Beta-Estradiol Regulates Voltage-Gated Calcium Channels and Estrogen Receptors in Telocytes from Human Myometrium

 ,
,  , ,
, , 
Abstract
:
1. Introduction
2. Results
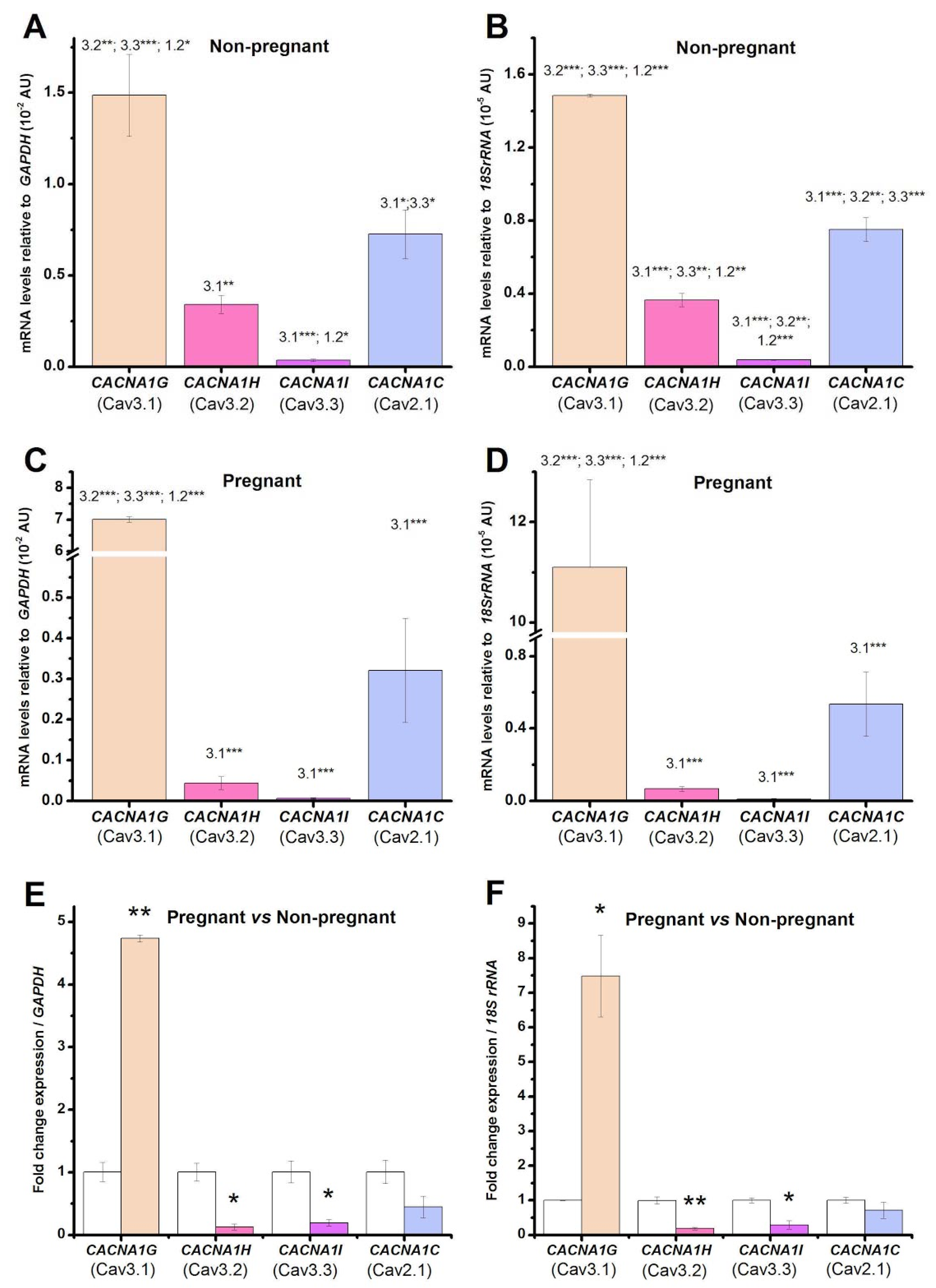
2.1. Pregnancy Induces Changes in mRNA Levels Encoding the Voltage-Gated Calcium Channels Compared to the Non-Pregnant Condition in Human Uterine Myometrial TCs
2.2. Pregnancy Induces Changes in mRNA Levels Encoding the Estrogen Receptors Compared to the Non-Pregnant Condition
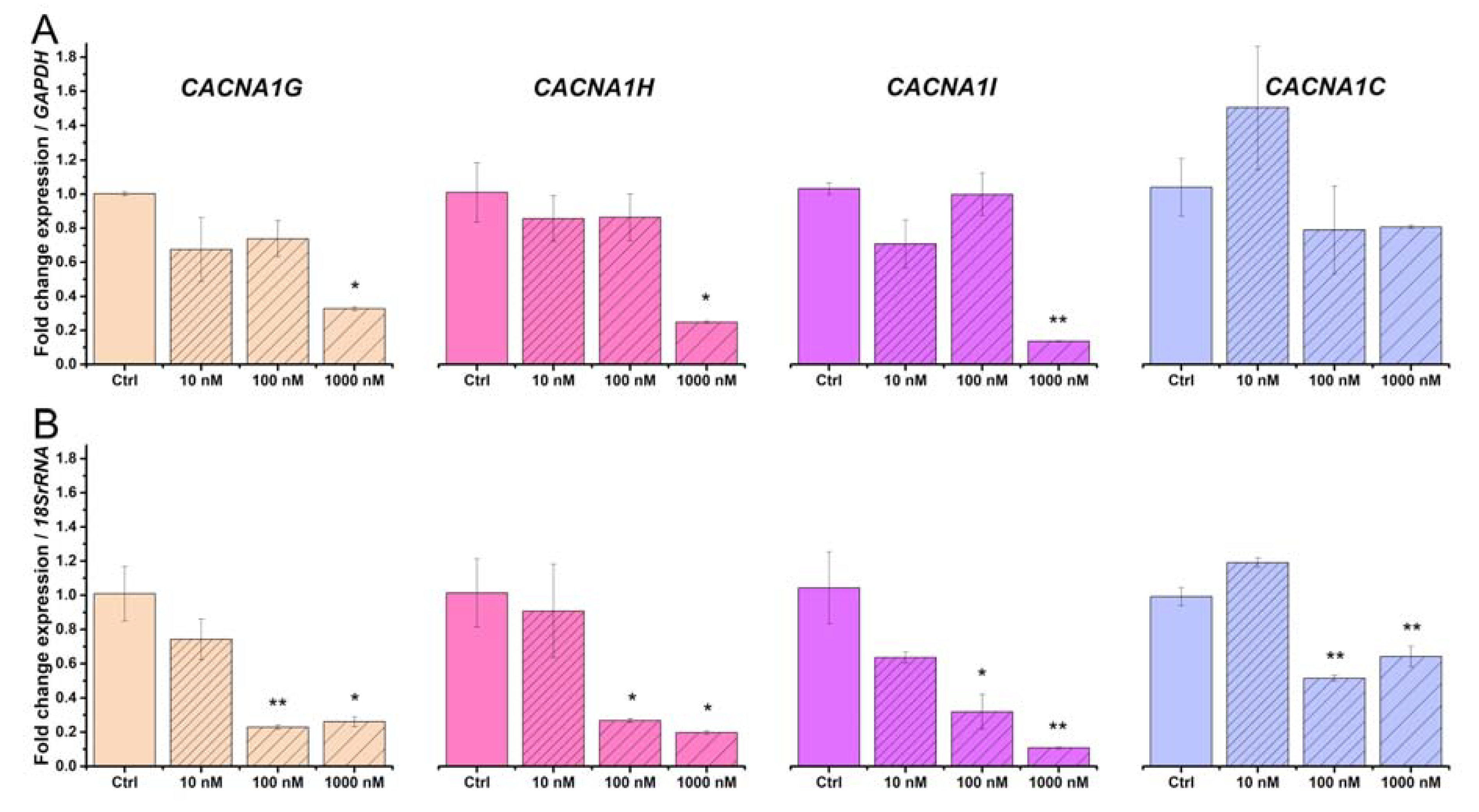
2.3. Beta-Estradiol Downregulates Voltage-Gated Calcium Channels in TCs from Human Pregnant Uterine Myometrial Cultures
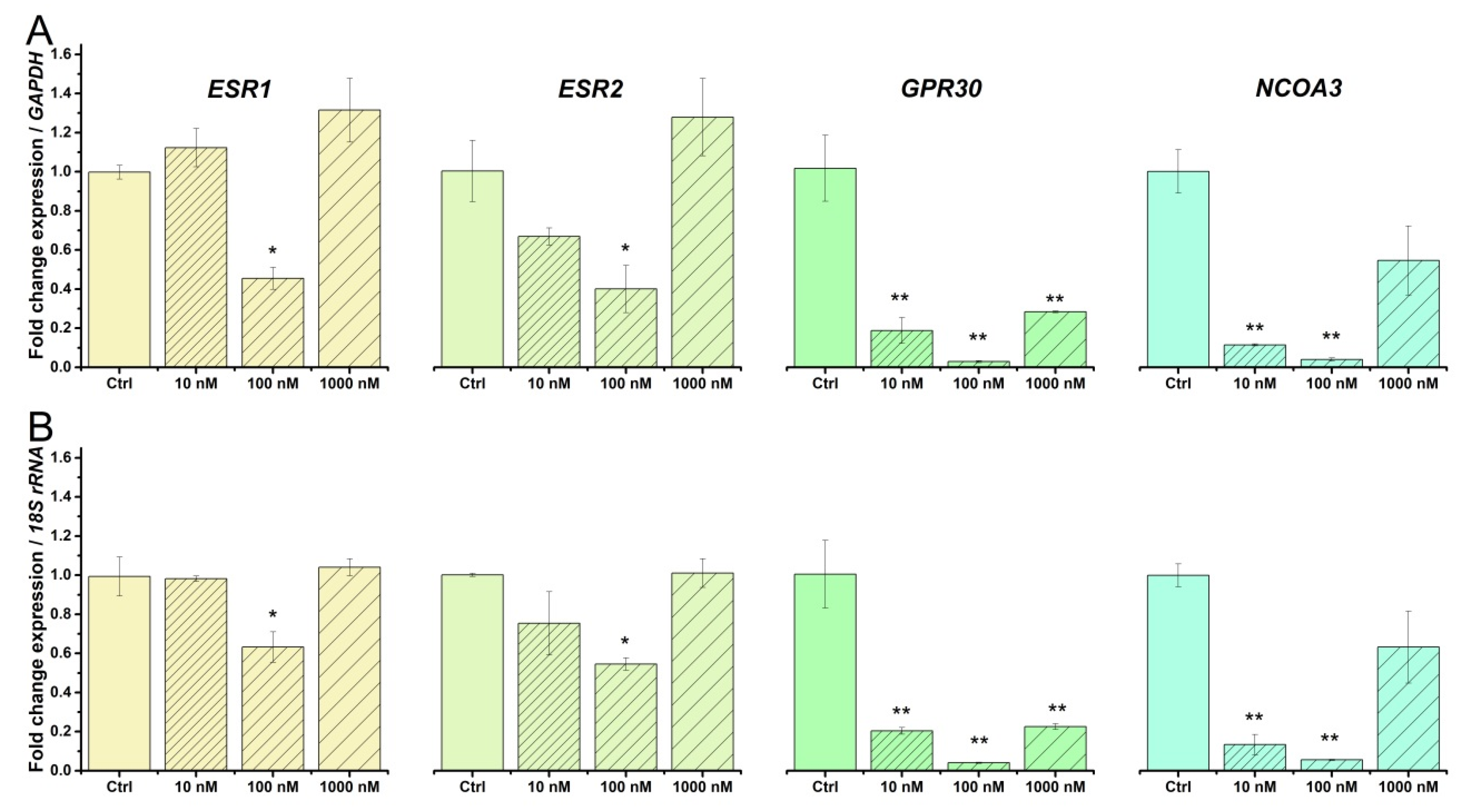
2.4. Beta-Estradiol Downregulates Estrogen Receptors in TCs from Human Pregnant Uterine Myometrial Cultures
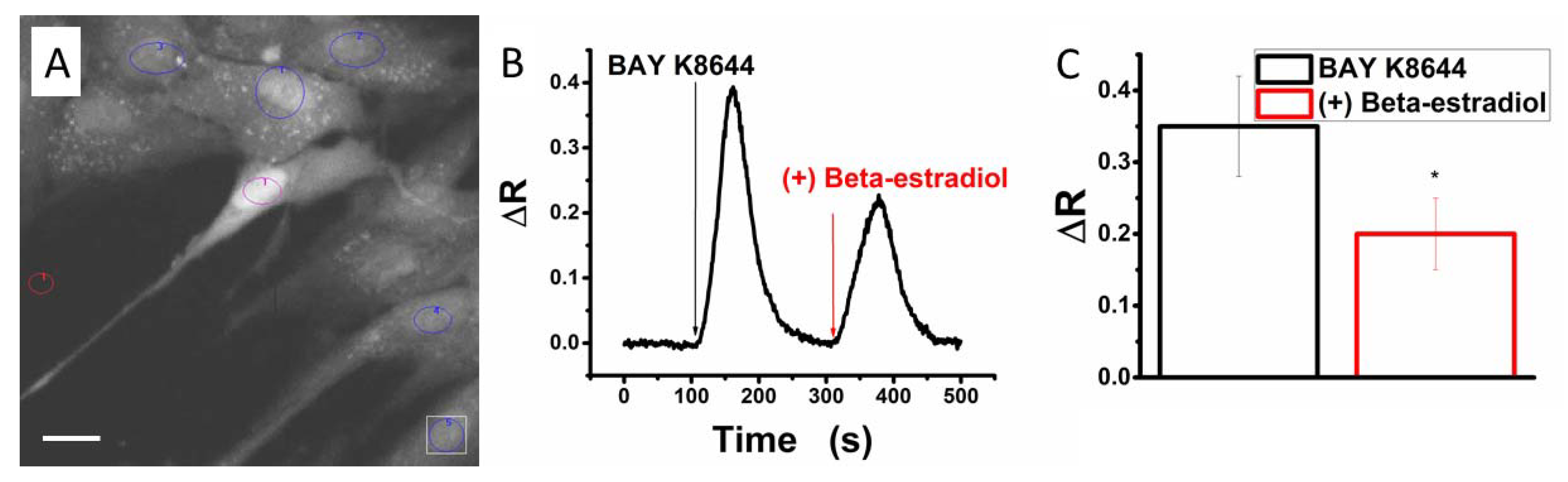
2.5. Beta-Estradiol Partly Blocks Bay K8644-Induced Calcium Transients in Human Myometrial Uterine TCs
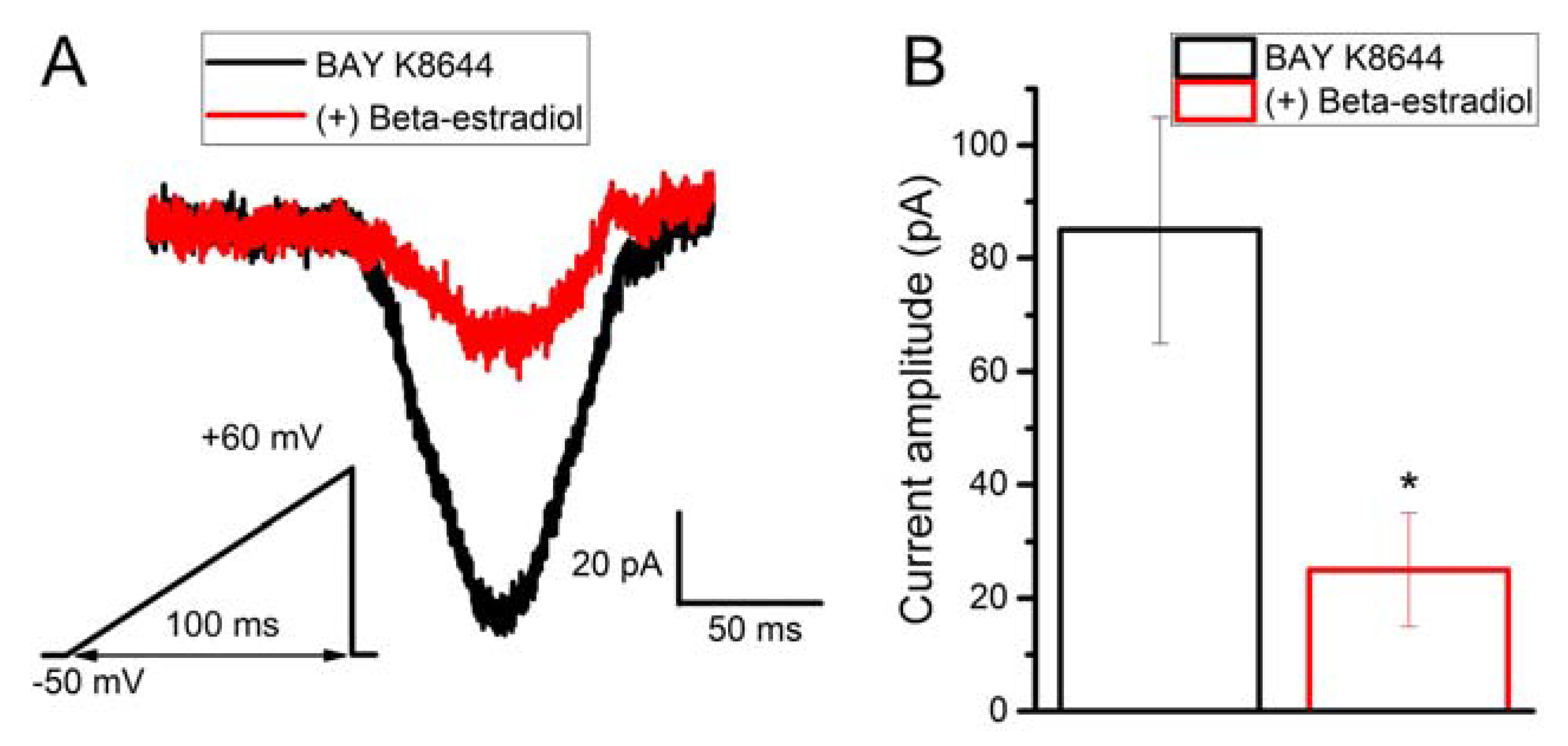
2.6. Beta-Estradiol Partly Inhibits Bay K8644-Induced Calcium Current in Human Myometrial Uterine TCs
3. Discussion
3.1. Beta-Estradiol Regulates Voltage-Gated Calcium Channels in Human Myometrial TCs from Pregnant Uterus
3.2. Beta-Estradiol Regulates Estrogen Receptors in Human Myometrial TCs from Pregnant Uterus
4. Materials and Methods
4.1. Human Uterine Samples
4.2. Myometrial Cell Cultures
4.3. qRT-PCR
4.4. Intracellular Calcium Imaging
4.5. Patch-Clamp Recordings on TCs
4.6. Data Analysis
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Wray, S.; Jones, K.; Kupittayanant, S.; Li, Y.; Matthew, A.; Monir-Bishty, E.; Noble, K.; Pierce, S.J.; Quenby, S.; Shmygol, A.V. Calcium signaling and uterine contractility. J. Soc. Gynecol. Investig. 2003, 10, 252–264. [Google Scholar] [CrossRef]
- Herington, J.L.; Swale, D.R.; Brown, N.; Shelton, E.L.; Choi, H.; Williams, C.H.; Hong, C.C.; Paria, B.C.; Denton, J.S.; Reese, J. High-Throughput Screening of Myometrial Calcium-Mobilization to Identify Modulators of Uterine Contractility. PLoS ONE 2015, 10, e0143243. [Google Scholar] [CrossRef] [PubMed]
- Bernstein, K.; Vink, J.Y.; Fu, X.W.; Wakita, H.; Danielsson, J.; Wapner, R.; Gallos, G. Calcium-activated chloride channels anoctamin 1 and 2 promote murine uterine smooth muscle contractility. Am. J. Obstet. Gynecol. 2014, 211, 688.e1–688.e10. [Google Scholar] [CrossRef] [PubMed]
- Pistilli, M.J.; Petrik, J.J.; Holloway, A.C.; Crankshaw, D.J. Immunohistochemical and functional studies on calcium-sensing receptors in rat uterine smooth muscle. Clin. Exp. Pharmacol. Physiol. 2012, 39, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Ying, L.; Becard, M.; Lyell, D.; Han, X.; Shortliffe, L.; Husted, C.I.; Alvira, C.M.; Cornfield, D.N. The transient receptor potential vanilloid 4 channel modulates uterine tone during pregnancy. Sci. Transl. Med. 2015, 7, 319ra204. [Google Scholar] [CrossRef] [PubMed]
- Wakle-Prabagaran, M.; Lorca, R.A.; Ma, X.; Stamnes, S.J.; Amazu, C.; Hsiao, J.J.; Karch, C.M.; Hyrc, K.L.; Wright, M.E.; England, S.K. BKCa channel regulates calcium oscillations induced by alpha-2-macroglobulin in human myometrial smooth muscle cells. Proc. Natl. Acad. Sci. USA 2016, 113, E2335–E2344. [Google Scholar] [CrossRef] [PubMed]
- Wray, S. Insights from physiology into myometrial function and dysfunction. Exp. Physiol. 2015, 100, 1468–1476. [Google Scholar] [CrossRef] [PubMed]
- Wray, S.; Burdyga, T.; Noble, D.; Noble, K.; Borysova, L.; Arrowsmith, S. Progress in understanding electro-mechanical signalling in the myometrium. Acta Physiol. 2015, 213, 417–431. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.E.; Ahn, D.S.; Lee, Y.H. Role of T-type Ca Channels in the Spontaneous Phasic Contraction of Pregnant Rat Uterine Smooth Muscle. Korean J. Physiol. Pharmacol. 2009, 13, 241–249. [Google Scholar] [CrossRef] [PubMed]
- Li, X.T.; Qiu, X.Y. 17β-Estradiol Upregulated Expression of α and β Subunits of Larger-Conductance Calcium-Activated K(+) Channels (BK) via Estrogen Receptor β. J. Mol. Neurosci. 2015, 56, 799–807. [Google Scholar] [CrossRef] [PubMed]
- Rahbek, M.; Nazemi, S.; Odum, L.; Gupta, S.; Poulsen, S.S.; Hay-Schmidt, A.; Klaerke, D.A. Expression of the small conductance Ca2+-activated potassium channel subtype 3 (SK3) in rat uterus after stimulation with 17β-estradiol. PLoS ONE 2014, 9, e87652. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Ye, Q.; Lu, R.; Cao, J.; Wang, J.; Ding, H.; Gao, R.; Xiao, H. Effects of estradiol on high-voltage-activated Ca(2+) channels in cultured rat cortical neurons. Endocr. Res. 2014, 39, 44–49. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, J.C.; López-Zapata, D.F.; Pinzón, O.A. Effects of 17beta-estradiol and IGF-1 on L-type voltage-activated and stretch-activated calcium currents in cultured rat cortical neurons. Neuroendocrinol. Lett. 2014, 35, 724–732. [Google Scholar] [PubMed]
- Zhang, F.; Ram, J.L.; Standley, P.R.; Sowers, J.R. 17 beta-Estradiol attenuates voltage-dependent Ca2+ currents in A7r5 vascular smooth muscle cell line. Am. J. Physiol. 1994, 266, C975–C980. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, J.C.; López-Zapata, D.F.; Francis, L.; De Los Reyes, L. Effects of estradiol and IGF-1 on the sodium calcium exchanger in rat cultured cortical neurons. Cell. Mol. Neurobiol. 2011, 31, 619–627. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.; Seo, H.; Kim, M.; Ka, H. Dynamic expression of calcium-regulatory molecules, TRPV6 and S100G, in the uterine endometrium during pregnancy in pigs. Biol. Reprod. 2009, 81, 1122–1130. [Google Scholar] [CrossRef] [PubMed]
- Pohóczky, K.; Kun, J.; Szalontai, B.; Szőke, É.; Sághy, É.; Payrits, M.; Kajtár, B.; Kovács, K.; Környei, J.L.; Garai, J.; et al. Estrogen-dependent up-regulation of TRPA1 and TRPV1 receptor proteins in the rat endometrium. J. Mol. Endocrinol. 2016, 56, 135–149. [Google Scholar] [CrossRef] [PubMed]
- Tulchinsky, D.; Korenman, S.G. The plasma estradiol as an index of fetoplacental function. J. Clin. Investig. 1971, 50, 1490–1497. [Google Scholar] [CrossRef] [PubMed]
- Hatthachote, P.; Gillespie, J.I. Complex interactions between sex steroids and cytokines in the human pregnant myometrium: Evidence for an autocrine signaling system at term. Endocrinology 1999, 140, 2533–2540. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.J.; Geimonen, E.; Andersen, J. Increased expression of estrogen receptor beta in human uterine smooth muscle at term. Eur. J. Endocrinol. 2000, 142, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Cretoiu, S.M.; Cretoiu, D.; Popescu, L.M. Human myometrium—The ultrastructural 3D network of telocytes. J. Cell. Mol. Med. 2012, 16, 2844–2849. [Google Scholar] [CrossRef] [PubMed]
- Cretoiu, S.M.; Popescu, L.M. Telocytes revisited. Biomol. Concepts 2014, 5, 353–369. [Google Scholar] [CrossRef] [PubMed]
- Song, D.; Cretoiu, D.; Cretoiu, S.M.; Wang, X. Telocytes and lung disease. Histol. Histopathol. 2016, 31, 1303–1314. [Google Scholar] [PubMed]
- Cretoiu, S.M.; Cretoiu, D.; Marin, A.; Radu, B.M.; Popescu, L.M. Telocytes: Ultrastructural, immunohistochemical and electrophysiological characteristics in human myometrium. Reproduction 2013, 145, 357–370. [Google Scholar] [CrossRef] [PubMed]
- Song, D.; Cretoiu, D.; Zheng, M.; Qian, M.; Zhang, M.; Cretoiu, S.M.; Chen, L.; Fang, H.; Popescu, L.M.; Wang, X. Comparison of Chromosome 4 gene expression profile between lung telocytes and other local cell types. J. Cell. Mol. Med. 2016, 20, 71–80. [Google Scholar] [CrossRef] [PubMed]
- Albulescu, R.; Tanase, C.; Codrici, E.; Popescu, D.I.; Cretoiu, S.M.; Popescu, L.M. The secretome of myocardial telocytes modulates the activity of cardiac stem cells. J. Cell. Mol. Med. 2015, 19, 1783–1794. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Cretoiu, D.; Yan, G.; Cretoiu, S.M.; Popescu, L.M.; Fang, H.; Wang, X. Protein profiling of human lung telocytes and microvascular endothelial cells using iTRAQ quantitative proteomics. J. Cell. Mol. Med. 2014, 18, 1035–1059. [Google Scholar] [CrossRef] [PubMed]
- Cretoiu, S.M.; Cretoiu, D.; Simionescu, A.; Popescu, L.M. Telocytes in human fallopian tube and uterus express estrogen and progesterone receptors. In Sex Steroids; Kahn, S., Ed.; InTech: Rijeka, Croatia, 2012; pp. 91–114. [Google Scholar]
- Roatesi, I.; Radu, B.M.; Cretoiu, D.; Cretoiu, S.M. Uterine Telocytes: A Review of Current Knowledge. Biol. Reprod. 2015, 93, 10. [Google Scholar] [CrossRef] [PubMed]
- Cretoiu, D.; Xu, J.; Xiao, J.; Cretoiu, S.M. Telocytes and Their Extracellular Vesicles-Evidence and Hypotheses. Int. J. Mol. Sci. 2016, 17, 1322. [Google Scholar] [CrossRef] [PubMed]
- Cretoiu, D.; Cretoiu, S.M. Telocytes in the reproductive organs: Current understanding and future challenges. Semin. Cell Dev. Biol. 2016, 55, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Cretoiu, D.; Radu, B.M.; Banciu, A.; Banciu, D.D.; Cretoiu, S.M. Telocytes heterogeneity: From cellular morphology to functional evidence. Semin. Cell Dev. Biol. 2017, 64, 26–39. [Google Scholar] [CrossRef] [PubMed]
- Rusu, M.C.; Cretoiu, D.; Vrapciu, A.D.; Hostiuc, S.; Dermengiu, D.; Manoiu, V.S.; Cretoiu, S.M.; Mirancea, N. Telocytes of the human adult trigeminal ganglion. Cell Biol. Toxicol. 2016, 32, 199–207. [Google Scholar] [CrossRef] [PubMed]
- Radu, B.M.; Banciu, A.; Banciu, D.D.; Radu, M.; Cretoiu, D.; Cretoiu, S.M. Calcium Signaling in Interstitial Cells: Focus on Telocytes. Int. J. Mol. Sci. 2017, 18, 397. [Google Scholar] [CrossRef] [PubMed]
- Cretoiu, S.M.; Radu, B.M.; Banciu, A.; Banciu, D.D.; Cretoiu, D.; Ceafalan, L.C.; Popescu, L.M. Isolated human uterine telocytes: Immunocytochemistry and electrophysiology of T-type calcium channels. Histochem. Cell Biol. 2015, 143, 83–94. [Google Scholar] [CrossRef] [PubMed]
- Campeanu, R.A.; Radu, B.M.; Cretoiu, S.M.; Banciu, D.D.; Banciu, A.; Cretoiu, D.; Popescu, L.M. Near-infrared low-level laser stimulation of telocytes from human myometrium. Lasers Med. Sci. 2014, 29, 1867–1874. [Google Scholar] [CrossRef] [PubMed]
- Othman, E.R.; Elgamal, D.A.; Refaiy, A.M.; Abdelaal, I.I.; Abdel-Mola, A.F.; Al-Hendy, A. Identification and potential role of telocytes in human uterine leiomyoma. Contracept. Reprod. Med. 2016, 20, 12. [Google Scholar] [CrossRef] [PubMed]
- Varga, I.; Klein, M.; Urban, L.; Danihel, L., Jr.; Polak, S.; Danihel, L., Sr. Recently discovered interstitial cells “telocytes” as players in the pathogenesis of uterine leiomyomas. Med. Hypotheses 2018, 110, 64–67. [Google Scholar] [CrossRef] [PubMed]
- Cretoiu, S.M. Immunohistochemistry of Telocytes in the Uterus and Fallopian Tubes. Adv. Exp. Med. Biol. 2016, 913, 335–357. [Google Scholar] [PubMed]
- Vannucchi, M.G.; Faussone-Pellegrini, M.S. The Telocyte Subtypes. Adv. Exp. Med. Biol. 2016, 913, 115–126. [Google Scholar] [PubMed]
- Yang, J.; Li, Y.; Xue, F.; Liu, W.; Zhang, S. Exosomes derived from cardiac telocytes exert positive effects on endothelial cells. Am. J. Transl. Res. 2017, 9, 5375–5387. [Google Scholar] [PubMed]
- Rusu, M.C.; Hostiuc, S.; Vrapciu, A.D.; Mogoantă, L.; Mănoiu, V.S.; Grigoriu, F. Subsets of telocytes: Myocardial telocytes. Ann. Anat. 2017, 209, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Uluer, E.T.; Inan, S.; Ozbilgin, K.; Karaca, F.; Dicle, N.; Sancı, M. The role of hypoxia related angiogenesis in uterine smooth muscle tumors. Biotech. Histochem. 2015, 90, 102–110. [Google Scholar] [CrossRef] [PubMed]
- Sajewicz, M.; Konarska, M.; Wrona, A.N.; Aleksandrovych, V.; Bereza, T.; Komnata, K.; Solewski, B.; Maleszka, A.; Depukat, P.; Warchoł, Ł. Vascular density, angiogenesis and pro-angiogenic factors in uterine fibroids. Folia Med. Cracov. 2016, 56, 27–32. [Google Scholar] [PubMed]
- Salama, N. Immunohistochemical characterization of telocytes in rat uterus in different reproductive states. Egypt J. Histol. 2013, 36, 85–194. [Google Scholar]
- Kim, Y.H.; Chung, S.; Lee, Y.H.; Kim, E.C.; Ahn, D.S. Increase of L-type Ca2+ current by protease-activated receptor 2 activation contributes to augmentation of spontaneous uterine contractility in pregnant rats. Biochem. Biophys. Res. Commun. 2012, 418, 167–172. [Google Scholar] [CrossRef] [PubMed]
- Seda, M.; Pinto, F.M.; Wray, S.; Cintado, C.G.; Noheda, P.; Buschmann, H.; Candenas, L. Functional and molecular characterization of voltage-gated sodium channels in uteri from nonpregnant rats. Biol. Reprod. 2007, 77, 855–863. [Google Scholar] [CrossRef] [PubMed]
- Tica, A.A.; Dun, E.C.; Tica, O.S.; Gao, X.; Arterburn, J.B.; Brailoiu, G.C.; Oprea, T.I.; Brailoiu, E. G protein-coupled estrogen receptor 1-mediated effects in the rat myometrium. Am. J. Physiol. Cell Physiol. 2011, 301, C1262–C1269. [Google Scholar] [CrossRef] [PubMed]
- Silva, E.S.; Scoggin, K.E.; Canisso, I.F.; Troedsson, M.H.; Squires, E.L.; Ball, B.A. Expression of receptors for ovarian steroids and prostaglandin E2 in the endometrium and myometrium of mares during estrus, diestrus and early pregnancy. Anim. Reprod. Sci. 2014, 151, 169–181. [Google Scholar] [CrossRef] [PubMed]
- Kautz, E.; Gram, A.; Aslan, S.; Ay, S.S.; Selçuk, M.; Kanca, H.; Koldaş, E.; Akal, E.; Karakaş, K.; Findik, M.; et al. Expression of genes involved in the embryo-maternal interaction in the early-pregnant canine uterus. Reproduction 2014, 147, 703–717. [Google Scholar] [CrossRef] [PubMed]
- Ilicic, M.; Butler, T.; Zakar, T.; Paul, J.W. The expression of genes involved in myometrial contractility changes during ex situ culture of pregnant human uterine smooth muscle tissue. J. Smooth Muscle Res. 2017, 53, 73–89. [Google Scholar] [CrossRef] [PubMed]
- Vodstrcil, L.A.; Shynlova, O.; Westcott, K.; Laker, R.; Simpson, E.; Wlodek, M.E.; Parry, L.J. Progesterone withdrawal, and not increased circulating relaxin, mediates the decrease in myometrial relaxin receptor (RXFP1) expression in late gestation in rats. Biol. Reprod. 2010, 83, 825–832. [Google Scholar] [CrossRef] [PubMed]
- Welsh, T.; Johnson, M.; Yi, L.; Tan, H.; Rahman, R.; Merlino, A.; Zakar, T.; Mesiano, S. Estrogen receptor (ER) expression and function in the pregnant human myometrium: Estradiol via ERα activates ERK1/2 signaling in term myometrium. J. Endocrinol. 2012, 212, 227–238. [Google Scholar] [CrossRef] [PubMed]
- Chandran, S.; Cairns, M.T.; O’Brien, M.; Smith, T.J. Transcriptomic effects of estradiol treatment on cultured human uterine smooth muscle cells. Mol. Cell. Endocrinol. 2014, 393, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Zhu, J.; Li, X.; Li, S.; Lan, Z.; Ko, J.; Lei, Z. Expression of genomic functional estrogen receptor 1 in mouse sertoli cells. Reprod. Sci. 2014, 21, 1411–1422. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, N.; Yuri, K. Estrogen-dependent changes in estrogen receptor-β mRNA expression in middle-aged female rat brain. Brain Res. 2014, 1543, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Murata, T.; Narita, K.; Ichimaru, T. Rat uterine oxytocin receptor and estrogen receptor α and β mRNA levels are regulated by estrogen through multiple estrogen receptors. J. Reprod. 2014, 60, 55–61. [Google Scholar] [CrossRef]
- Bechem, M.; Hoffmann, H. The molecular mode of action of the Ca agonist (-) BAY K 8644 on the cardiac Ca channel. Pflugers Arch. 1993, 424, 343–353. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.Y.; Chai, Y.G.; Lee, E.B.; Kim, K.W.; Nah, S.Y.; Oh, T.H.; Rhim, H. 17Beta-estradiol inhibits high-voltage-activated calcium channel currents in rat sensory neurons via a non-genomic mechanism. Life Sci. 2002, 70, 2047–2059. [Google Scholar] [CrossRef]
- Bosch, M.A.; Hou, J.; Fang, Y.; Kelly, M.J.; Rønnekleiv, O.K. 17Beta-estradiol regulation of the mRNA expression of T-type calcium channel subunits: Role of estrogen receptor alpha and estrogen receptor beta. J. Comp. Neurol. 2009, 512, 347–358. [Google Scholar] [CrossRef] [PubMed]
- Ravanos, K.; Dagklis, T.; Petousis, S.; Margioula-Siarkou, C.; Prapas, Y.; Prapas, N. Factors implicated in the initiation of human parturition in term and preterm labor: A review. Gynecol. Endocrinol. 2015, 31, 679–683. [Google Scholar] [CrossRef] [PubMed]
- Luoma, J.I.; Kelley, B.G.; Mermelstein, P.G. Progesterone inhibition of voltage-gated calcium channels is a potential neuroprotective mechanism against excitotoxicity. Steroids 2011, 76, 845–855. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Moenter, S.M. Progesterone treatment inhibits and dihydrotestosterone (DHT) treatment potentiates voltage-gated calcium currents in gonadotropin-releasing hormone (GnRH) neurons. Endocrinology 2010, 151, 5349–5358. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Hill, S.J.; Khan, R.N. Oxytocin inhibits T-type calcium current of human decidual stromal cells. J. Clin. Endocrinol. Metab. 2005, 90, 4191–4197. [Google Scholar] [CrossRef] [PubMed]
- Murata, T.; Narita, K.; Honda, K.; Matsukawa, S.; Higuchi, T. Differential regulation of estrogen receptor alpha and beta mRNAs in the rat uterus during pregnancy and labor: Possible involvement of estrogen receptors in oxytocin receptor regulation. Endocr. J. 2003, 50, 579–587. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Chen, Z.; Zhou, X.; Shi, S.; Qi, H.; Baker, P.N.; Zhang, H. Imbalance between proliferation and apoptosis-related impaired GPR30 expression is involved in preeclampsia. Cell Tissue Res. 2016, 366, 499–508. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Zhang, Z.; Liao, H.; Wu, L.; Wu, X.; Zhou, D.; Xi, X.; Zhu, Y.; Feng, Y. Nuclear estrogen receptor-mediated Notch signaling and GPR30-mediated PI3K/AKT signaling in the regulation of endometrial cancer cell proliferation. Oncol. Rep. 2012, 27, 504–510. [Google Scholar] [PubMed]
- Cretoiu, D.; Ciontea, S.M.; Popescu, L.M.; Ceafalan, L.; Ardeleanu, C. Interstitial Cajal-like cells (ICLC) as steroid hormone sensors in human myometrium: Immunocytochemical approach. J. Cell. Mol. Med. 2006, 10, 789–795. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.Y.; Zhang, S.; Li, Y.G.; Wang, Y. Isolation, culture, purification and ultrastructural investigation of cardiac telocytes. Mol. Med. Rep. 2016, 14, 1194–1200. [Google Scholar] [CrossRef] [PubMed]
- Sakaguchi, H.; Fujimoto, J.; Sun, W.S.; Tamaya, T. Clinical implications of steroid receptor coactivator (SRC)-3 in uterine endometrial cancers. J. Steroid Biochem. Mol. Biol. 2007, 104, 237–240. [Google Scholar] [CrossRef] [PubMed]
- Szwarc, M.M.; Kommagani, R.; Lessey, B.A.; Lydon, J.P. The p160/steroid receptor coactivator family: Potent arbiters of uterine physiology and dysfunction. Biol. Reprod. 2014, 91, 122. [Google Scholar] [CrossRef] [PubMed]
- Divekar, S.D.; Storchan, G.B.; Sperle, K.; Veselik, D.J.; Johnson, E.; Dakshanamurthy, S.; Lajiminmuhip, Y.N.; Nakles, R.E.; Huang, L.; Martin, M.B. The role of calcium in the activation of estrogen receptor-alpha. Cancer Res. 2011, 71, 1658–1668. [Google Scholar] [CrossRef] [PubMed]
- Tajbakhsh, A.; Pasdar, A.; Rezaee, M.; Fazeli, M.; Soleimanpour, S.; Hassanian, S.M.; FarshchiyanYazdi, Z.; Younesi Rad, T.; Ferns, G.A.; Avan, A. The current status and perspectives regarding the clinical implication of intracellular calcium in breast cancer. J. Cell. Physiol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Leclercq, G. Calcium-induced activation of estrogen receptor alpha—New insight. Steroids 2012, 77, 924–927. [Google Scholar] [CrossRef] [PubMed]
- Corcoran, J.J.; Nicholson, C.; Sweeney, M.; Charnock, J.C.; Robson, S.C.; Westwood, M.; Taggart, M.J. Human uterine and placental arteries exhibit tissue-specific acute responses to 17β-estradiol and estrogen-receptor-specific agonists. Mol. Hum. Reprod. 2014, 20, 433–441. [Google Scholar] [CrossRef] [PubMed]
- Comunanza, V.; Carbone, E.; Marcantoni, A.; Sher, E.; Ursu, D. Calcium-dependent inhibition of T-type calcium channels by TRPV1 activation in rat sensory neurons. Pflügers Arch. Eur. J. Physiol. 2011, 462, 709–722. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−∆∆Ct. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Gene | Protein Encoded |
|---|---|---|
| Hs00167681_m1 | CACNA1C | Cav1.2, L-type calcium channel |
| Hs00367969_m1 | CACNA1G | Cav3.1, T-type calcium channel |
| Hs00234934_m1 | CACNA1H | Cav3.2, T-type calcium channel |
| Hs00184168_m1 | CACNA1I | Cav3.3, T-type calcium channel |
| Hs01922715_s1 | GPR30, GPER1 | G protein-coupled estrogen receptor 1 |
| Hs01105253_m1 | NCOA3 | Nuclear receptor coactivator 3 |
| Hs01100353_m1 | ESR2 | Estrogen receptor 2 |
| Hs00174860_m1 | ESR1 | Estrogen receptor 1 |
| Hs99999905_m1 | GAPDH | Glyceraldehyde 3-phosphate dehydrogenase |
| Hs99999901_s1 | 18S rRNA | 18S ribosomal RNA |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Banciu, A.; Banciu, D.D.; Mustaciosu, C.C.; Radu, M.; Cretoiu, D.; Xiao, J.; Cretoiu, S.M.; Suciu, N.; Radu, B.M. Beta-Estradiol Regulates Voltage-Gated Calcium Channels and Estrogen Receptors in Telocytes from Human Myometrium. Int. J. Mol. Sci. 2018, 19, 1413. https://doi.org/10.3390/ijms19051413
Banciu A, Banciu DD, Mustaciosu CC, Radu M, Cretoiu D, Xiao J, Cretoiu SM, Suciu N, Radu BM. Beta-Estradiol Regulates Voltage-Gated Calcium Channels and Estrogen Receptors in Telocytes from Human Myometrium. International Journal of Molecular Sciences. 2018; 19(5):1413. https://doi.org/10.3390/ijms19051413
Chicago/Turabian StyleBanciu, Adela, Daniel Dumitru Banciu, Cosmin Catalin Mustaciosu, Mihai Radu, Dragos Cretoiu, Junjie Xiao, Sanda Maria Cretoiu, Nicolae Suciu, and Beatrice Mihaela Radu. 2018. "Beta-Estradiol Regulates Voltage-Gated Calcium Channels and Estrogen Receptors in Telocytes from Human Myometrium" International Journal of Molecular Sciences 19, no. 5: 1413. https://doi.org/10.3390/ijms19051413