
Abstract
Mu-opioid receptors (MOPRs) are critically involved in the modulation of pain and analgesia, and represent a candidate mechanism for the development of biomarkers of pain conditions and their responses to treatment. To further understand the human implications of genetic variation within the opioid system in pain and opioid-mediated placebo responses, we investigated the association between the functional single-nucleotide polymorphism (SNP) in the μ-opioid receptor gene (OPRM1), A118G, and psychophysical responses, personality traits, and neurotransmitter systems (dopamine (DA), opioid) related to pain and placebo analgesia. OPRM1 G carriers, compared with AA homozygotes, showed an overall reduction of baseline μ-opioid receptor availability in regions implicated in pain and affective regulation. In response to a sustained painful stimulus, we found no effect of A118G on pain-induced endogenous opioid release. Instead, AA homozygotes showed a blunted DA response in the nucleus accumbens (NAc) in response to the pain challenge. After placebo administration, G carriers showed more pronounced mood disturbances and lower placebo-induced μ-opioid system activation in the anterior insula (aINS), the amygdala (AMY), the NAc, the thalamus (THA), and the brainstem, as well as lower levels of DA D2/3 activation in the NAc. At a trait level, G carriers reported higher NEO-Neuroticism scores; a personality trait previously associated with increased pain and lower placebo responses, which were negatively correlated with baseline μ-opioid receptor availability in the aINS and subgenual anterior cingulate cortex (sgACC). Our results demonstrate that the A118G OPRM1 polymorphism contributes to interindividual variations in the function of neurotransmitters responsive to pain (endogenous opioid and dopamine), as well as their regulation through cognitive-emotional influences in the context of therapeutic expectations, the so-called placebo effect. These effects are relevant to human vulnerability to disease processes where these neurotransmitters have a role, such as persistent pain, mood, and substance use disorders, and responses to their treatments.
Similar content being viewed by others

INTRODUCTION
Mu-opioid receptors (MOPRs), the primary site of action of endogenous opioid peptides (Zadina et al, 1997), have been consistently implicated in the neurobiology of pain and placebo analgesic effects (Benedetti et al, 2005; Zubieta et al, 2001). Encoded by the OPRM1 gene (6q24-q25), MOPRs are widely distributed in the brain and attain their highest levels in the thalamus (THA) and in components of the limbic system and basal ganglia, including the amygdala (AMY), nucleus accumbens (NAc), and anterior cingulate cortex (ACC) (Oroszi and Goldman, 2004). Genetic variation of the OPRM1 has been associated with pain and analgesia, drug addiction, and response to their treatments (Mague and Blendy, 2010) as well as social stressors (Way et al, 2009); however, the molecular mechanism underlying these associations is poorly understood.
In humans, a commonly investigated single-nucleotide polymorphism (SNP, rs1799971) occurs in exon 1 of the OPRM1, in which an adenine to guanine substitution (A118G) exchanges an asparagine for an aspartic acid at a putative N-glycosylation site (N40D) (Bond et al, 1998). Initial studies in vitro revealed that expression of the human G118 OPRM1 variant in AV-12 cells increased the binding affinity and function of the receptor for β-endorphin, compared with the A118 OPRM1 (Bond et al, 1998). However, those initial results were not replicated in subsequent studies, which appear to point to the G118 variant as associated with loss of function. Zhang et al (2005) reported a 1.5-fold reduction in allele-specific mRNA expression in post-mortem brain tissue and a 10-fold reduction in protein levels in CHO cells expressing the G118 variant; Kroslak et al (2007) also demonstrated that cell culture systems expressing the G118 allele had lower surface receptor expression, decreased forskolin-induced cAMP activation, and lower agonist-induced MOPR activation. In a study using a knock-in mouse equivalent (A112G, N38D) of the human SNP, A/A and G/G mice did not differ in MOPRs binding affinity for β-endorphin, morphine, and naloxone; however, G/G mice had reduced MOPR mRNA brain expression, lower MOPR protein levels and reduced antinociceptive responses to morphine compared with A/A mice (Mague et al, 2009). A different study using quantitative in vitro autoradiography, showed that MOPR levels were lower in G/G compared with A/A mice, in the cingulate and INS, AMY, periaqueductal gray (PAG), the NAc and the ventral tegmental area (VTA) (Wang et al, 2012). In humans, neuroimaging studies using the μ-opioid agonist selective radiotracer [11C]carfentanil and positron emission tomography (PET) showed data consistent with loss of function for the G allele carriers. In a sample of tobacco smokers, Ray et al (2011) reported reduced MOPR availability in vivo in G carriers, compared with AA homozygote, and Weerts et al (2013) described similar findings in patients with alcohol dependence. However, these studies have only been performed in affected samples, and similar studies in healthy humans are lacking to clarify the effects of this generic variant in the absence of interactions with disease processes.
Here, we investigated the molecular mechanisms underlying the effects of the OPRM1 A118G on psychophysical measures of pain and placebo responses, both of which are known to be regulated by MOPRs and DA interactions (Scott et al, 2008), in a sample of 50 healthy subjects. We used in vivo receptor measures with PET and the μ-opioid and D2/3 receptor selective radiotracers [11C]carfentanil and [11C]raclopride, at baseline and during pain- and placebo-induced opioid and DA release. On the basis of the above literature, compared with AA homozygotes, G carriers were expected to show lower μ-opioid receptor availability at baseline. Because of the excitatory effects of MOR activation on DA neuronal excitability (through GABA interneurons) in the VTA (Johnson and North, 1992), we also hypothesized lower pain and placebo-induced endogenous opioid and DA release and MOPR and D2/3 activation, as well as greater pain report and lower analgesic response to the placebo. We also aimed to investigate the effects of the OPRM1 A118G on personality traits that might explain interindividual variability in responses to pain and placebo. Specifically, we hypothesized a genetic effect of A118G on Neuroticism, the tendency to experience negative emotions in stressful situations. Neuroticism has been consistently associated with increased negative affect and neural responses to negative stimuli (Canli, 2004; Norris et al, 2007), as well as pain catastrophizing, pain-related fear (Goubert et al, 2004), and the development of chronic somatic and visceral pain following injury (Taylor et al, 2010) or surgery (Bisgaard et al, 2001). In addition, higher Neuroticism scores have predicted lower placebo effects and opioid release in response to placebo treatment (Pecina et al, 2013a). On the basis of this evidence, we hypothesized higher Neuroticism scores in G carriers, compared with AA homozygotes, by virtue of lower μ-opioid receptor availability at baseline.
METHODS
Subjects and Genotyping
Fifty right-handed, healthy, non-smoking subjects (29F/21M, age: (mean±1SD): 26±0.7) were recruited via advertisement. In addition to completing physical and neurological examinations, study participants were screened using the non-patient version of the Structured Clinical Interview for DSM-IV. Participants had no history of or current medical, neurological, or psychiatric illnesses, including substance abuse, or dependence, and had alcohol intake of less than five drinks per week. Women had regular menstrual cycles of 26–32 days’ duration and had not used hormonal birth control for at least 1 year. We attempted to study women during the follicular phase using menstrual cycle diaries at screening and confirmatory progesterone levels at each PET scanning session (progesterone levels ∼ or <1 ng/ml were confirmed in 23 of the 29 women who completed a PET scan). Written informed consent was obtained in all cases. All of the procedures used were approved by the University of Michigan Investigational Review Board for Human Subject Use and the Radioactive Drug Research Committee.
Results on subjects in the current sample were part of several previous manuscripts examining the effects of placebo administration on μ-opioid and DA neurotransmission (Scott et al, 2008), personality predictors of placebo effects (Pecina et al, 2012a), the cognitive mechanisms involved in placebo analgesia (Pecina et al, 2013b), the role of opioid neurotransmission in the recall of placebo analgesia (Pecina et al, 2012b), and the association between placebo effects and genetic variation within the endocannabinoid system (Pecina et al, 2014). Main effects of placebo in the current sample replicated those in the previous study (Scott et al, 2008) and therefore are not reported here.
Genotyping
Genotyping was successfully performed in all the participants using the Illumina Golden Gate platform at 1536-plex on Sentrix arrays with the Addictions Array content of 130 genes and 186 Ancestry Informative Markers (AIMs) described elsewhere (Hodgkinson et al, 2008). Clustering and genotype calling for each locus were manually verified and loci with call rates <90% were excluded. AIM scores were calculated by comparison with similarly derived genotype data from the CEPH Diversity panel using a minimum of 160 markers. Genotyping accuracy was confirmed by replicate genotyping of 10% of the total sample. Completion rates were >93% (mean 99.4%, median 100%) and replicates showed no errors at these loci.
A118G MOPR genotyped subjects were assigned to two groups: individuals homozygous for the major A118 allele (AA genotype: n=36) and individuals carrying one or two copies of the variant G118 allele (AG: n=12 or GG genotype: n=2). A118G OPRM1 genotype distribution was in Hardy–Weinberg equilibrium (H-W eq., χ2=0.57, p=0.44), and there were no significant differences with respect to sex (F/M) (G carriers: 9/5; AA: 20/16; χ2=0.31, p=0.75), the European AIM scores (mean±SD, G carriers: 0.7±0.4; AA: 0.7±0.4; t=0.13, p=0.9) or age (G carriers: 25±5; AA: 26±5; t=−0.75, p=0.4) among the two genotype groups.
To test for population stratification, we performed Spearman correlations between the European AIM scores (since the sample was predominantly Caucasian) and the neuroimaging data, the psychophysical measures and the NEO-PI scores. No confounding was present due to ethnic differences.
Experimental Design
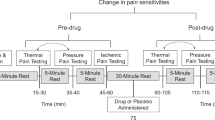
As previously described (Scott et al, 2008), participants completed a total of four 90 min PET scanning sessions (Siemens HR+, Knoxville, Tennessee), two with [11C]carfentanil (n=50) and two with [11C]raclopride (n=49, PET data were missing for one of the conditions), during a pain challenge with and without placebo administration. Each scanning session, with and without placebo administration, consisted of a control condition (baseline: 0.9% isotonic saline, 5–25 min after start of scanning infused in the right masseter muscle) and a painful condition (5% hypertonic saline, 45–65 min after start of scanning), infused in the left masseter muscle (see Supplementary Figure 1).
Subjects were placed in the scanner gantry with needles placed in both masseter muscles and two intravenous (antecubital) lines ∼30 min before radiotracer administration. In the pain condition, a steady state of moderate muscle pain was maintained for 20 min by a computer-controlled delivery system through the infusion of medication-grade hypertonic saline solution (5%) into the left masseter muscle. In this model of sustained deep somatic pain, the intensity of the painful stimulus is standardized across subjects (Stohler and Kowalski, 1999; Zhang et al, 1993) (see Supplementary Methods). The same individual infusion profiles generated during the pain challenges were used for the studies with placebo administration (Scott et al, 2008), maintaining the algesic input the same across both, the pain and the pain+placebo condition.
During the placebo condition, subjects were given the following instructions before its administration: ‘We are studying the effect of a potential pain relief medication. This medication is thought to have analgesic effects through the activation of natural brain systems that suppress pain.’ The placebo condition consisted of the introduction of 1 ml of 0.9% isotonic saline into one of the intravenous ports every 4 min, with the volunteer being made aware by a computer-generated human voice recording, starting 2 min before the pain challenges, and lasting for 15 s each time.
The primary end point for the assessment of placebo responses was a 0–100 visual analog scale (VAS) of momentary pain intensity rating acquired every 15 s during the pain challenge in the absence and presence of placebo, and the McGill Pain Questionnaire (MPQ) (Melzack and Torgerson, 1971), administered immediately after the pain and the pain+placebo challenges. The internal affective state of the volunteers was rated with the Positive and Negative Affectivity Scale (PANAS) (Watson et al, 1988) and the Profile of Mood States (POMS) before and after the scanning session. The composite POMS Total Mood Disturbance score was used to evaluate transient elevations in negative mood (POMS-TMD) (McNair et al, 1992).
Neuroimaging Methods
After image reconstruction (see Supplementary Methods), three receptor-related measures were calculated for each radiotracer: (1) baseline μ-opioid and DA D2/3 BPND during the control condition; (2) μ-opioid and DA D2/3 BPND during the pain challenge, and (3) μ-opioid and DA D2/3 BPND during the pain+placebo challenge. Activation of the μ-opioid and DA D2/3 systems during pain and placebo administration was assessed by calculating the difference between the control and pain conditions and between pain and pain+placebo conditions, respectively. Reductions in the in vivo availability of μ-opioid and DA receptors after an acute challenge are thought to reflect opioid and DA release and competition between radiotracer and endogenous ligand for the receptor sites (Narendran and Martinez, 2008).
Data Analysis
A mixed-effects analysis of variance was applied on a voxel-by-voxel basis for μ-opioid and D2/3 receptor maps, including four within-subject conditions (pain and pain+placebo condition and their corresponding baseline conditions) and A118G OPRM1 as the between-subject factor. A priori hypothesized regions involved in MOPR regulation of pain and during placebo analgesia (ACC, DLPFC and orbitofrontal (OFC) cortices, insula (INS), NAc, AMY, medial THA, and PAG) (Scott et al, 2008) were deemed significant at p<0.0001, and an extent threshold (K) of>10 voxels. For other regions, significant differences were detected using a p<0.05 FWE or FDR-corrected (Friston et al, 1994).
The regional data were extracted for quantification of regional changes in BPND, graphing, and determination of correlation coefficients (Pearson/Spearman correlations at P<0.05 for normal and non-normally distributed data, respectively). Imaging and behavioral statistical analyses were controlled for pain intensity (average VAS rated by the volunteers every 15 s for the duration of the challenges), as variables of no interest.
Trait Measures (NEO-PI-R)
We assessed the effects of A118G on the Neuroticism domain of the NEO-PI-R (Costa and McCrae, 1992), defined as the tendency to experience negative emotions, particularly in stressful situations (Costa and McCrae, 1980) and previously associated with the risk for developing chronic pain (Taylor et al, 2010) and to experience lower placebo analgesic effects (Pecina et al, 2013a). We further investigated the effect of A118G OPRM1 on each of the components of the Neuroticism domain (Anxiety, Hostility, Depression, Self-Consciousness, Impulsiveness, and Vulnerability to Stress).
RESULTS
State Measures: OPRM1 A118G Effects on Baseline Affect and Pain- and Placebo-Induced Changes in Affect and Pain Ratings
We found no effect of A118G either in the reported pain experience (measured with the MPQ, average VAS scores, the volume of hypertonic saline to achieve average target pain ratings, or the ratio between VAS ratings and the volume infused to achieve the target ratings) or in the positive or negative affect scales (PANAS) or mood disturbance (POMS) at baseline or in their change after the pain challenge.
Instead, we identified a significant effect of OPRM1 A118G (AA>AG/GG) on changes in mood disturbance after placebo administration, as measured with the POMS-TMD, but not with the PANAS. Compared with G carriers, AA homozygotes showed greater reductions in POMS-TMD scores after placebo (POMS-TMD (mean diff.±SD), AA: −0.8±1.4; G carriers: 2.8±0.9; F=5.9, p=0.02).
OPRM1 A118G Effects on μ-Opioid and D2/3 BPND at Baseline
Analyses of the molecular imaging data showed an OPRM1 A118G genotype effect on μ-opioid receptor BPND at baseline, but no effect on D2/3 BPND. We observed a significant effect of OPRM1 A118G (AA> G carriers) on baseline μ-opioid receptor BPND extensively throughout the brain (Figure 1; Table 1) in regions that included cortically the DLPFC, the dorso/ventromedial (DM/VM) PFC, the OFC, the inferior frontal gyrus (IFG), the rostral, subgenual and dorsal (r/sg/d) ACC, the mid- and posterior cingulate cortex (MCC and PCC), the anterior and posterior INS, the AMY and parahippocampal (paraHipp) gyrus and hippocampus (Hipp). Subcortically, effects in the same direction were observed in the NAc/ventral pallidum (vPAL), the caudate (CAU), and the THA. No significant effect was observed for the contrast OPRM1 G carriers> AA homozygotes.

OPRM1 A118G effect on μ-opioid receptor availability at baseline (AA>G carriers). AA homozygotes, compared with G carriers, showed greater μ-opioid receptor binding in regions that included the anterior cingulate cortex (subgenual, rostral, and dorsal ACC), the ventral striatum (NAc: nucleus accumbens) and the thalamus (THA) among others (see Table 1).
Lower μ-opioid receptor concentrations in the PCC, subgenual anterior cingulate cortex (sgACC), and vmPFC were negatively correlated with higher pain reports (PCC (MPQ: r=−0.3, p=0.03; MPQ sensory: r=−0.34, p=0.01)) and POMS-TMD ratings (sgACC: r=−0.3, p=0.03; vmPFC: r=−0.3, p=0.03), although, these correlations were uncorrected for multiple comparisons. μ-Opioid receptor BPND was not associated with reductions in pain ratings after placebo administration.
OPRM1 A118G Effects on Pain and Placebo Induced Changes in μ-Opioid and D2/3 BPND
No significant gene effect was observed on pain-induced endogenous opioid release. Conversely, compared with G carriers, AA homozygotes showed an overall effect of DA deactivation in the NAc in response to the pain challenge (Table 2, Figure 2). However, changes in pain-induced DA release in the NAc were not correlated with changes in affective state or subjective pain reports.

OPRM1 A118G effect on changes in D2/3 BPND during the pain challenge. AA homozygotes, compared with G carriers, showed blunted DA response in the NAc during the pain challenge.
Conversely, we observed a significant effect of OPRM1 A118G (AA> AG/GG) during placebo-induced μ-opioid system activation in the anterior insula (aINS), AMY, NAc, THA, and the brainstem/PAG region (Table 2; Figure 3). Placebo-induced activation of μ-opioid neurotransmission in the NAc was positively correlated with reductions in the sensory/affective/total dimensions of pain (MPQ sensory, r=0.3, p=0.03; MPQ affect: r=0.37, p=0.01; MPQ total: r=0.3, p=0.02) and in POMS-TMD scores of the volunteers (r=0.30, p=0.03). POMS-TMD scores were also associated with the magnitude of placebo-induced endogenous opioid release in the THA (r=0.29, p=0.04). In the same direction, AA homozygotes, compared with G carriers, showed greater placebo-induced activation of DA D2/3 neurotransmission in the NAc (Table 2; Figure 3) that were not correlated with changes in the affective state or subjective pain reports after placebo administration.

OPRM1 A118G effect on changes in μ-opioid and D2/3 BPND during placebo administration. AA homozygotes, compared with G carriers, showed greater placebo induced μ-opioid (a, d) and D2/3 (b, d) activation systems in the NAc after placebo. AA homozygotes showed lower scores in the NEO-Depression and NEO-Vulnerability facets of the NEO-Neuroticism domain (c).
In addition, changes in μ-opioid and D2/3 DA neurotransmission during the pain and the placebo challenges were significantly correlated with μ-opioid BPND at baseline. While opioid BPND at baseline was positively associated with placebo-induced opioid release in the aINS, AMY, NAc, THA, and brainstem and DA release in the NAc, it was negatively correlated with pain-induced DA release in the NAc (Table 1).
Trait Measures: OPRM1 A118G Effects on the NEO-Neuroticism Domain (Vulnerability and Depression Facets)
A significant effect of A118G (G carriers> AA) was observed for the Neuroticism domain of the NEO Personality Inventory (Costa and McCrae, 1992) (AA: 75±4.4; G carriers: 93.6±7.3, F=4.7, p=0.035). Then, we tested for an effect of A118G OPRM1 on each of the components of the Neuroticism domain (Anxiety, Hostility, Depression, Self-Consciousness, Impulsiveness and Vulnerability to Stress). In the same direction, we found a significant effect of A118G on the Vulnerability to Stress (AA: 8.4±0.7; G carriers: 12.6±1.2, F=8.8, p=0.005) and as a trend, the Depression scores (AA: 11.0±1.0; G carriers: 15.5±1.8, F=3.8, p=0.056), where G carriers showed higher scores compared with AA homozygotes.
Although uncorrected for multiple comparison, higher NEO-Neuroticism scores were negatively correlated with baseline μ-opioid receptor BPND in regions involved in affect and stress regulation (sgACC: r=−0.25; p=0.05, aINS: r=−0.28; p=0.03), as were its facets NEO-Vulnerability (sgACC: r=−0.28; p=0.03, aINS: r=−0.33; p=0.01, Hipp: r=−0.33; p=.01) and NEO-Depression (sgACC: r=−0.28; p=0.03, aINS: r=−0.27; p=0.04). Placebo-induced opioid release in the brainstem was negatively correlated with NEO-vulnerability (r=−0.30; p=0.04). No significant associations were obtained between these trait measures and pain-induced DA activation.
DISCUSSION
The OPRM1 A118G SNP has been implicated in the pathophysiology of a range of disorders, such as pain and analgesia, drug addiction, and responses to their treatments (Mague and Blendy, 2010). In this manuscript, we provide initial evidence for some of the molecular mechanisms that appear to underlie these associations in healthy humans. We identified a vulnerability phenotype in G carriers, defined by an overall reduction in μ-opioid receptor BPND, lower responses of endogenous opioid and DA systems to placebo during expectation of analgesia, and higher NEO-Neuroticism scores (driven by Vulnerability to Stress and Depression facets). AA homozygotes, compared with G carriers, showed a blunted DA response in the NAc in response to the sustained pain challenge that suggest a reduction in NAc DA responses to an emotionally negatively salient stimulus (Scott et al, 2007).
OPRM1 118G carriers, compared with AA homozygotes, showed an overall reduction of MOR BPND in multiple regions implicated in pain, stress, and emotion regulation (Zubieta et al, 2001). These results are consistent with previous human postmortem (Zhang et al, 2005) and neuroimaging studies in alcohol dependence (Weerts et al, 2013), tobacco smokers (Ray et al, 2011), and the role of μ-opioid receptors in the suppression of stress, pain and negative emotional states (Prossin et al, 2010; Zubieta et al, 2003b; Zubieta et al, 2001). Similar reductions (NAc, AMY, and dorsal ACC) have also been observed in patients with fibromyalgia (Harris et al, 2007), a persistent pain syndrome frequently comorbid with mood disturbances. Consistently with our results, Harris et al (2007) showed that reductions in μ-opioid availability in vivo in limbic regions were associated with greater clinical pain ratings, and in particular the affective quality of pain (Zubieta et al, 2001). In the present study, we observed that OPRM1 118G carriers, compared with AA homozygotes, reported higher Neuroticism scores, specifically Vulnerability to Stress and Depression facets, that were correlated reductions in μ-opioid receptor availability in the sgACC and aINS (although these correlations were uncorrected for multiple comparisons). Importantly, Neuroticism has been consistently associated with increased negative effect, anxiety, and autonomic and neural responses to negative stimuli (Canli, 2004; Norris et al, 2007), the development of chronic pain after injury (Taylor et al, 2010) or surgery (Bisgaard et al, 2001), and lower μ-opioid mediated response to placebo (Pecina et al, 2013a).
Surprisingly, we did not observe an effect of the OPRM1 A118G on the subjective pain experience or changes in opioid neurotransmission during the sustained pain challenge. In previous work, this polymorphism has been linked to altered pain thresholds and analgesic responses to opioid administration. In particular, the G118 allele has been associated with greater pain ratings after cesarean surgery (Sia et al, 2008; Tan et al, 2009), as well as reduced responses to opioid drugs after surgery (Chou et al, 2006a, 2006b; Oertel et al, 2006; Sia et al, 2008; Tan et al, 2009) or during experimental pain (Oertel et al, 2006). The results presented here suggest that these associations are probably related to the lower receptor availability of μ-opioid receptors in G-allele carriers, as described above, but not to effects on presynaptic endogenous opioid function, since A118G showed no association with pain-induced activation of opioid neurotransmission. Alternatively, the effects of A118G on pain thresholds and analgesic responses could also be explained by indirect effects of A118G on DA neurotransmission. OPRM1 AA homozygotes, compared with 118G carriers, showed a blunted DA response in the NAc in responses to the pain challenge. The NAc is thought to have a broad role in responding to and modulating salient stimuli with both rewarding and non-rewarding valences (Berridge and Robinson, 1998; Pruessner et al, 2004; Scott et al, 2007; Zald et al, 2004). Studies on the neurobiology of pain have shown that both DA and opioid peptides regulate responses to painful stimuli in the NAc (Gear et al, 1999; Horvitz, 2000; Zubieta et al, 2003a), and therefore could explain the relationship between this polymorphism and altered pain thresholds described previously (Sia et al, 2008; Tan et al, 2009). However, this blunted DA response in OPRM1 AA homozygotes in the NAc was not associated with the subjective pain experience of the participants, supporting the hypothesis that the effects of A118G on pain thresholds are mediated by the reductions in μ-opioid receptor availability.
Paradoxically, given the above results, we observed an effect of A118G on placebo-induced endogenous opioid system activation. OPRM1 G carriers, compared with AA homozygotes, showed greater mood disturbance after placebo administration as measured with the POMS-TMD, and reduced endogenous opioid release in the aINS, AMY, NAc, THA and brainstem/PAG, and DA release in the NAc. Previous studies have described the regulatory role of μ-receptors and MOPR-mediated neurotransmission in clinical and experimental pain (Harris et al, 2007; Zubieta et al, 2001) and placebo-induced analgesic responses (Zubieta et al, 2005). In these studies, placebo-induced endogenous opioid release in similar regions was associated with the suppression of both the sensory and affective qualities of pain, and the negative affective state induced by sustained experimental pain. Here, we observed a selective effect of the OPRM1 A118G polymorphism on placebo-induced, but not pain-induced endogenous opioid release. Albeit this was an unexpected result, it is likely to reflect the involvement of this neurotransmitter system in cognitive and emotional processes that take place during social interactions and environmental cues (Hsu et al, 2013; Panksepp and Beatty, 1980), beyond its role in responses to stress. These social and environmental cues are thought to form an essential part of the therapeutic millieu linked to the formation of placebo effects in humans (Benedetti, 2013; Kelley et al, 2009). During the administration of the placebo, AA homozygotes, compared with G carriers, in addition to showing greater MOPR-mediated neurotransmission in a number of reward-related regions, showed an increased DA response to the positive valence challenge (placebo administration) in the NAc. It is worth highlighting that the findings on the effects of OPRM1 A118G gene on DA neurotransmission in response to placebo administration robustly map onto the neurobiology of opioid and DA interactions in reward processing. DA neurons in the VTA that project to the NAc are part of a well-defined pathway involved in reward responses and encoding (Nestler, 2005). GABAergic interneurons in the VTA maintain a tonic inhibition over DA neurons. Agonist-induced activation of MOPRs on these interneurons reduces their activity, resulting in disinhibition of the DA neurons and elevations of DA release in the NAc (Shoji et al, 1999). Moreover, the results presented here point to a valence-specific effect of OPRM1 A118G on DA neurotransmission in the NAc, where G carriers showed greater DA release in the NAc in response to the pain challenge but reductions in DA release in the same regions in responses to the placebo challenge. Our results also suggest that these effects might be under the regulation of the opioid system, as suggested by the relationship between DA release in responses to both challenges and μ-opioid receptor availability across multiple brain regions.
In this manuscript, we provide a mechanistic explanation of the effect of the OPRM1 A118G on the regulation of pain, placebo analgesia and stress-related personality traits. These findings point to a potentially significant role of OPRM1 A118G in illnesses where vulnerability to physical and emotional stressors represents a core feature, which include persistent pain conditions, the addictions and the mood disorders, as well as their comorbidities. This genotype may therefore serve as a potential biomarker of vulnerability for those conditions and inter-individual variations in responses to their treatments. These data also add to an emerging literature on genetic influences on the formation of placebo analgesic effects (Hall et al, 2012; Pecina et al, 2014). If confirmed in larger samples, then genetic tests could be used as biomarkers that allow the prediction of placebo responses in clinical trials, helping decisions regarding patient stratification and the contribution of trait factors to the separation of drug and placebo treatment effects.
FUNDING AND DISCLOSURE
Work was supported by R01 DA 022520, R01 DA 27494 (JKZ), and the Phil F. Jenkins Foundation. The authors declare no conflict of interest.
References
Benedetti F (2013). Placebo and the new physiology of the doctor-patient relationship. Physiol Rev 93: 1207–1246.
Benedetti F, Mayberg HS, Wager TD, Stohler CS, Zubieta JK (2005). Neurobiological mechanisms of the placebo effect. J Neurosci 25: 10390–10402.
Berridge KC, Robinson TE (1998). What is the role of dopamine in reward: hedonic impact, reward learning, or incentive salience? Brain Res Brain Res Rev 28: 309–369.
Bisgaard T, Klarskov B, Rosenberg J, Kehlet H (2001). Characteristics and prediction of early pain after laparoscopic cholecystectomy. Pain 90: 261–269.
Bond C, LaForge KS, Tian M, Melia D, Zhang S, Borg L et al (1998). Single-nucleotide polymorphism in the human mu opioid receptor gene alters beta-endorphin binding and activity: possible implications for opiate addiction. Proc Natl Acad Sci USA 95: 9608–9613.
Canli T (2004). Functional brain mapping of extraversion and neuroticism: learning from individual differences in emotion processing. J Pers 72: 1105–1132.
Chou WY, Wang CH, Liu PH, Liu CC, Tseng CC, Jawan B (2006a). Human opioid receptor A118G polymorphism affects intravenous patient-controlled analgesia morphine consumption after total abdominal hysterectomy. Anesthesiology 105: 334–337.
Chou WY, Yang LC, Lu HF, Ko JY, Wang CH, Lin SH et al (2006b). Association of mu-opioid receptor gene polymorphism (A118G) with variations in morphine consumption for analgesia after total knee arthroplasty. Acta Anaesthesiol Scand 50: 787–792.
Costa PT Jr., McCrae RR (1980). Influence of extraversion and neuroticism on subjective well-being: happy and unhappy people. J Pers Soc Psychol 38: 668–678.
Costa PT, McCrae RR (1992) Revised NEO Personality Inventory (NEO-PI-R) and NEO Five-Factor Inventory Professional Manual. Psychological Assessment Resources: Odessa, FL.
Friston KJ, Worsley KJ, Frackowiak RS, Mazziotta JC, Evans AC (1994). Assessing the significance of focal activations using their spatial extent. Hum Brain Mapp 1: 210–220.
Gear RW, Aley KO, Levine JD (1999). Pain-induced analgesia mediated by mesolimbic reward circuits. J Neurosci 19: 7175–7181.
Goubert L, Crombez G, Van Damme S (2004). The role of neuroticism, pain catastrophizing and pain-related fear in vigilance to pain: a structural equations approach. Pain 107: 234–241.
Hall KT, Lembo AJ, Kirsch I, Ziogas DC, Douaiher J, Jensen KB et al (2012). Catechol-O-methyltransferase val158met polymorphism predicts placebo effect in irritable bowel syndrome. PLoS ONE 7: e48135.
Harris RE, Clauw DJ, Scott DJ, McLean SA, Gracely RH, Zubieta JK (2007). Decreased central mu-opioid receptor availability in fibromyalgia. J Neurosci 27: 10000–10006.
Hodgkinson CA, Yuan Q, Xu K, Shen PH, Heinz E, Lobos EA et al (2008). Addictions biology: haplotype-based analysis for 130 candidate genes on a single array. Alcohol Alcohol 43: 505–515.
Horvitz JC (2000). Mesolimbocortical and nigrostriatal dopamine responses to salient non-reward events. Neuroscience 96: 651–656.
Hsu DT, Sanford BJ, Meyers KK, Love TM, Hazlett KE, Wang H et al (2013). Response of the mu-opioid system to social rejection and acceptance. Mol Psychiatry 18: 1211–1217.
Johnson SW, North RA (1992). Opioids excite dopamine neurons by hyperpolarization of local interneurons. J Neurosci 12: 483–488.
Kelley JM, Lembo AJ, Ablon JS, Villanueva JJ, Conboy LA, Levy R et al (2009). Patient and practitioner influences on the placebo effect in irritable bowel syndrome. Psychosom Med 71: 789–797.
Kroslak T, Laforge KS, Gianotti RJ, Ho A, Nielsen DA, Kreek MJ (2007). The single nucleotide polymorphism A118G alters functional properties of the human mu opioid receptor. J Neurochem 103: 77–87.
Mague SD, Blendy JA (2010). OPRM1 SNP (A118G): involvement in disease development, treatment response, and animal models. Drug Alcohol Depend 108: 172–182.
Mague SD, Isiegas C, Huang P, Liu-Chen LY, Lerman C, Blendy JA (2009). Mouse model of OPRM1 (A118G) polymorphism has sex-specific effects on drug-mediated behavior. Proc Natl Acad Sci USA 106: 10847–10852.
McNair D, Lorr M, Droppleman I (1992) EdITS Manual for the Profile of Mood States. Educational and Industrial Testing Service (EdITS): San Diego.
Melzack R, Torgerson WS (1971). On the language of pain. Anesthesiology 34: 50–59.
Narendran R, Martinez D (2008). Cocaine abuse and sensitization of striatal dopamine transmission: a critical review of the preclinical and clinical imaging literature. Synapse 62: 851–869.
Nestler EJ (2005). Is there a common molecular pathway for addiction? Nat Neurosci 8: 1445–1449.
Norris CJ, Larsen JT, Cacioppo JT (2007). Neuroticism is associated with larger and more prolonged electrodermal responses to emotionally evocative pictures. Psychophysiology 44: 823–826.
Oertel BG, Schmidt R, Schneider A, Geisslinger G, Lotsch J (2006). The mu-opioid receptor gene polymorphism 118A>G depletes alfentanil-induced analgesia and protects against respiratory depression in homozygous carriers. Pharmacogenet Genomics 16: 625–636.
Oroszi G, Goldman D (2004). Alcoholism: genes and mechanisms. Pharmacogenomics 5: 1037–1048.
Panksepp J, Beatty WW (1980). Social deprivation and play in rats. Behav Neural Biol 30: 197–206.
Pecina M, Azhar H, Love TM, Lu T, Fredrickson BL, Stohler CS et al (2012a). Personality trait predictors of placebo analgesia and neurobiological correlates. Neuropsychopharmacology 38: 639–646.
Pecina M, Azhar H, Love TM, Lu T, Fredrickson BL, Stohler CS et al (2013a). Personality trait predictors of placebo analgesia and neurobiological correlates. Neuropsychopharmacology 38: 639–646.
Pecina M, Martinez-Jauand M, Hodgkinson C, Stohler CS, Goldman D, Zubieta JK (2014). FAAH selectively influences placebo effects. Mol Psychiatry 19: 385–391.
Pecina M, Stohler CS, Zubieta JK (2012b). Role of mu-opioid system in the formation of memory of placebo responses. Mol Psychiatry 18: 135–137.
Pecina M, Stohler CS, Zubieta JK (2013b). Neurobiology of placebo effects: expectations or learning? Soc Cogn Affect Neurosci 9: 1013–1021.
Prossin AR, Love TM, Koeppe RA, Zubieta JK, Silk KR (2010). Dysregulation of regional endogenous opioid function in borderline personality disorder. Am J Psychiatry 167: 925–933.
Pruessner JC, Champagne F, Meaney MJ, Dagher A (2004). Dopamine release in response to a psychological stress in humans and its relationship to early life maternal care: a positron emission tomography study using [11C]raclopride. J Neurosci 24: 2825–2831.
Ray R, Ruparel K, Newberg A, Wileyto EP, Loughead JW, Divgi C et al (2011). Human Mu Opioid Receptor (OPRM1 A118G) polymorphism is associated with brain mu-opioid receptor binding potential in smokers. Proc Natl Acad Sci USA 108: 9268–9273.
Scott DJ, Stohler CS, Egnatuk CM, Wang H, Koeppe RA, Zubieta JK (2007). Individual differences in reward responding explain placebo-induced expectations and effects. Neuron 55: 325–336.
Scott DJ, Stohler CS, Egnatuk CM, Wang H, Koeppe RA, Zubieta JK (2008). Placebo and nocebo effects are defined by opposite opioid and dopaminergic responses. Arch Gen Psychiatry 65: 220–231.
Shoji Y, Delfs J, Williams JT (1999). Presynaptic inhibition of GABA(B)-mediated synaptic potentials in the ventral tegmental area during morphine withdrawal. J Neurosci 19: 2347–2355.
Sia AT, Lim Y, Lim EC, Goh RW, Law HY, Landau R et al (2008). A118G single nucleotide polymorphism of human mu-opioid receptor gene influences pain perception and patient-controlled intravenous morphine consumption after intrathecal morphine for postcesarean analgesia. Anesthesiology 109: 520–526.
Stohler CS, Kowalski CJ (1999). Spatial and temporal summation of sensory and affective dimensions of deep somatic pain. Pain 79: 165–173.
Tan EC, Lim EC, Teo YY, Lim Y, Law HY, Sia AT (2009). Ethnicity and OPRM variant independently predict pain perception and patient-controlled analgesia usage for post-operative pain. Mol Pain 5: 32.
Taylor KS, Anastakis DJ, Davis KD (2010). Chronic pain and sensorimotor deficits following peripheral nerve injury. Pain 151: 582–591.
Wang YJ, Huang P, Ung A, Blendy JA, Liu-Chen LY (2012). Reduced expression of the mu opioid receptor in some, but not all, brain regions in mice with OPRM1 A112G. Neuroscience 205: 178–184.
Watson D, Clark LA, Tellegen A (1988). Development and validation of brief measures of positive and negative affect: the PANAS scales. J Pers Soc Psychol 54: 1063–1070.
Way BM, Taylor SE, Eisenberger NI (2009). Variation in the mu-opioid receptor gene (OPRM1) is associated with dispositional and neural sensitivity to social rejection. Proc Natl Acad Sci USA 106: 15079–15084.
Weerts EM, McCaul ME, Kuwabara H, Yang X, Xu X, Dannals RF et al (2013). Influence of OPRM1 Asn40Asp variant (A118G) on [11C]carfentanil binding potential: preliminary findings in human subjects. Int J Neuropsychopharmacol 16: 47–53.
Zadina JE, Hackler L, Ge LJ, Kastin AJ (1997). A potent and selective endogenous agonist for the mu-opiate receptor. Nature 386: 499–502.
Zald DH, Boileau I, El-Dearedy W, Gunn R, McGlone F, Dichter GS et al (2004). Dopamine transmission in the human striatum during monetary reward tasks. J Neurosci 24: 4105–4112.
Zhang X, Ashton-Miller JA, Stohler CS (1993). A closed-loop system for maintaining constant experimental muscle pain in man. IEEE Trans Biomed Eng 40: 344–352.
Zhang Y, Wang D, Johnson AD, Papp AC, Sadee W (2005). Allelic expression imbalance of human mu opioid receptor (OPRM1) caused by variant A118G. J Biol Chem 280: 32618–32624.
Zubieta JK, Bueller JA, Jackson LR, Scott DJ, Xu Y, Koeppe RA et al (2005). Placebo effects mediated by endogenous opioid activity on mu-opioid receptors. J Neurosci 25: 7754–7762.
Zubieta JK, Heitzeg MM, Smith YR, Bueller JA, Xu K, Xu Y et al (2003a). COMT val158met genotype affects mu-opioid neurotransmitter responses to a pain stressor. Science 299: 1240–1243.
Zubieta JK, Ketter TA, Bueller JA, Xu Y, Kilbourn MR, Young EA et al (2003b). Regulation of human affective responses by anterior cingulate and limbic mu-opioid neurotransmission. Arch Gen psychiatry 60: 1145–1153.
Zubieta JK, Smith YR, Bueller JA, Xu Y, Kilbourn MR, Jewett DM et al (2001). Regional mu opioid receptor regulation of sensory and affective dimensions of pain. Science 293: 311–315.
Acknowledgements
We would also like to acknowledge the contribution of the technologists of the PET Center at the University of Michigan.
Author information
Authors and Affiliations
Corresponding author
Additional information
Supplementary Information accompanies the paper on the Neuropsychopharmacology website
Supplementary information
Rights and permissions
About this article

Cite this article
Peciña, M., Love, T., Stohler, C. et al. Effects of the Mu Opioid Receptor Polymorphism (OPRM1 A118G) on Pain Regulation, Placebo Effects and Associated Personality Trait Measures. Neuropsychopharmacol 40, 957–965 (2015). https://doi.org/10.1038/npp.2014.272
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/npp.2014.272
This article is cited by
-
Is OPRM1 genotype a valuable predictor of VAS in patients undergoing laparoscopic radical resection of colorectal cancer with fentanyl?
BMC Anesthesiology (2023)
-
A mesocorticolimbic signature of pleasure in the human brain
Nature Human Behaviour (2023)
-
Effects of placebo administration on immune mechanisms and relationships with central endogenous opioid neurotransmission
Molecular Psychiatry (2022)
-
Unique and joint associations of polygenic risk for major depression and opioid use disorder with endogenous opioid system function
Neuropsychopharmacology (2022)
-
Personality traits and polymorphisms of genes coding neurotransmitter receptors or transporters: review of single gene and genome-wide association studies
Annals of General Psychiatry (2021)

{kind=link}