothers, Preterm Infants, and Term Infants and Childhood Psychomotor and Visual Development: A Systematic Review and Meta-Analysis. J. Nutr. 2018, 148, 409–418. [Google Scholar] [CrossRef]
- Lapillonne, A.; Moltu, S.J. Long-Chain Polyunsaturated Fatty Acids and Clinical Outcomes of Preterm Infants. Ann. Nutr. Metab. 2016, 69, 36–44. [Google Scholar] [CrossRef]
- Klevebro, S.; Juul, S.E.; Wood, T.R. A More Comprehensive Approach to the Neuroprotective Potential of Long-Chain Polyunsaturated Fatty Acids in Preterm Infants Is Needed—Should We Consider Maternal Diet and the n-6:N-3 Fatty Acid Ratio? Front. Pediatr. 2020, 7, 533. [Google Scholar] [CrossRef]
- Koletzko, B.; Bergmann, K.; Brenna, J.T.; Calder, P.C.; Campoy, C.; Clandinin, M.T.; Colombo, J.; Daly, M.; Decsi, T.; Demmelmair, H.; et al. Should Formula for Infants Provide Arachidonic Acid along with DHA? A Position Paper of the European Academy of Paediatrics and the Child Health Foundation. Am. J. Clin. Nutr. 2019, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Crawford, M.A.; Golfetto, I.; Ghebremeskel, K.; Min, Y.; Moodley, T.; Poston, L.; Phylactos, A.; Cunnane, S.; Schmidt, W. The Potential Role for Arachidonic and Docosahexaenoic Acids in Protection against Some Central Nervous System Injuries in Preterm Infants. Lipids 2003, 38, 303–315. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, A.T.; Anthony, J.C.; Diersen-Schade, D.A.; Rumsey, S.C.; Lawrence, P.; Li, C.; Nathanielsz, P.W.; Brenna, J.T. The Influence of Moderate and High Dietary Long Chain Polyunsaturated Fatty Acids (LCPUFA) on Baboon Neonate Tissue Fatty Acids. Pediatr. Res. 2007, 61, 537–545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Birch, E.E.; Carlson, S.E.; Hoffman, D.R.; Fitzgerald-Gustafson, K.M.; Fu, V.L.N.; Drover, J.R.; Castañeda, Y.S.; Minns, L.; Wheaton, D.K.H.; Mundy, D.; et al. The DIAMOND (DHA Intake and Measurement of Neural Development) Study: A Double-Masked, Randomized Controlled Clinical Trial of the Maturation of Infant Visual Acuity as a Function of the Dietary Level of Docosahexaenoic Acid. Am. J. Clin. Nutr. 2010, 91, 848–859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drover, J.R.; Hoffman, D.R.; Castañeda, Y.S.; Morale, S.E.; Garfield, S.; Wheaton, D.H.; Birch, E.E. Cognitive Function in 18-Month-Old Term Infants of the DIAMOND Study: A Randomized, Controlled Clinical Trial with Multiple Dietary Levels of Docosahexaenoic Acid. Early Hum. Dev. 2011, 87, 223–230. [Google Scholar] [CrossRef]
- Martinez, M. Tissue Levels of Polyunsaturated Fatty Acids during Early Human Development. J. Pediatr. 1992, 120, S129–S138. [Google Scholar] [CrossRef]
- Rogers, L.K.; Valentine, C.J.; Pennell, M.; Velten, M.; Britt, R.D.; Dingess, K.; Zhao, X.; Welty, S.E.; Tipple, T.E. Maternal Docosahexaenoic Acid Supplementation Decreases Lung Inflammation in Hyperoxia-Exposed Newborn Mice. J. Nutr. 2011, 141, 214–222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crawford, M.A.; Costeloe, K.; Ghebremeskel, K.; Phylactos, A.; Skirvin, L.; Stacey, F. Are Deficits of Arachidonic and Docosahexaenoic Acids Responsible for the Neural and Vascular Complications of Preterm Babies? Am. J. Clin. Nutr. 1997, 66, 1032S–1041S. [Google Scholar] [CrossRef] [Green Version]
- Ramsden, C.E.; Hennebelle, M.; Schuster, S.; Keyes, G.S.; Johnson, C.D.; Kirpich, I.A.; Dahlen, J.E.; Horowitz, M.S.; Zamora, D.; Feldstein, A.E.; et al. Effects of Diets Enriched in Linoleic Acid and Its Peroxidation Products on Brain Fatty Acids, Oxylipins, and Aldehydes in Mice. Biochim. Et Biophys. Acta-Mol. Cell Biol. Lipids 2018, 1863, 1206–1213. [Google Scholar] [CrossRef] [Green Version]
- Böckmann, K.A.; von Stumpff, A.; Bernhard, W.; Shunova, A.; Minarski, M.; Frische, B.; Warmann, S.; Schleicher, E.; Poets, C.F.; Franz, A.R. Fatty Acid Composition of Adipose Tissue at Term Indicates Deficiency of Arachidonic and Docosahexaenoic Acid and Excessive Linoleic Acid Supply in Preterm Infants. Eur. J. Nutr. 2020, 60, 861–872. [Google Scholar] [CrossRef] [PubMed]
- Hrboticky, N.; MacKinnon, M.J.; Innis, S.M. Effect of a Vegetable Oil Formula Rich in Linoleic Acid on Tissue Fatty Acid Accretion in the Brain, Liver, Plasma, and Erythrocytes of Infant Piglets. Am. J. Clin. Nutr. 1990, 51, 173–182. [Google Scholar] [CrossRef] [Green Version]
- Hrboticky, N.; MacKinnon, M.J. Retina Fatty Acid Composition of Piglets Fed from Birth with a Linoleic Acid-Rich Vegetable-Oil Formula for Infants. Am. J. Clin. Nutr. 1991, 53, 483–490. [Google Scholar] [CrossRef]
- Martinez, M.; Ballabriga, A. Effects of Parenteral Nutrition with High Doses of Linoleate on the Developing Human Liver and Brain. Lipids 1987, 22, 133–138. [Google Scholar] [CrossRef]
- Colombo, J.; Carlson, S.E.; Cheatham, C.L.; Shaddy, D.J.; Kerling, E.H.; Thodosoff, J.M.; Gustafson, K.M.; Brez, C. Long-Term Effects of LCPUFA Supplementation on Childhood Cognitive Outcomes. Am. J. Clin. Nutr. 2013, 98, 403–412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lepping, R.J.; Honea, R.A.; Martin, L.E.; Liao, K.; Choi, I.; Lee, P.; Papa, V.B.; Brooks, W.M.; Shaddy, D.J.; Carlson, S.E.; et al. Long-Chain Polyunsaturated Fatty Acid Supplementation in the First Year of Life Affects Brain Function, Structure, and Metabolism at Age Nine Years. Dev. Psychobiol. 2019, 61, 5–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luxwolda, M.F.; Kuipers, R.S.; Koops, J.H.; Muller, S.; de Graaf, D.; Dijck-Brouwer, D.A.J.; Muskiet, F.A.J. Interrelationships between Maternal DHA in Erythrocytes, Milk and Adipose Tissue. Is 1 Wt% DHA the Optimal Human Milk Content? Data from Four Tanzanian Tribes Differing in Lifetime Stable Intakes of Fish. Br. J. Nutr. 2014, 111, 854–866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jandacek, R.J. Linoleic Acid: A Nutritional Quandary. Healthcare 2017, 5, 25. [Google Scholar] [CrossRef] [Green Version]
- Raatz, S.K.; Conrad, Z.; Jahns, L. Trends in Linoleic Acid Intake in the United States Adult Population: NHANES 1999–2014. Prostaglandins Leukot. Essent. Fat. Acids 2018, 133, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Innis, S.M. Human Milk: Maternal Dietary Lipids and Infant Development. Proc. Nutr. Soc. 2007, 66, 397–404. [Google Scholar] [CrossRef] [PubMed]
- Lattka, E.; Rzehak, P.; Szabó, É.; Jakobik, V.; Weck, M.; Weyermann, M.; Grallert, H.; Rothenbacher, D.; Heinrich, J.; Brenner, H.; et al. Genetic Variants in the FADS Gene Cluster Are Associated with Arachidonic Acid Concentrations of Human Breast Milk at 1.5 and 6 Mo Postpartum and Influence the Course of Milk Dodecanoic, Tetracosenoic, and Trans-9-Octadecenoic Acid Concentrations over duration of lactation. Am. J. Clin. Nutr. 2011, 93, 382–391. [Google Scholar] [CrossRef] [Green Version]
- Steer, C.D.; Lattka, E.; Koletzko, B.; Golding, J.; Hibbeln, J.R. Maternal Fatty Acids in Pregnancy, FADS Polymorphisms, and Child Intelligence Quotient at 8 y of Age. Am. J. Clin. Nutr. 2013, 98, 1575–1582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lorenzo, I.S.; Tonato, A.M.C.; Puentes, A. de la G.; Nieto, A.; Herrmann, F.; Dieguez, E.; Castellote, A.I.; López-Sabater, M.C.; Rodríguez-Palmero, M.; Campoy, C. The Effect of an Infant Formula Supplemented with AA and DHA on Fatty Acid Levels of Infants with Different FADS Genotypes: The COGNIS Study. Nutrients 2019, 11, 602. [Google Scholar] [CrossRef] [Green Version]
- Gould, J.F.; Anderson, A.J.; Yelland, L.N.; Gibson, R.A.; Makrides, M. Maternal Characteristics Influence Response to DHA during Pregnancy. Prostaglandins Leukot. Essent. Fat. Acids 2016, 108, 5–12. [Google Scholar] [CrossRef] [PubMed]
- Ramakrishnan, U.; Stinger, A.; Digirolamo, A.M.; Martorell, R.; Neufeld, L.M.; Rivera, J.A.; Schnaas, L.; Stein, A.D.; Wang, M. Prenatal Docosahexaenoic Acid Supplementation and Offspring Development at 18 Months: Randomized Controlled Trial. PLoS ONE 2015, 10, e0120065. [Google Scholar] [CrossRef] [Green Version]
- Middleton, P.; Gomersall, J.C.; Gould, J.F.; Shepherd, E.; Olsen, S.F.; Makrides, M. Omega-3 Fatty Acid Addition during Pregnancy. In Cochrane Database of Systematic Reviews; John Wiley and Sons Ltd.: Hoboken, NJ, USA, 2018; p. CD003402. [Google Scholar] [CrossRef]
- Kim, H.; Kim, H.; Lee, E.; Kim, Y.; Ha, E.H.; Chang, N. Association between Maternal Intake of N-6 to n-3 Fatty Acid Ratio during Pregnancy and Infant Neurodevelopment at 6 Months of Age: Results of the MOCEH Cohort Study. Nutr. J. 2017, 16, 23. [Google Scholar] [CrossRef] [Green Version]
- Bernard, J.Y.; de Agostini, M.; Forhan, A.; de Lauzon-Guillain, B.; Charles, M.A.; Heude, B.; Annesi-Maesano, I.; Botton, J.; Charles, M.A.; Dargent-Molina, P.; et al. The Dietary N6:N3 Fatty Acid Ratio during Pregnancy Is Inversely Associated with Child Neurodevelopment in the EDEN Mother-Child Cohort. J. Nutr. 2013, 143, 1481–1488. [Google Scholar] [CrossRef] [Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Heath, R.J.; Klevebro, S.; Wood, T.R. Maternal and Neonatal Polyunsaturated Fatty Acid Intake and Risk of Neurodevelopmental Impairment in Premature Infants. Int. J. Mol. Sci. 2022, 23, 700. https://doi.org/10.3390/ijms23020700
Heath RJ, Klevebro S, Wood TR. Maternal and Neonatal Polyunsaturated Fatty Acid Intake and Risk of Neurodevelopmental Impairment in Premature Infants. International Journal of Molecular Sciences. 2022; 23(2):700. https://doi.org/10.3390/ijms23020700
Chicago/Turabian StyleHeath, Rory J., Susanna Klevebro, and Thomas R. Wood. 2022. "Maternal and Neonatal Polyunsaturated Fatty Acid Intake and Risk of Neurodevelopmental Impairment in Premature Infants" International Journal of Molecular Sciences 23, no. 2: 700. https://doi.org/10.3390/ijms23020700