
Reversing the Psychiatric Effects of Neurodevelopmental Cannabinoid Exposure: Exploring Pharmacotherapeutic Interventions for Symptom Improvement

Abstract
:1. Introduction
2. Long-Lasting Consequences of Sustained Cannabis Use during Adolescence: Clinical Evidence
3. Effects of THC Exposure during Adolescence: Insight from Neurodevelopmental Animal Models
3.1. Risks for Schizophrenia and Cognitive Impairments
3.2. Risks for Depressive Like-Phenotype and Long-Term Anxiety
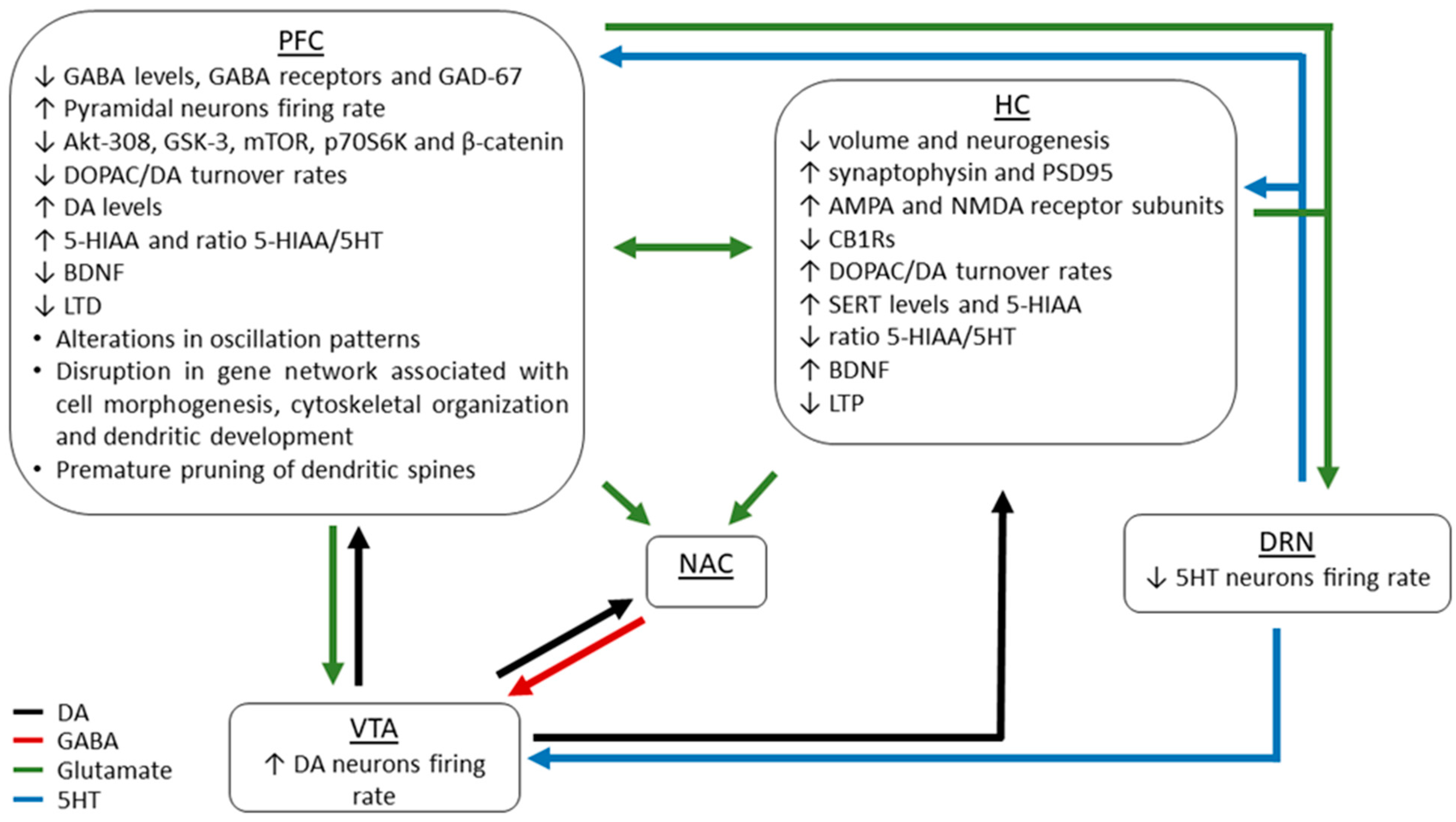
4. Neurobiological Mechanisms Underlying THC-Related Detrimental Consequences
5. Therapeutic Perspective: Potential Intervention for Mitigate the Neuropsychiatric Risks
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dhein, S. Different Effects of Cannabis Abuse on Adolescent and Adult Brain. Pharmacology 2020. [Google Scholar] [CrossRef] [PubMed]
- Hammond, C.J.; Chaney, A.; Hendrickson, B.; Sharma, P. Cannabis use among U.S. adolescents in the era of marijuana legalization: A review of changing use patterns, comorbidity, and health correlates. Int. Rev. Psychiatry 2020, 32, 221–234. [Google Scholar] [CrossRef] [PubMed]
- Canadian Cannabis Survey 2020: Summary. Available online: https://www.canada.ca/en/health-canada/services/drugs-medication/cannabis/research-data/canadian-cannabis-survey-2020-summary.html (accessed on 21 December 2020).
- Maccarrone, M. Missing Pieces to the Endocannabinoid Puzzle. Trends Mol. Med. 2020, 26, 263–272. [Google Scholar] [CrossRef] [PubMed]
- Tirado-Muñoz, J.; Lopez-Rodriguez, A.B.; Fonseca, F.; Farré, M.; Torrens, M.; Viveros, M.P. Effects of cannabis exposure in the prenatal and adolescent periods: Preclinical and clinical studies in both sexes. Front. Neuroendocrinol. 2020, 57, 100841. [Google Scholar] [CrossRef] [PubMed]
- Thorpe, H.H.A.; Hamidullah, S.; Jenkins, B.W.; Khokhar, J.Y. Adolescent neurodevelopment and substance use: Receptor expression and behavioral consequences. Pharmacol. Ther. 2020, 206, 107431. [Google Scholar] [CrossRef] [PubMed]
- Fischer, A.S.; Tapert, S.F.; Louie, D.L.; Schatzberg, A.F.; Singh, M.K. Cannabis and the Developing Adolescent Brain. Curr. Treat. Options Psychiatry 2020, 7, 144–161. [Google Scholar] [CrossRef] [PubMed]
- Stuyt, E. The Problem with the Current High Potency THC Marijuana from the Perspective of an Addiction Psychiatrist. Mo. Med. 2018, 115, 482–486. [Google Scholar]
- Freeman, T.P.; Craft, S.; Wilson, J.; Stylianou, S.; ElSohly, M.; Di Forti, M.; Lynskey, M.T. Changes in delta-9-tetrahydrocannabinol (THC) and cannabidiol (CBD) concentrations in cannabis over time: Systematic review and meta-analysis. Addiction 2020, 1000–1010. [Google Scholar] [CrossRef] [PubMed]
- Freeman, A.M.; Petrilli, K.; Lees, R.; Hindocha, C.; Mokrysz, C.; Curran, H.V.; Saunders, R.; Freeman, T.P. How does cannabidiol (CBD) influence the acute effects of delta-9-tetrahydrocannabinol (THC) in humans? A systematic review. Neurosci. Biobehav. Rev. 2019, 107, 586–712. [Google Scholar] [CrossRef]
- Lorenzetti, V.; Solowij, N.; Yücel, M. The role of cannabinoids in neuroanatomic alterations in cannabis users. Biol. Psychiatry 2016, 79, e17–e31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solowij, N.; Broyd, S.; Greenwood, L.M.; van Hell, H.; Martelozzo, D.; Rueb, K.; Todd, J.; Liu, Z.; Galettis, P.; Martin, J.; et al. A randomised controlled trial of vaporised Δ9-tetrahydrocannabinol and cannabidiol alone and in combination in frequent and infrequent cannabis users: Acute intoxication effects. Eur. Arch. Psychiatry Clin. Neurosci. 2019, 269, 17–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hines, L.A.; Freeman, T.P.; Gage, S.H.; Zammit, S.; Hickman, M.; Cannon, M.; Munafo, M.; MacLeod, J.; Heron, J. Association of High-Potency Cannabis Use with Mental Health and Substance Use in Adolescence. JAMA Psychiatry 2020, 77, 1044–1051. [Google Scholar] [CrossRef] [PubMed]
- Di Forti, M.; Marconi, A.; Carra, E.; Fraietta, S.; Trotta, A.; Bonomo, M.; Bianconi, F.; Gardner-Sood, P.; O’Connor, J.; Russo, M.; et al. Proportion of patients in south London with first-episode psychosis attributable to use of high potency cannabis: A case-control study. Lancet Psychiatry 2015, 2, 233–238. [Google Scholar] [CrossRef] [Green Version]
- Di Forti, M.; Quattrone, D.; Freeman, T.P.; Tripoli, G.; Gayer-Anderson, C.; Quigley, H.; Rodriguez, V.; Jongsma, H.E.; Ferraro, L.; La Cascia, C.; et al. The contribution of cannabis use to variation in the incidence of psychotic disorder across Europe (EU-GEI): A multicentre case-control study. Lancet Psychiatry 2019, 6, 427–436. [Google Scholar] [CrossRef] [Green Version]
- Schoeler, T.; Petros, N.; Di Forti, M.; Klamerus, E.; Foglia, E.; Ajnakina, O.; Gayer-Anderson, C.; Colizzi, M.; Quattrone, D.; Behlke, I.; et al. Effects of continuation, frequency, and type of cannabis use on relapse in the first 2 years after onset of psychosis: An observational study. Lancet Psychiatry 2016, 3, 947–953. [Google Scholar] [CrossRef] [Green Version]
- Andréasson, S.; Allebeck, P.; Engström, A.; Rydberg, U. Cannabis and schizophrenia. A longitudinal study of Swedish conscripts. Lancet 1987, 2, 1483–1486. [Google Scholar] [CrossRef]
- Stefanis, N.C.; Delespaul, P.; Henquet, C.; Bakoula, C.; Stefanis, C.N.; Van Os, J. Early adolescent cannabis exposure and positive and negative dimensions of psychosis. Addiction 2004, 99, 1333–1341. [Google Scholar] [CrossRef] [PubMed]
- Gobbi, G.; Atkin, T.; Zytynski, T.; Wang, S.; Askari, S.; Boruff, J.; Ware, M.; Marmorstein, N.; Cipriani, A.; Dendukuri, N.; et al. Association of Cannabis Use in Adolescence and Risk of Depression, Anxiety, and Suicidality in Young Adulthood: A Systematic Review and Meta-analysis. JAMA Psychiatry 2019, 76, 426–434. [Google Scholar] [CrossRef] [PubMed]
- Leadbeater, B.J.; Ames, M.E.; Linden-Carmichael, A.N. Age-varying effects of cannabis use frequency and disorder on symptoms of psychosis, depression and anxiety in adolescents and adults. Addiction 2019, 114, 278–293. [Google Scholar] [CrossRef] [Green Version]
- Harvey, P.D. Smoking cannabis and acquired impairments in cognition: Starting early seems like a really bad idea. Am. J. Psychiatry 2019, 176, 90–91. [Google Scholar] [CrossRef]
- Renard, J.; Krebs, M.; Pen, G.L.; Jay, T.M. Long-term consequences of adolescent cannabinoid exposure in adult psychopathology. Front. Neurosci. 2014, 8, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meier, M.H.; Caspi, A.; Ambler, A.; Harrington, H.L.; Houts, R.; Keefe, R.S.E.; McDonald, K.; Ward, A.; Poulton, R.; Moffitt, T.E. Persistent cannabis users show neuropsychological decline from childhood to midlife. Proc. Natl. Acad. Sci. USA 2012, 109, 2657–2664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morin, J.F.G.; Afzali, M.H.; Bourque, J.; Stewart, S.H.; Séguin, J.R.; O’Leary-Barrett, M.; Conrod, P.J. A population-based analysis of the relationship between substance use and adolescent cognitive development. Am. J. Psychiatry 2019, 176, 98–106. [Google Scholar] [CrossRef] [PubMed]
- Jacobus, J.; Courtney, K.E.; Hodgdon, E.A.; Baca, R. Cannabis and the developing brain: What does the evidence say? Birth Defects Res. 2019, 111, 1302–1307. [Google Scholar] [CrossRef] [PubMed]
- Gruber, S.A.; Sagar, K.A. Marijuana on the Mind? The Impact of Marijuana on Cognition, Brain Structure, and Brain Function, and Related Public Policy Implications. Policy Insights Behav. Brain Sci. 2017, 4, 104–111. [Google Scholar] [CrossRef]
- Filbey, F.M.; McQueeny, T.; DeWitt, S.J.; Mishra, V. Preliminary findings demonstrating latent effects of early adolescent marijuana use onset on cortical architecture. Dev. Cogn. Neurosci. 2015, 16, 16–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez-Larson, M.P.; Bogorodzki, P.; Rogowska, J.; McGlade, E.; King, J.B.; Terry, J.; Todd, D.Y.; Yurgelun-Todd, D. Altered Prefrontal and Insular Cortical Thickness. Behav. Brain Res. 2011, 220, 164–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, W.; Mathew, R.; Turkington, T.; Hawk, T.; Coleman, R.E.; Provenzale, J. Brain morphological changes and early marijuana use: A magnetic resonance and positron emission tomography study. J. Addict. Dis. 2000, 19, 1–22. [Google Scholar] [CrossRef]
- Schoeler, T.; Bhattacharyya, S. The effect of cannabis use on memory function: An update. Subst. Abuse Rehabil. 2013, 11–27. [Google Scholar] [CrossRef] [Green Version]
- Sachs, J.; McGlade, E.; Yurgelun-Todd, D. Safety and Toxicology of Cannabinoids. Neurotherapeutics 2015, 12, 735–746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCormick, M.A.; Shekhar, A. Review of Marijuana Use in the Adolescent Population and Implications of its Legalization in the United States. J. Drug Metab. Toxicol. 2014, 5, 7–10. [Google Scholar] [CrossRef] [Green Version]
- Yücel, M.; Solowij, N.; Respondek, C.; Whittle, S.; Fornito, A.; Pantelis, C.; Lubman, D.I. Regional brain abnormalities associated with long-term heavy cannabis use. Arch. Gen. Psychiatry 2008, 65, 694–701. [Google Scholar] [CrossRef] [PubMed]
- Zalesky, A.; Solowij, N.; Yücel, M.; Lubman, D.I.; Takagi, M.; Harding, I.H.; Lorenzetti, V.; Wang, R.; Searle, K.; Pantelis, C.; et al. Effect of long-term cannabis use on axonal fibre connectivity. Brain 2012, 135, 2245–2255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mato, S.; Del Olmo, E.; Pazos, A. Ontogenetic development of cannabinoid receptor expression and signal transduction functionality in the human brain. Eur. J. Neurosci. 2003, 17, 1747–1754. [Google Scholar] [CrossRef] [PubMed]
- Prescot, A.P.; Renshaw, P.F.; Yurgelun-Todd, D.A. γ-Amino butyric acid and glutamate abnormalities in adolescent chronic marijuana smokers. Drug Alcohol Depend. 2013, 129, 232–239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rigucci, S.; Xin, L.; Klauser, P.; Baumann, P.S.; Alameda, L.; Cleusix, M.; Jenni, R.; Ferrari, C.; Pompili, M.; Gruetter, R.; et al. Cannabis use in early psychosis is associated with reduced glutamate levels in the prefrontal cortex. Psychopharmacology 2018, 235, 13–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashimoto, T.; Volk, D.W.; Eggan, S.M.; Mirnics, K.; Pierri, J.N.; Sun, Z.; Sampson, A.R.; Lewis, D.A. Gene expression deficits in a subclass of GABA neurons in the prefrontal cortex of subjects with schizophrenia. J. Neurosci. 2003, 23, 6315–6326. [Google Scholar] [CrossRef] [PubMed]
- Volk, D.W.; Austin, M.C.; Pierri, J.N.; Sampson, A.R.; Lewis, D.A. GABA transporter-1 mRNA in the prefrontal cortex in schizophrenia: Decreased expression in a subset of neurons. Am. J. Psychiatry 2001, 158, 256–265. [Google Scholar] [CrossRef]
- Maldonado-Aviles, J.G.; Curley, A.A.; Hashimoto, T.; Morrow, L.; Ramsey, A.J.; O’Donnell, P.; Volk, D.W.; Lewis, D.A. Altered Markers of Tonic Inhibition in the Dorsolateral Prefrontal Cortex of Subjects with Schizophrenia. Am. J. Psychiatry 2009, 166, 450–459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Jonge, J.C.; Vinkers, C.H.; Hulshoff Pol, H.E.; Marsman, A. GABAergic mechanisms in schizophrenia: Linking postmortem and In vivo studies. Front. Psychiatry 2017, 8, 118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karolewicz, B.; Maciag, D.; O’Dwyer, G.; Stockmeier, C.A.; Feyissa, A.M.; Rajkowka, G. Reduced Level of Glutamic Acid Decarboxylase-67 kDa in the Prefrontal Cortex in Major Depression. Int. J. Neuropsychopharmacol. 2010, 13, 411–420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luscher, B.; Shen, Q.; Sahir, N. The GABAergic deficit hypothesis of major depressive disorder. Mol. Psychiatry 2011, 16, 383–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fogaça, M.V.; Duman, R.S. Cortical GABAergic dysfunction in stress and depression: New insights for therapeutic interventions. Front. Cell. Neurosci. 2019, 13, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Bloomfield, M.A.P.; Morgan, C.J.A.; Egerton, A.; Kapur, S.; Curran, H.V.; Howes, O.D. Dopaminergic function in cannabis users and its relationship to cannabis-induced psychotic symptoms. Biol. Psychiatry 2014, 75, 470–478. [Google Scholar] [CrossRef] [PubMed]
- Urban, N.B.; Slifstein, M.; Thompson, J.L.; Xu, X.; Girgis, R.R.; Raheja, S.; Haney, M.; Abi-dargham, A. Dopamine release in chronic cannabis users: A [11C]raclopride Positron Emission Tomography study. Biol. Psychiatry 2012, 71, 677–683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caspi, A.; Moffitt, T.E.; Cannon, M.; McClay, J.; Murray, R.; Harrington, H.L.; Taylor, A.; Arseneault, L.; Williams, B.; Braithwaite, A.; et al. Moderation of the effect of adolescent-onset cannabis use on adult psychosis by a functional polymorphism in the catechol-O-methyltransferase gene: Longitudinal evidence of a gene X environment interaction. Biol. Psychiatry 2005, 57, 1117–1127. [Google Scholar] [CrossRef] [PubMed]
- Morgan, C.J.A.; Freeman, T.P.; Powell, J.; Curran, H.V. AKT1 genotype moderates the acute psychotomimetic effects of naturalistically smoked cannabis in young cannabis smokers. Transl. Psychiatry 2016, 6, e738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Forti, M.; Iyegbe, C.; Sallis, H.; Kolliakou, A.; Falcone, M.A.; Paparelli, A.; Sirianni, M.; La Cascia, C.; Stilo, S.A.; Marques, T.R.; et al. Confirmation that the AKT1 (rs2494732) genotype influences the risk of psychosis in cannabis users. Biol. Psychiatry 2012, 72, 811–816. [Google Scholar] [CrossRef] [PubMed]
- Miguez, M.J.; Chan, W.; Espinoza, L.; Tarter, R.; Perez, C. Marijuana use among adolescents is associated with deleterious alterations in mature BDNF. AIMS Public Health 2019, 6, 4–14. [Google Scholar] [CrossRef] [PubMed]
- Mané, A.; Bergé, D.; Penzol, M.J.; Parellada, M.; Bioque, M.; Lobo, A.; González-Pinto, A.; Corripio, I.; Cabrera, B.; Sánchez-Torres, A.M.; et al. Cannabis use, COMT, BDNF and age at first-episode psychosis. Psychiatry Res. 2017, 250, 38–43. [Google Scholar] [CrossRef] [PubMed]
- Tirelli, E.; Laviola, G.; Adriani, W. Ontogenesis of behavioral sensitization and conditioned place preference induced by psychostimulants in laboratory rodents. Neurosci. Biobehav. Rev. 2003, 27, 163–178. [Google Scholar] [CrossRef]
- Braff, D.L.; Geyer, M.A. Sensorimotor Gating and Schizophrenia. Human and animal model studies. Arch. Gen. Psychiatry 1990, 47, 181–188. [Google Scholar] [CrossRef] [PubMed]
- Renard, J.; Rosen, L.G.; Loureiro, M.; De Oliveira, C.; Schmid, S.; Rushlow, W.J.; Laviolette, S.R. Adolescent Cannabinoid Exposure Induces a Persistent Sub-Cortical Hyper-Dopaminergic State and Associated Molecular Adaptations in the Prefrontal Cortex. Cereb. Cortex 2017, 27, 1297–1310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abela, A.R.; Rahbarnia, A.; Wood, S.; Lê, A.D.; Fletcher, P.J. Adolescent exposure to Δ9-tetrahydrocannabinol delays acquisition of paired-associates learning in adulthood. Psychopharmacology 2019, 236, 1875–1886. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Nuechterlein, K.H.; Knowlton, B.J.; Bearden, C.E.; Cannon, T.D.; Fiske, A.P.; Ghermezi, L.; Hayata, J.N.; Hellemann, G.S.; Horan, W.P.; et al. Episodic Memory for Dynamic Social Interaction Across Phase of Illness in Schizophrenia. Schizophr. Bull. 2018, 44, 620–630. [Google Scholar] [CrossRef] [PubMed]
- Rubino, T.; Parolaro, D. The impact of exposure to cannabinoids in adolescence: Insights from animal models. Biol. Psychiatry 2016, 79, 578–585. [Google Scholar] [CrossRef] [PubMed]
- Renard, J.; Rushlow, W.J.; Laviolette, S.R. What can rats tell us about adolescent cannabis exposure? Insights from. Can. J. Psychiatry 2016, 61, 328–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Renard, J.; Szkudlarek, H.J.; Kramar, C.P.; Jobson, C.E.L.; Moura, K.; Rushlow, W.J.; Laviolette, S.R. Adolescent THC Exposure Causes Enduring Prefrontal Cortical Disruption of GABAergic Inhibition and Dysregulation of Sub-Cortical Dopamine Function. Sci. Rep. 2017, 7, 11420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poulia, N.; Delis, F.; Brakatselos, C.; Lekkas, P.; Kokras, N.; Dalla, C.; Antoniou, K. Escalating low-dose Δ9-tetrahydrocannabinol exposure during adolescence induces differential behavioral and neurochemical effects in male and female adult rats. Eur. J. Neurosci. 2020, 52, 2681–2693. [Google Scholar] [CrossRef] [PubMed]
- Rubino, T.; Parolaro, D. Sex-dependent vulnerability to Cannabis abuse in adolescence. Front. Psychiatry 2015, 6, 1–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poulia, N.; Delis, F.; Brakatselos, C.; Polissidis, A.; Koutmani, Y.; Kokras, N.; Dalla, C.; Politis, P.K.; Antoniou, K. Detrimental effects of adolescent escalating low-dose Δ9-tetrahydrocannabinol leads to a specific bio-behavioural profile in adult male rats. Br. J. Pharmacol. 2021, 178, 1722–1736. [Google Scholar] [CrossRef]
- Chen, H.T.; Mackie, K. Adolescent Δ9-Tetrahydrocannabinol Exposure Selectively Impairs Working Memory but Not Several Other mPFC-Mediated Behaviors. Front. Psychiatry 2020, 11, 576214. [Google Scholar] [CrossRef] [PubMed]
- Kasten, C.R.; Zhang, Y.; Boehm, S.L. Acute cannabinoids produce robust anxiety-like and locomotor effects in mice, but long-term consequences are age- and sex-dependent. Front. Behav. Neurosci. 2019, 13, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kasten, C.R.; Zhang, Y.; Boehm, S.L. Acute and Long-Term Effects of Δ9-tetrahydrocannabinol on Object Recognition and Anxiety-Like Activity are Age- and Strain-Dependent in Mice. Pharmacol. Biochem. Behav. 2018, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Nelson, N.G.; Law, W.X.; Weingarten, M.J.; Carnevale, L.N.; Das, A.; Liang, N.C. Combined ∆9-tetrahydrocannabinol and moderate alcohol administration: Effects on ingestive behaviors in adolescent male rats. Psychopharmacology 2019, 236, 671–684. [Google Scholar] [CrossRef] [PubMed]
- Gibula-Tarlowska, E.; Wydra, K.; Kotlinska, J.H. Deleterious effects of ethanol, ∆(9)-tetrahydrocannabinol (thc), and their combination on the spatial memory and cognitive flexibility in adolescent and adult male rats in the barnes maze task. Pharmaceutics 2020, 12, 654. [Google Scholar] [CrossRef] [PubMed]
- De Gregorio, D.; Dean Conway, J.; Canul, M.-L.; Posa, L.; Bambico, F.R.; Gobbi, G. Effects of Chronic Exposure to Low-Dose delta-9-Tetrahydrocannabinol in Adolescence and Adulthood on Serotonin/Norepinephrine Neurotransmission and Emotional Behavior. Int. J. Neuropsychopharmacol. 2020, 751–761. [Google Scholar] [CrossRef] [PubMed]
- Prini, P.; Rusconi, F.; Zamberletti, E.; Gabaglio, M.; Penna, F.; Fasano, M.; Battaglioli, E.; Parolaro, D.; Rubino, T. Adolescent THC exposure in female rats leads to cognitive deficits through a mechanism involving chromatin modifications in the prefrontal cortex. J. Psychiatry Neurosci. 2017, 43, 170082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Realini, N.; Vigano’, D.; Guidali, C.; Zamberletti, E.; Rubino, T.; Parolaro, D. Chronic URB597 treatment at adulthood reverted most depressive-like symptoms induced by adolescent exposure to THC in female rats. Neuropharmacology 2011, 60, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Rubino, T.; Vigano, D.; Realini, N.; Guidali, C.; Braida, D.; Capurro, V.; Castiglioni, C.; Cherubino, F.; Romualdi, P.; Candeletti, S.; et al. Chronic delta-9-Tetrahydrocannabinol During Adolescence Provokes Sex-Dependent Changes in the Emotional Profile in Adult Rats: Behavioral and Biochemical Correlates. Neuropsychopharmacology 2008, 33, 2760–2771. [Google Scholar] [CrossRef] [PubMed]
- Saravia, R.; Ten-Blanco, M.; Julià-Hernández, M.; Gagliano, H.; Andero, R.; Armario, A.; Maldonado, R.; Berrendero, F. Concomitant THC and stress adolescent exposure induces impaired fear extinction and related neurobiological changes in adulthood. Neuropharmacology 2019, 144, 345–357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cadoni, C.; Simola, N.; Espa, E.; Fenu, S.; Di Chiara, G. Strain dependence of adolescent Cannabis influence on heroin reward and mesolimbic dopamine transmission in adult Lewis and Fischer 344 rats. Addict. Biol. 2015, 20, 132–142. [Google Scholar] [CrossRef] [PubMed]
- Bruijnzeel, A.W.; Knight, P.; Panunzio, S.; Xue, S.; Bruner, M.M.; Wall, S.C.; Pompilus, M.; Febo, M.; Setlow, B. Effects in rats of adolescent exposure to cannabis smoke or THC on emotional behavior and cognitive function in adulthood. Psychopharmacology 2019, 236, 2773–2784. [Google Scholar] [CrossRef] [PubMed]
- Llorente-berzal, A.; Puighermanal, E.; Burokas, A. Sex-Dependent Psychoneuroendocrine Effects of THC and MDMA in an Animal Model of Adolescent Drug Consumption. PLoS ONE 2013, 8. [Google Scholar] [CrossRef]
- Stringfield, S.J.; Torregrossa, M.M. Intravenous self-administration of delta-9-THC in adolescent rats produces long-lasting alterations in behavior and receptor protein expression. Psychopharmacology 2021, 238, 305–319. [Google Scholar] [CrossRef] [PubMed]
- Hirvonen, J.; Goodwind, R.; Li, C.-T.; Terry, G.; Zoghbi, S.; Morse, C.; Pike, V.; Volkow, N.; Huestis, M.; Innis, R. Reversible and regionally selective downregulation of brain cannabinoid CB1 receptors in chronic daily cannabis smokers. Mol. Psychiatry 2012, 17, 642–649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burston, J.J.; Wiley, J.L.; Craig, A.A.; Selley, D.E.; Sim-Selley, L.J. Regional enhancement of cannabinoid CB 1 receptor desensitization in female adolescent rats following repeated Δ9-tetrahydrocannabinol exposure. Br. J. Pharmacol. 2010, 161, 103–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, M.L.; Chadwick, B.; Dickstein, D.L.; Purushothaman, I.; Egervari, G.; Rahman, T.; Tessereau, C.; Hof, P.R.; Roussos, P.; Shen, L.; et al. Adolescent exposure to Δ9-tetrahydrocannabinol alters the transcriptional trajectory and dendritic architecture of prefrontal pyramidal neurons. Mol. Psychiatry 2019, 24, 588–600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zamberletti, E.; Beggiato, S.; Steardo, L.; Prini, P.; Antonelli, T.; Ferraro, L.; Rubino, T.; Parolaro, D. Neurobiology of Disease Alterations of prefrontal cortex GABAergic transmission in the complex psychotic-like phenotype induced by adolescent delta-9-tetrahydrocannabinol exposure in rats. Neurobiol. Dis. 2014, 63, 35–47. [Google Scholar] [CrossRef] [PubMed]
- Renard, J.; Rushlow, W.J.; Laviolette, S.R. Effects of adolescent THC exposure on the prefrontal GABAergic system: Implications for schizophrenia-related psychopathology. Front. Psychiatry 2018, 9, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Swerdlow, N.R.; Geyer, M.A.; Braff, D.L. Neural circuit regulation of prepulse inhibition of startle in the rat: Current knowledge and future challenges. Psychopharmacology 2001, 156, 194–215. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.H.; Chan, P.Y.S.; Niddam, D.M.; Tsai, S.Y.; Hsu, S.C.; Liu, C.Y. Sensory gating, inhibition control and gamma oscillations in the human somatosensory cortex. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef] [PubMed]
- Draycott, B.; Loureiro, M.; Ahmad, T.; Tan, H.; Zunder, J.; Laviolette, S.R. Cannabinoid transmission in the prefrontal cortex Bi-phasically controls emotional memory formation via functional interactions with the ventral tegmental area. J. Neurosci. 2014, 34, 13096–13109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calakos, K.C.; Bhatt, S.; Foster, D.W.; Cosgrove, K.P. Author Correction: Underlying Sex Differences in Cannabis Use. Curr. Addict. Rep. 2017, 4, 439–453. [Google Scholar] [CrossRef] [PubMed]
- Zamberletti, E.; Gabaglio, M.; Grilli, M.; Prini, P.; Catanese, A.; Pittaluga, A.; Marchi, M.; Rubino, T.; Parolaro, D. Long-term hippocampal glutamate synapse and astrocyte dysfunctions underlying the altered phenotype induced by adolescent THC treatment in male rats. Pharmacol. Res. 2016, 111, 459–470. [Google Scholar] [CrossRef]
- Cuccurazzu, B.; Zamberletti, E.; Nazzaro, C.; Prini, P.; Trusel, M.; Grilli, M.; Parolaro, D.; Tonini, R.; Rubino, T. Adult Cellular Neuroadaptations Induced by Adolescent THC Exposure in Female Rats Are Rescued by Enhancing Anandamide Signaling. Int. J. Neuropsychopharmacol. 2018, 21, 1014–1024. [Google Scholar] [CrossRef] [Green Version]
- Bagot, R.C.; Parise, E.M.; Peña, C.J.; Zhang, H.X.; Maze, I.; Chaudhury, D.; Persaud, B.; Cachope, R.; Bolaños-Guzmán, C.A.; Cheer, J.; et al. Ventral hippocampal afferents to the nucleus accumbens regulate susceptibility to depression. Nat. Commun. 2015, 6. [Google Scholar] [CrossRef] [PubMed]
- Loureiro, M.; Kramar, C.; Renard, J.; Rosen, L.G.; Laviolette, S.R. Cannabinoid Transmission in the Hippocampus Activates Nucleus Accumbens Neurons and Modulates Reward and Aversion-Related Emotional Salience. Biol. Psychiatry 2016, 80, 216–225. [Google Scholar] [CrossRef]
- Hudson, R.; Renard, J.; Norris, C.; Rushlow, W.J.; Laviolette, S.R. Cannabidiol Counteracts the Psychotropic Side-Effects of Δ-9-Tetrahydrocannabinol in the Ventral Hippocampus through Bidirectional Control of ERK1-2 Phosphorylation. J. Neurosci. 2019, 39, 8762–8777. [Google Scholar] [CrossRef] [PubMed]
- Lewis, D.A.; Cho, R.Y.; Carter, C.S.; Eklund, K.R.; Forster, S.; Kelly, M.A.; Montrose, D. Subunit-Selective Modulation of GABA Type A Receptor Neurotransmission and Cognition in Schizophrenia. Am. J. Psychiatry 2008, 165, 1585–1593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopes Sakamoto, F.; Metzker Pereira Ribeiro, R.; Amador Bueno, A.; Oliveira Santos, H. Psychotropic effects of L-theanine and its clinical properties: From the management of anxiety and stress to a potential use in schizophrenia. Pharmacol. Res. 2019, 147, 104395. [Google Scholar] [CrossRef] [PubMed]
- Hidese, S.; Ota, M.; Wakabayashi, C.; Noda, T.; Ozawa, H.; Okubo, T.; Kunugi, H. Effects of chronic l-theanine administration in patients with major depressive disorder: An open-label study. Acta Neuropsychiatr. 2017, 29, 72–79. [Google Scholar] [CrossRef] [PubMed]
- Shen, M.; Yang, Y.; Wu, Y.; Zhang, B.; Wu, H.; Wang, L.; Tang, H.; Chen, J. L-theanine ameliorate depressive-like behavior in a chronic unpredictable mild stress rat model via modulating the monoamine levels in limbic–cortical–striatal–pallidal–thalamic-circuit related brain regions. Phyther. Res. 2019, 33, 412–421. [Google Scholar] [CrossRef] [PubMed]
- Ota, M.; Wakabayashi, C.; Sato, N.; Hori, H.; Hattori, K.; Teraishi, T.; Ozawa, H.; Okubo, T.; Kunugi, H. Effect of L-theanine on glutamatergic function in patients with schizophrenia. Acta Neuropsychiatr. 2015, 27, 291–296. [Google Scholar] [CrossRef] [PubMed]
- De Felice, M.; Renard, J.; Hudson, R.; Szkudlarek, H.J.; Pereira, B.J.; Schmid, S.; Rushlow, W.J.; Laviolette, S.R. L-Theanine Prevents Long-Term Affective and Cognitive Side Effects of Adolescent Δ-9-Tetrahydrocannabinol Exposure and Blocks Associated Molecular and Neuronal Abnormalities in the Mesocorticolimbic Circuitry. J. Neurosci. 2021, 41, 739–750. [Google Scholar] [CrossRef] [PubMed]
- Sutton, L.P.; Rushlow, W.J. The dopamine D2 receptor regulates Akt and GSK-3 via Dvl-3. Int. J. Neuropsychopharmacol. 2012, 3, 965–979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takeshima, M.; Miyazaki, I.; Murakami, S.; Kita, T.; Asanuma, M. L-Theanine protects against excess dopamine-induced neurotoxicity in the precence of astrocytes. J. Clin. Biochem. Nutr. 2016, 59, 93–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berthoux, C.; Hamieh, A.M.; Rogliardo, A.; Doucet, E.L.; Coudert, C.; Ango, F.; Grychowska, K.; Chaumont-Dubel, S.; Zajdel, P.; Maldonado, R.; et al. Early 5-HT6 receptor blockade prevents symptom onset in a model of adolescent cannabis abuse. EMBO Mol. Med. 2020, 12, 1–17. [Google Scholar] [CrossRef]
- Meffre, J.; Chaumont-Dubel, S.; Mannoury la Cour, C.; Loiseau, F.; Watson, D.J.G.; Dekeyne, A.; Séveno, M.; Rivet, J.M.; Gaven, F.; Déléris, P.; et al. 5-HT6 receptor recruitment of mTOR as a mechanism for perturbed cognition in schizophrenia. EMBO Mol. Med. 2012, 4, 1043–1056. [Google Scholar] [CrossRef] [PubMed]
- Renard, J.; Norris, C.; Rushlow, W.; Laviolette, S.R. Neuronal and molecular effects of cannabidiol on the mesolimbic dopamine system: Implications for novel schizophrenia treatments. Neurosci. Biobehav. Rev. 2017, 75, 157–165. [Google Scholar] [CrossRef]
- Renard, J.; Loureiro, M.; Rosen, L.G.; Zunder, J.; De Oliveira, C.; Schmid, S.; Rushlow, W.J.; Laviolette, S.R. Cannabidiol counteracts amphetamine-induced neuronal and behavioral sensitization of the mesolimbic dopamine pathway through a novel mTOR/p70S6 kinase signaling pathway. J. Neurosci. 2016, 36, 5160–5169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Norris, C.; Loureiro, M.; Kramar, C.; Zunder, J.; Renard, J.; Rushlow, W.; Laviolette, S.R. Cannabidiol modulates fear memory formation through interactions with serotonergic transmission in the mesolimbic system. Neuropsychopharmacology 2016, 41, 2839–2850. [Google Scholar] [CrossRef] [PubMed]
- Peres, F.F.; Diana, M.C.; Levin, R.; Suiama, M.A.; Almeida, V.; Vendramini, A.M.; Santos, C.M.; Zuardi, A.W.; Hallak, J.E.C.; Crippa, J.A.; et al. Cannabidiol administered during peri-adolescence prevents behavioral abnormalities in an animal model of schizophrenia. Front. Pharmacol. 2018, 9, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solowij, N.; Broyd, S.J.; Beale, C.; Prick, J.A.; Greenwood, L.M.; Van Hell, H.; Suo, C.; Galettis, P.; Pai, N.; Fu, S.; et al. Therapeutic Effects of Prolonged Cannabidiol Treatment on Psychological Symptoms and Cognitive Function in Regular Cannabis Users: A Pragmatic Open-Label Clinical Trial. Cannabis Cannabinoid Res. 2018, 3, 21–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yücel, M.; Lorenzetti, V.; Suo, C.; Zalesky, A.; Fornito, A.; Takagi, M.J.; Lubman, D.I.; Solowij, N. Hippocampal harms, protection and recovery following regular cannabis use. Transl. Psychiatry 2016, 6, e710. [Google Scholar] [CrossRef] [Green Version]
- Beale, C.; Broyd, S.J.; Chye, Y.; Suo, C.; Schira, M.; Galettis, P.; Martin, J.H.; Yücel, M.; Solowij, N. Prolonged cannabidiol treatment effects on hippocampal subfield volumes in current cannabis users. Cannabis Cannabinoid Res. 2018, 3, 94–107. [Google Scholar] [CrossRef] [PubMed]
- Murphy, M.; Mills, S.; Winstone, J.; Leishman, E.; Wager-Miller, J.; Bradshaw, H.; Mackie, K. Chronic Adolescent Δ9-Tetrahydrocannabinol Treatment of Male Mice Leads to Long-Term Cognitive and Behavioral Dysfunction, Which Are Prevented by Concurrent Cannabidiol Treatment. Cannabis Cannabinoid Res. 2017, 2, 235–246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, H.; Fierro, A.; David Pessoa-Mahana, C. Cannabidiol binding and negative allosteric modulation at the cannabinoid type 1 receptor in the presence of delta-9-tetrahydrocannabinol: An In Silico study. PLoS ONE 2019, 14, e0220025. [Google Scholar] [CrossRef] [PubMed]
- Hudson, R.; Rushlow, W.; Laviolette, S.R. Phytocannabinoids modulate emotional memory processing through interactions with the ventral hippocampus and mesolimbic dopamine system: Implications for neuropsychiatric pathology. Psychopharmacology 2018, 235, 447–458. [Google Scholar] [CrossRef] [PubMed]
- Jung, T.; Hudson, R.; Rushlow, W.; Laviolette, S.R. Functional interactions between cannabinoids, omega-3 fatty acids and peroxisome proliferator-activated receptors: Implications for mental health pharmacotherapies. Eur. J. Neurosci. 2020. [Google Scholar] [CrossRef] [PubMed]
- Campos, A.C.; Fogaça, M.V.; Sonego, A.B.; Guimarães, F.S. Cannabidiol, neuroprotection and neuropsychiatric disorders. Pharmacol. Res. 2016, 112, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Leweke, F.M.; Piomelli, D.; Pahlisch, F.; Muhl, D.; Gerth, C.W.; Hoyer, C.; Klosterkötter, J.; Hellmich, M.; Koethe, D. Cannabidiol enhances anandamide signaling and alleviates psychotic symptoms of schizophrenia. Transl. Psychiatry 2012, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Felice, M.; Melis, M.; Aroni, S.; Muntoni, A.L.; Fanni, S.; Frau, R.; Devoto, P.; Pistis, M. The PPARα agonist fenofibrate attenuates disruption of dopamine function in a maternal immune activation rat model of schizophrenia. CNS Neurosci. Ther. 2019, 25, 549–561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rolland, B.; Marche, K.; Cottencin, O.; Bordet, R. The PPARα Agonist Fenofibrate Reduces Prepulse Inhibition Disruption in a Neurodevelopmental Model of Schizophrenia. Schizophr. Res. Treat. 2012, 2012, 839853. [Google Scholar] [CrossRef] [Green Version]
- Segal-Gavish, H.; Gazit, N.; Barhum, Y.; Ben-Zur, T.; Taler, M.; Hornfeld, S.H.; Gil-Ad, I.; Weizman, A.; Slutsky, I.; Niwa, M.; et al. BDNF overexpression prevents cognitive deficit elicited by adolescent cannabis exposure and host susceptibility interaction. Hum. Mol. Genet. 2017, 26, 2462–2471. [Google Scholar] [CrossRef] [PubMed]
- Jouroukhin, Y.; Zhu, X.; Shevelkin, A.V.; Hasegawa, Y.; Abazyan, B.; Saito, A.; Pevsner, J.; Kamiya, A.; Pletnikov, M.V. Adolescent Δ9-Tetrahydrocannabinol Exposure and Astrocyte-Specific Genetic Vulnerability Converge on Nuclear Factor-κB–Cyclooxygenase-2 Signaling to Impair Memory in Adulthood. Biol. Psychiatry 2019, 85, 891–903. [Google Scholar] [CrossRef] [PubMed]
- Long, L.E.; Chesworth, R.; Huang, X.F.; McGregor, I.S.; Arnold, J.C.; Karl, T. Transmembrane domain Nrg1 mutant mice show altered susceptibility to the neurobehavioural actions of repeated THC exposure in adolescence. Int. J. Neuropsychopharmacol. 2013, 16, 163–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spencer, J.R.; Darbyshire, K.M.E.; Boucher, A.A.; Kashem, M.A.; Long, L.E.; McGregor, I.S.; Karl, T.; Arnold, J.C. Novel molecular changes induced by Nrg1 hypomorphism and Nrg1-cannabinoid interaction in adolescence: A hippocampal proteomic study in mice. Front. Cell. Neurosci. 2013, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shrestha, S.; Offer, S.M. Epigenetic Regulations of GABAergic Neurotransmission: Relevance for Neurological Disorders and Epigenetic Therapy. Med. Epigenet. 2016, 4, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Dunn, A.L.; Michie, P.T.; Hodgson, D.M.; Harms, L. Adolescent cannabinoid exposure interacts with other risk factors in schizophrenia: A review of the evidence from animal models. Neurosci. Biobehav. Rev. 2020, 116, 202–220. [Google Scholar] [CrossRef]
- Zamberletti, E.; Prini, P.; Speziali, S.; Gabaglio, M.; Solinas, M.; Parolaro, D.; Rubino, T. Gender-dependent behavioral and biochemical effects of adolescent delta-9-tetrahydrocannabinol in adult maternally deprived rats. Neuroscience 2012, 204, 245–257. [Google Scholar] [CrossRef] [PubMed]
- Lecca, S.; Luchicchi, A.; Scherma, M.; Fadda, P.; Muntoni, A.L.; Pistis, M. Δ9-Tetrahydrocannabinol during Adolescence Attenuates Disruption of Dopamine Function Induced in Rats by Maternal Immune Activation. Front. Behav. Neurosci. 2019, 13, 202. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Therapeutic Intervention | THC Adolescent Treatment | Outcomes 1 | |
|---|---|---|---|
| Muscimol 500 ng/0.5 μL intra-PFC infusions (male SD rats, >PND 75) | THC 2.5–10 mg/kg i.p. twice daily (male SD rats, PND 35–45) | ↑ novel object recognition index ↑ social motivation and recognition scores ↑ locomotor activity ↓ anxiety ↓ VTA DA firing activity ↓ VTA DA bursting activity | [59] |
| L-theanine 10 mg/kg i.p. twice daily (male SD rats, PND 35–45) | THC 2.5–10 mg/kg i.p. twice daily (male SD rats, PND 35–45) | ↑ sensory motor gating ↑ social memory ↑ novel object recognition index ↑ sucrose preference ↓ anxiety ↓ VTA DA firing activity ↓ PFC pyramidal bursting rate ↑ PFC gamma oscillations ↑ p-GSK3α-β ↑ p-GSK3α-β/t-GSK3α/β | [96] |
| SB258585 2.5 mg/kg or rapamycin 1.5 mg/kg i.p. daily (male C57BL/6J mice, PND 30–45) | THC 5 mg/kg i.p. daily (male C57BL/6J mice, PND 30–45) | ↓ PFC mTOR ↓ PFC p-p70s6K ↑ object discrimination index ↑ sociability and social discrimination index ↑ PFC mIPSC frequency ↓ PFC mEPSC frequency ↓ PFC resting membrane potential ↑ action potential threshold ↑ rheobase ↓ LTD | [99] |
| CBD 3 mg/kg i.p. daily (male CD1 mice, PND 28–48) | THC 3 mg/kg i.p. daily (male CD1 mice, PND 28–48) | ↑ object discrimination index ↓ compulsive behaviors ↓ anxiety | [108] |
| URB597 0.3 mg/kg i.p. daily (female SD rats, >PND 75) | THC 2.5–10 mg/kg i.p. twice daily (female SD rats, PND 35–45) | ↓ immobility, ↑ swimming in FST ↑ sucrose consumption ↑ social interaction = recognition memory | [70] |
| URB597 0.3 mg/kg i.p. daily (female SD rats, >PND 75) | THC 2.5–10 mg/kg i.p. twice daily (female SD rats, PND 35–45) | ↑ PFC CB1Rs ↑ PFC LTD ↑ DG dendritic arborization ↑ DG DCX+ cells ↓ immobility, ↑ swimming in FST | [87] |
| BDNF over-expression in dorsal HC | THC 10 mg/kg i.p. daily (male DN-DISC1 C57BL/6J mice, PND 42–51) | ↑ object recognition memory = locomotor activity = anxiety | [116] |
| NS398 10 mg/kg s.c. daily (male and female astrocytic DN-DISC1 mice, PND 30–51) or NS398 20 μM in culture medium | THC 8 mg/kg s.c. daily (male and female astrocytic DN-DISC1 mice, PND 30–51) or THC 5 mM in culture medium | ↑ spatial memory ↑ novel object recognition index ↑ novel place recognition index ↑ glutamate secretion | [117] |
| Genetic modification in Nrg1 gene | THC 10 mg/kg i.p. daily (male Nrg1 HET mice, PND 30–51) | = locomotor activity = anxiety = PPI and startle reflex ↑ sniffing behavior ↑ Substantia nigra CB1Rs binding ↑ Anterior insula 5HT2A binding | [118] |
| Genetic modification in Nrg1 gene | THC 10 mg/kg i.p. daily (male Nrg1 HET mice, PND 30–51) | ↑ APOA1 ↓ GPSM2 ↓ FLOT1 | [119] |
| Chaetocin 0.05 mg/kg i.p daily (female SD rats, PND 35–45) | THC 2.5–10 mg/kg i.p. twice daily (female SD rats, PND 35–45) | ↑ novel object recognition index = social behavior = immobility, = swimming in FST | [69] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Felice, M.; Laviolette, S.R. Reversing the Psychiatric Effects of Neurodevelopmental Cannabinoid Exposure: Exploring Pharmacotherapeutic Interventions for Symptom Improvement. Int. J. Mol. Sci. 2021, 22, 7861. https://doi.org/10.3390/ijms22157861
De Felice M, Laviolette SR. Reversing the Psychiatric Effects of Neurodevelopmental Cannabinoid Exposure: Exploring Pharmacotherapeutic Interventions for Symptom Improvement. International Journal of Molecular Sciences. 2021; 22(15):7861. https://doi.org/10.3390/ijms22157861
Chicago/Turabian StyleDe Felice, Marta, and Steven R. Laviolette. 2021. "Reversing the Psychiatric Effects of Neurodevelopmental Cannabinoid Exposure: Exploring Pharmacotherapeutic Interventions for Symptom Improvement" International Journal of Molecular Sciences 22, no. 15: 7861. https://doi.org/10.3390/ijms22157861