
Abstract
Mitochondria are highly dynamic organelles that undergo frequent fusion and fission. Optic atrophy 1 (OPA1) is an essential GTPase protein for both mitochondrial inner membrane (IM) fusion and cristae morphology1,2. Under mitochondria-stress conditions, membrane-anchored L-OPA1 is proteolytically cleaved to form peripheral S-OPA1, leading to the selection of damaged mitochondria for mitophagy2,3,4. However, molecular details of the selective mitochondrial fusion are less well understood. Here, we showed that L-OPA1 and cardiolipin (CL) cooperate in heterotypic mitochondrial IM fusion. We reconstituted an in vitro membrane fusion reaction using purified human L-OPA1 protein expressed in silkworm, and found that L-OPA1 on one side of the membrane and CL on the other side are sufficient for fusion. GTP-independent membrane tethering through L-OPA1 and CL primes the subsequent GTP-hydrolysis-dependent fusion, which can be modulated by the presence of S-OPA1. These results unveil the most minimal intracellular membrane fusion machinery. In contrast, independent of CL, a homotypic trans-OPA1 interaction mediates membrane tethering, thereby supporting the cristae structure. Thus, multiple OPA1 functions are modulated by local CL conditions for regulation of mitochondrial morphology and quality control.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 print issues and online access
$209.00 per year
only $17.42 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout





Similar content being viewed by others

References
Friedman, J. R. & Nunnari, J. Mitochondrial form and function. Nature 505, 335–343 (2014).
Pernas, L. & Scorrano, L. Mito-morphosis: mitochondrial fusion, fission, and cristae remodeling as key mediators of cellular function. Annu. Rev. Physiol. 78, 505–531 (2016).
MacVicar, T. & Langer, T. OPA1 processing in cell death and disease—the long and short of it. J. Cell. Sci. 129, 2297–2306 (2016).
Ishihara, N., Fujita, Y., Oka, T. & Mihara, K. Regulation of mitochondrial morphology through proteolytic cleavage of OPA1. EMBO J. 25, 2966–2977 (2006).
McNew, J. A., Sondermann, H., Lee, T., Stern, M. & Brandizzi, F. GTP-dependent membrane fusion. Annu. Rev. Cell. Dev. Biol. 29, 529–550 (2013).
Delettre, C. et al. Nuclear gene OPA1, encoding a mitochondrial dynamin-related protein, is mutated in dominant optic atrophy. Nat. Genet. 26, 207–210 (2000).
Anand, R. et al. The i-AAA protease YME1L and OMA1 cleave OPA1 to balance mitochondrial fusion and fission. J. Cell. Biol. 204, 919–929 (2014).
Griparic, L., van der Wel, N. N., Orozco, I. J., Peters, P. J. & van der Bliek, A. M. Loss of the intermembrane space protein Mgm1/OPA1 induces swelling and localized constrictions along the lengths of mitochondria. J. Biol. Chem. 279, 18792–18798 (2004).
Frezza, C. et al. OPA1 controls apoptotic cristae remodeling independently from mitochondrial fusion. Cell 126, 177–189 (2006).
McQuibban, G. A., Saurya, S. & Freeman, M. Mitochondrial membrane remodelling regulated by a conserved rhomboid protease. Nature 423, 537–541 (2003).
Herlan, M., Bornhovd, C., Hell, K., Neupert, W. & Reichert, A. S. Alternative topogenesis of Mgm1 and mitochondrial morphology depend on ATP and a functional import motor. J. Cell. Biol. 165, 167–173 (2004).
DeVay, R. M. et al. Coassembly of Mgm1 isoforms requires cardiolipin and mediates mitochondrial inner membrane fusion. J. Cell. Biol. 186, 793–803 (2009).
Meeusen, S. et al. Mitochondrial inner-membrane fusion and crista maintenance requires the dynamin-related GTPase Mgm1. Cell 127, 383–395 (2006).
Kajikawa, M. et al. Efficient silkworm expression of human GPCR (nociceptin receptor) by a Bombyx mori bacmid DNA system. Biochem. Biophys. Res. Commun. 385, 375–379 (2009).
Ardail, D. et al. Mitochondrial contact sites. Lipid composition and dynamics. J. Biol. Chem. 265, 18797–18802 (1990).
Meglei, G. & McQuibban, G. A. The dynamin-related protein Mgm1p assembles into oligomers and hydrolyzes GTP to function in mitochondrial membrane fusion. Biochemistry 48, 1774–1784 (2009).
Oh, K. J. et al. Conformational changes in BAK, a pore-forming proapoptotic Bcl-2 family member, upon membrane insertion and direct evidence for the existence of BH3-BH3 contact interface in BAK homo-oligomers. J. Biol. Chem. 285, 28924–28937 (2010).
Song, Z., Ghochani, M., McCaffery, J. M., Frey, T. G. & Chan, D. C. Mitofusins and OPA1 mediate sequential steps in mitochondrial membrane fusion. Mol. Biol. Cell. 20, 3525–3532 (2009).
Claypool, S. M. Cardiolipin, a critical determinant of mitochondrial carrier protein assembly and function. Biochim. Biophys. Acta 1788, 2059–2068 (2009).
Lutter, M. et al. Cardiolipin provides specificity for targeting of tBid to mitochondria. Nat. Cell. Biol. 2, 754–761 (2000).
Choi, S. Y. et al. A common lipid links Mfn-mediated mitochondrial fusion and SNARE-regulated exocytosis. Nat. Cell. Biol. 8, 1255–1262 (2006).
Chu, C. T. et al. Cardiolipin externalization to the outer mitochondrial membrane acts as an elimination signal for mitophagy in neuronal cells. Nat. Cell. Biol. 15, 1197–1205 (2013).
Weber, T. et al. SNAREpins: minimal machinery for membrane fusion. Cell 92, 759–772 (1998).
Mima, J. & Wickner, W. Complex lipid requirements for SNARE- and SNARE chaperone-dependent membrane fusion. J. Biol. Chem. 284, 27114–27122 (2009).
Koshiba, T. et al. Structural basis of mitochondrial tethering by mitofusin complexes. Science 305, 858–862 (2004).
Ishihara, N., Eura, Y. & Mihara, K. Mitofusin 1 and 2 play distinct roles in mitochondrial fusion reactions via GTPase activity. J. Cell. Sci. 117, 6535–6546 (2004).
Orso, G. et al. Homotypic fusion of ER membranes requires the dynamin-like GTPase atlastin. Nature 460, 978–983 (2009).
Patten, D. A. et al. OPA1-dependent cristae modulation is essential for cellular adaptation to metabolic demand. EMBO J. 33, 2676–2691 (2014).
Ban, T., Heymann, J. A., Song, Z., Hinshaw, J. E. & Chan, D. C. OPA1 disease alleles causing dominant optic atrophy have defects in cardiolipin-stimulated GTP hydrolysis and membrane tubulation. Hum. Mol. Genet. 19, 2113–2122 (2010).
Schlame, M. & Ren, M. The role of cardiolipin in the structural organization of mitochondrial membranes. Biochim. Biophys. Acta 1788, 2080–2083 (2009).
Mishra, P., Carelli, V., Manfredi, G. & Chan, D. C. Proteolytic cleavage of Opa1 stimulates mitochondrial inner membrane fusion and couples fusion to oxidative phosphorylation. Cell. Metab. 19, 630–641 (2014).
Eckert, D. M. & Kim, P. S. Mechanisms of viral membrane fusion and its inhibition. Annu. Rev. Biochem. 70, 777–810 (2001).
Ban, T. & Ishihara, N. Expression and purification of recombinant human L-OPA1 using BmNPV bacmid-silkworm expression system. Protoc. Exch. http://dx.doi.org/10.1038/protex.2017.053 (2017).
Mima, J., Hickey, C. M., Xu, H., Jun, Y. & Wickner, W. Reconstituted membrane fusion requires regulatory lipids, SNAREs and synergistic SNARE chaperones. EMBO J. 27, 2031–2042 (2008).
Sugiura, S. & Mima, J. Physiological lipid composition is vital for homotypic ER membrane fusion mediated by the dynamin-related GTPase Sey1p. Sci. Rep. 6, 20407 (2016).
Eura, Y., Ishihara, N., Yokota, S. & Mihara, K. Two mitofusin proteins, mammalian homologues of FZO, with distinct functions are both required for mitochondrial fusion. J. Biochem. 134, 333–344 (2003).
Saita, S. et al. Distinct types of protease systems are involved in homeostasis regulation of mitochondrial morphology via balanced fusion and fission. Genes Cells 21, 408–424 (2016).
Acknowledgements
We thank J. Mima (Osaka University) for advice on the preparation of assays with proteoliposomes, K. Tabata (Kyushu University) for advice on the silkworm expression system, Kurume University EM laboratory, and H. Yagi (Tottori University) and D. Ozawa (Osaka University) for helping with the EM observations, and W. Nishihira for technical support. This work was supported by JSPS KAKENHI grant numbers 26291044 and 16H01209 (N.I.), and 26840026 (T.B.), MEXT-Supported Program for the Strategic Research Foundation at Private Universities, the Takeda Science Foundation (N.I.), the Ono Medical Research Foundation (N.I.) and the Ishibashi Foundation (T.B.).
Author information
Authors and Affiliations
Contributions
N.I. and T.B. designed the research. T.B., T.I., H.K., S.S. and A.I. performed the experiments. K.Maenaka established and provided the bacmid expression vector system, and T.O. prepared basic procedures for expression and reconstitution of membrane proteins. K.Mihara supervised the study. N.I. and T.B. wrote the manuscript with the comments of the other authors.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Integrated supplementary information
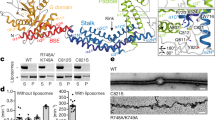
Supplementary Figure 1 Preparation of recombinant OPA1 and OPA1-dependent fusion in vitro or in vivo.
(a) Procedures for the fractionation and purification of recombinant OPA1. (b,c) SDS-PAGE analysis and Coomassie blue staining of L-OPA1 (b) and S-OPA1 (c). Asterisk: OPA1 protein. Fractionation experiments were carried out more than 10 times for L-OPA1 and more than 3 times for S-OPA1 with similar results. (d) Calculation of fusion activity by fitting to the exponential curve with the trace from the experiments shown in Fig. 2a. In the 1:4,000 condition, ∼25 L-OPA1 molecules, which might form ∼2.5 complexes (Fig. 1h) in each liposome (the number of lipid molecules ∼100,000)35, could mediate membrane fusion. The goodness of fit was evaluated by the R2 value. (e) Preparation of the OPA1 KO cell line using the CRISPR/Cas9 system. Whole cell homogenates from OPA1 WT or KO HeLa cells were prepared and subjected to immunoblot analysis with the indicated antibodies (the experiment was performed one time). (f) OPA1 WT or KO HeLa cells were fixed, permeabilized, subjected to immunofluorescence analysis with antibodies to TOM20 (red), and examined by confocal microscopy (representative images of more than 3 independent experiments with similar results). Magnified images are shown in the boxed regions. Scale bars, 10 μm. (g,h) Heterotypic mitochondrial fusion in HeLa cells. Mitochondrial fusion was analyzed in cell hybrids from cells expressing mitGFP and mitRFP using OPA1-deficient or control HeLa cells. After 6 h culture, fusion efficiency was analyzed by fluorescence microscopy (g) and cell counting (h). n, number of cell hybrids analyzed from 3 independent experiments: WT × WT (n = 30), WT × CRISPR-OPA1 (n = 30), and CRISPR-OPA1 × CRISPR-OPA1 (n = 30). Scale bar: 10 μm. Statistics source data are available in Supplementary Table 1. Unprocessed original scans of blot is shown in Supplementary Fig. 3.
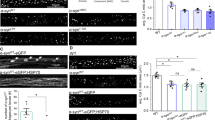
Supplementary Figure 2 CL conditions for regulation of mitochondrial morphology and quality control.
(a) Comparison of the number, length, and unsaturation of acyl chains of CL used in this study. (b) GTP hydrolysis activity of the deletion mutant (Δ581–941) of L-OPA1 (n = 3 independent measurements, data are mean ± s.d.). (c–g) KD of CLS1 using siRNA. (c) RNAs were analyzed by RT-PCR (the experiment was performed one time). Nuclear genome-encoded transcripts were analyzed. (d) Fluorescence images of the control and CLS1 KD cells (representative images of more than 3 independent experiments with similar results). Scale bar: 10 μm. (e) Immunoblot analysis of mitochondrial protein levels using the indicated antibodies (the experiment was performed one time). (f,g) Mitochondrial fusion in CLS1 KD cells and OPA1 KD cells. Mitochondrial fusion was analyzed using HeLa cells expressing mitGFP and mitRFP that were co-cultured and fused with the HVJ envelope. After 6 h culture, fusion efficiency was analyzed by fluorescence microscopy. n, number of cell hybrids analyzed from 3 independent experiments: cont × cont (n = 38), CLS1KD × CLS1KD (n = 41), and OPA1KD × OPA1KD (n = 32). Scale bar: 10 μm. Statistics source data are available in Supplementary Table 1. Unprocessed original scans of agarose gel and blots are shown in Supplementary Fig. 3. (h) Model of the isolation of damaged mitochondria from the active mitochondrial network. OPA1 inactivation and CL remodeling are used as markers for the specific selection of damaged mitochondria.
Supplementary information
Supplementary Information
Supplementary Information (PDF 1755 kb)
Supplementary Table 1
Supplementary Information (XLSX 57 kb)
Rights and permissions
About this article

Cite this article
Ban, T., Ishihara, T., Kohno, H. et al. Molecular basis of selective mitochondrial fusion by heterotypic action between OPA1 and cardiolipin. Nat Cell Biol 19, 856–863 (2017). https://doi.org/10.1038/ncb3560
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/ncb3560
This article is cited by
-
Stay in touch with the endoplasmic reticulum
Science China Life Sciences (2024)
-
FGF19 increases mitochondrial biogenesis and fusion in chondrocytes via the AMPKα-p38/MAPK pathway
Cell Communication and Signaling (2023)
-
Mitochondrial heterogeneity in diseases
Signal Transduction and Targeted Therapy (2023)
-
SynDLP is a dynamin-like protein of Synechocystis sp. PCC 6803 with eukaryotic features
Nature Communications (2023)
-
Structural mechanism of mitochondrial membrane remodelling by human OPA1
Nature (2023)
