
Abstract
Mammalian Kruppel-like transcription factors are implicated in regulating terminal differentiation of several tissue types1,2,3. Deficiency in Kruppel-like factor (KLF) 2 (also known as LKLF) leads to a massive loss of the peripheral T-cell pool4, suggesting KLF2 regulates T-cell quiescence and survival4,5,6,7. Here we show, however, that KLF2 is essential for T-cell trafficking. KLF2-deficient (Klf2-/-) thymocytes show impaired expression of several receptors required for thymocyte emigration and peripheral trafficking, including the sphingosine-1-phosphate (S1P) receptor S1P1, CD62L and β7 integrin. Furthermore, KLF2 both binds and transactivates the promoter for S1P1—a receptor that is critical for thymocyte egress and recirculation through peripheral lymphoid organs. Our findings suggest that KLF2 serves to license mature T cells for trafficking from the thymus and recirculation through secondary lymphoid tissues.
Similar content being viewed by others
Main
Kruppel-like factors (KLFs) are a family of zinc-finger transcription factors including at least 15 mammalian family members2. KLFs have critical roles in the development of specific cell lineages, as demonstrated by the profound phenotypes that result from gene-targeting KLF family members1,2. Several KLFs have an important role in cell maturation—a feature exemplified by erythroid KLF (KLF1), which is essential for erythrocyte production of adult-form haemoglobin1,2,3.
KLF2 is expressed in lung, endothelial cells and lymphocytes4,8,9,10,11, and is essential for normal blood-vessel integrity and lung development4,9,10,11. Although Klf2-/- thymocyte development is grossly normal, few KLF2-deficient T cells are found in peripheral lymphoid tissues4,6. Moreover, the few Klf2-/- T cells that are present in peripheral tissues show signs of activation and induction of cell death, suggesting KLF2 may have a critical role in T-cell quiescence and survival4,6. KLF2 is normally expressed in mature thymocytes, naive T cells and memory T cells, but its expression is dramatically downregulated with T-cell receptor activation4,12,13,14. Overexpression of KLF2 in the Jurkat T cell line leads to inhibition of cell-cycle progression, an effect that may involve KLF2 repression of c-myc transcription and/or induction of p21WAF1/CIP1 (refs 6, 15, 16). Taken together, these features have lead to the proposal that KLF2 functions to prevent spontaneous activation and subsequent death of mature T cells4,5,6,7.
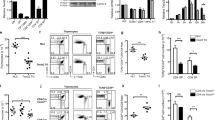
To test this model, we studied maintenance of Klf2-/- T cells in vivo. Because murine KLF2 deficiency is embryonic lethal9,11, we generated fetal liver chimaeras (FLCs) using embryonic day 12.5 fetal livers from Klf2-/- or Klf2+/- donors injected into irradiated Rag2-/- hosts (see Methods). After allowing haematopoietic reconstitution, we studied the T-cell pool in the thymus and peripheral lymphoid tissues of the chimaeras. Consistent with previous reports, which involved a distinct targeted allele of KLF2 (ref. 4), we found that thymic development of Klf2-/- T cells was grossly normal (Fig. 1a, b), but that there was a massive deficit of peripheral T cells (Fig. 1a, c). Notably, we also observed an increase in the representation of Klf2-/- mature CD4+CD8- (CD4 single-positive; CD4 SP) and CD4-CD8+ (CD8 single-positive; CD8 SP) thymocytes compared with controls (Fig. 1b). Similar results were obtained using FLCs generated in lethally irradiated C57BL/6 (rather than Rag2-/-) hosts (data not shown).

Fetal liver chimaeras (FLCs) were generated from Klf2-/- and Klf2+ (Klf2+/- or Klf2+/+) donors in Rag2-/- hosts. a, Representative CD4/CD8 staining of the indicated tissues from Klf2-/- and Klf2+ FLCs. Values represent the percentage of cells in each boxed region. b, c, Cell numbers for the indicated populations from thymus (b) and peripheral lymphoid tissues (c). Graphs show the average recovery from multiple experiments (n > 6 for b; n > 4 for c), with error bars representing standard deviation. Statistical comparisons between Klf2-/- and Klf2+ groups are indicated.
To study the proposed demise of mature Klf2-/- T cells, we performed adoptive transfer of Klf2-/- versus Klf2+/- thymocytes into congenic hosts and tracked maintenance of the donor population. Unexpectedly, donor Klf2-/- CD4 SP cells were recovered at similar total numbers compared to Klf2+/- CD4 SP cells 14 days following adoptive transfer (Fig. 2a, b), and even at later time points (30 days post-transfer; data not shown)—results that seemed to conflict with the model that KLF2 was required for mature T-cell survival in the periphery. We considered that KLF2 deficiency might deregulate T-cell cytokine dependence, such that the normal survival cytokines were superfluous for Klf2-/- T-cell persistence. However, in vitro culture of Klf2-/- and Klf2+/- T cells showed very similar survival characteristics, including dependence on interleukin (IL)-7 for maintenance (Supplementary Fig. S1). As KLF2 deficiency has also been proposed to induce a loss in T-cell quiescence4,6, it was possible that Klf2-/- T cells underwent spontaneous proliferation in vivo, balancing an increased rate of cell death. To explore this, we labelled proliferating cells in vivo with BrdU (5-bromodeoxyuridine). Both Klf2-/- and Klf2+/- T cells showed similar BrdU incorporation following adoptive transfer, arguing against differential proliferation of these populations (Fig. 2c). Although these data suggested Klf2-/- CD4 SP thymocytes were competent for short-term survival, we noted marked abnormalities in their tissue distribution: Whereas Klf2+/- donor T cells were found in blood, lymph nodes and spleen, Klf2-/- T cells segregated almost exclusively to the spleen (Fig. 2a, b), a pattern that was also seen at three and seven days following adoptive transfer (Supplementary Fig. S2a; data not shown). Recovery of both CD4+ and CD8+ donor Klf2-/- T cells was similar to controls soon after adoptive transfer (Supplementary Fig. S2a), but we did observe reduced recovery of Klf2-/- CD8+ (but not CD4+) T cells at later times (days 14 and 30; data not shown). However, it is currently unclear whether KLF2 has a direct role in long-term CD8+ T-cell survival, or if this gradual decline is secondary to altered T-cell trafficking.

a, b, Thymocytes from Klf2+/- or Klf2-/- FLCs were transferred into C57BL/6.SJL hosts and labelled with BrdU for 14 days. The percentage (a) and absolute numbers (b) of donor CD4+ T cells in the indicated tissues was determined (representative of n > 4 experiments). c, Representative BrdU incorporation in donor-derived CD4+ splenocytes. An allophycocyanin (APC)-conjugated anti-BrdU antibody was used to detect BrdU in cells. d, The schematic describes the generation and intrathymic injection of mixed bone-marrow chimaeras. Fluorescence-activated cell sorting (FACS) plots show Klf2-/- and wild-type biotinylated CD4+ cells from a representative mixed chimaera. The bar graph indicates the ratio of FLC-derived:normal donor cells in indicated tissues, normalized against the thymic ratio.
These results suggested KLF2 regulates T-cell trafficking, raising the possibility that the absence of Klf2-/- peripheral T cells might arise from defective thymocyte emigration. To test this model directly, Klf2-/- or Klf2+/- FLCs were used to generate secondary radiation bone-marrow chimaeras, in which donor FLC cells were placed in competition with congenic wild-type cells (Fig. 2d). Biotin was then administered by intrathymic injection and, 36 h later, the export of biotin-labelled thymocytes into peripheral lymphoid tissues was determined. Klf2+/- recent thymic emigrants were detected in blood, spleen and lymph nodes in the ratios expected from thymocyte labelling (Fig. 2d). In contrast, Klf2-/- recent thymic emigrants were very rare, despite efficient labelling of these cells in the thymus (Fig. 2d). These data contrast with those expected if KLF2 deficiency caused T-cell death following thymic egress, in which case most peripheral Klf2-/- T cells would presumably be thymic emigrants. Hence, these results argue for impaired thymic emigration of Klf2-/- T cells.
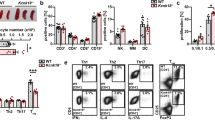
To understand the basis for these altered trafficking patterns, we analysed the phenotype of Klf2-/- T cells. CD4+CD8+ (double-positive; DP) thymocytes from Klf2-/- and Klf2+/- chimaeras had very similar phenotypes (Fig. 3a), consistent with the fact that KLF2 is upregulated only after positive selection in the thymus4,17. Single-positive (SP) thymocytes from Klf2-/- and Klf2+/- chimaeras showed similar expression of some markers, including T-cell receptor β (TCRβ), CD25 and CD5 (Fig. 3a; data not shown), but other molecules were altered in KLF2-deficient cells. Klf2-/- SP thymocytes were CD69high and CD62Llow (Fig. 3a), suggestive of a ‘semi-mature’ phenotype18,19,20; yet these cells were also CD24low and Qa2high (Fig. 3a; data not shown), a phenotype consistent with full maturity18,19,20. Despite their CD69 expression, the CD24lowCD25low phenotype of Klf2-/- T cells argues against them being activated. Furthermore, expression levels of the activation/memory marker CD44 were similar on Klf2-/- and Klf2+/- thymocytes, although there was considerable variability in CD44 expression in individual FLCs (data not shown). With regard to trafficking, the expression of CD62L, CCR7 and β7 integrin—all of which are involved in the entry of T cells into peripheral lymphoid tissues21,22,23,24—was decreased on Klf2-/- T cells (Fig. 3a). Furthermore, CD69 expression by Klf2-/- SP thymocytes is potentially relevant, as CD69 can impair T-cell migration19,25,26. The rare peripheral Klf2-/- T cells were phenotypically similar to SP thymocytes (Fig. 3a; data not shown), and the CD69hiCD62Llo phenotype of Klf2-/- SP thymocytes was maintained at 6 (Supplementary Fig. S2b), 14 or 30 (data not shown) days after adoptive transfer, indicating that the deregulated expression of these markers by Klf2-/- T cells is stable.

a, Phenotype of thymocytes and CD4+ splenocytes from Klf2-/- and Klf2+/- FLCs (representative of n > 4 experiments). b, S1P1, CD62L, β7 integrin and BCL2 mRNA abundance was assessed by real-time RT–PCR analysis of sorted DP and CD4 SP thymocytes from Klf2-/- and Klf2+/- FLCs. Bar graphs show the average signal from n≧3 experiments, with error bars representing standard deviation. P-values from comparisons between Klf2-/- and Klf2+/- CD4 SP samples are indicated.
In order to further explore the maturation state and trafficking potential of Klf2-/- SP thymocytes, we sorted Klf2-/- and Klf2+/- DP and CD4 SP thymocytes and performed real-time polymerase chain reaction following reverse transcription (RT–PCR) on genes known to change in expression during thymocyte maturation (Supplementary Fig. S3). Most genes showed a similar pattern of expression in Klf2+/- and Klf2-/- thymocyte populations, consistent with normal thymocyte maturation in the absence of KLF2 (Supplementary Fig. S3). Real-time RT–PCR analysis did, however, demonstrate a dramatic decrease in the expression of mRNA encoding CD62L in Klf2-/- SP thymocytes (Fig. 3b and Supplementary Fig. S3) and a more modest impairment in the expression of β7 integrin mRNA (Fig. 3b and Supplementary Fig. S3). Notably, loss of KLF2 had no substantial effect on the expression of CCR7 mRNA, although expression of the CCR7 protein was reproducibly reduced on Klf2-/- SP thymocytes (Fig. 3a). Also of note, two molecules involved in the regulation of apoptosis—Bcl2 and Ian1—were unaffected at the mRNA level by KLF2 loss (Fig. 3b and Supplementary Fig. S3). Although these assays focused on CD4 SP thymocytes, preliminary data indicate similar gene expression patterns in Klf2-/- CD8 SP cells (data not shown).
Reduced expression of CD62L, CCR7 and β7 integrin immediately suggested a basis for the altered trafficking of Klf2-/- thymocytes: CD62L and CCR7 are essential for trafficking into peripheral lymph nodes; β7 integrin for entry into Peyer's patches; and access to mesenteric lymph nodes requires either CD62L or β7 integrin21,22,23,24. However, although the loss of these molecules could explain aspects of altered Klf2-/- T-cell trafficking in the adoptive transfer experiments (Fig. 2a, b), they were insufficient to account for the defect in Klf2-/- thymocyte emigration (Fig. 2d). Recently, reports have demonstrated that the sphingosine-1-phosphate receptor S1P1 (also known as Edg1) is critical for thymocyte emigration and T-cell recirculation26,27,28. Hence, we examined the expression of S1P1 in Klf2-/- thymocytes. Real-time RT–PCR analysis demonstrated the expected upregulation of S1P1 mRNA expression between the DP and CD4 SP stages in control thymocytes, but S1P1 expression was not upregulated in Klf2-/- CD4 SP thymocytes (Fig. 3b), nor in Klf2-/- CD8 SP thymocytes (data not shown). On the basis of previous studies27,28,29, defective S1P1 expression could explain the crippled Klf2-/- thymocyte egress observed (Fig. 2d), as well as the absence of Klf2-/- T cells in peripheral blood following adoptive transfer (Fig. 2a, b).
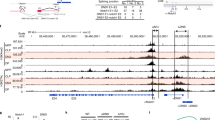
These effects on gene transcription might indicate direct regulation by KLF2 or an indirect consequence of KLF2 deficiency altering thymocyte maturation. Given the primacy of S1P1 in directing thymocyte egress, we focused on the ability of KLF2 to interact with the S1P1 promoter. Chromatin immunoprecipitation (ChIP) assays were performed using an inducible KLF2 expression system6,15 as described previously14, and resulted in co-immunoprecipitation of KLF2 with the proximal S1P1 promoter region (Fig. 4a). Furthermore, reporter assays demonstrated transactivation of the S1P1 promoter by KLF2 (Fig. 4b), arguing that KLF2 positively regulates S1P1 expression. These findings are consistent with previous reports showing upregulation of S1P1 mRNA with KLF2 overexpression16. Interestingly, a consensus KLF family binding-site motif (CACCC) is found just upstream of the S1P1 transcriptional start site (Supplementary Fig. S4), although KLF2 could also be acting through neighbouring SP1 sites, as demonstrated previously15. Together with our analysis of thymocyte gene expression, these findings suggest KLF2 directly induces S1P1 expression in developing thymocytes.

a, Jurkat T cells were induced (“ + Dox”) or not (“ - Dox”) to express HA-tagged KLF2 and chromatin immunoprecipitation (ChIP) performed. PCR for S1P1 promoter fragments was conducted on chromosomal DNA (“Input”), DNA co-precipitated with KLF2 (“HA”), and control immunoprecipitations, as indicated. Primer set 1 amplifies a region immediately upstream of the S1P1 transcriptional start site (Supplementary Fig. S4), whereas primer set 2 amplifies a region about 300 bp further upstream. b, KLF2 transactivation of the S1P1 promoter was assessed by reporter assay14,15. Jurkat (T) and HepG2 (liver) cells were co-transfected with either Klf2 cDNA (“Klf2”) or empty vector (“pcDNA”), plus either the S1P1 promoter luciferase reporter (“S1P1luc”) or control plasmid (“pGL3”). Data are representative of n = 3 experiments. Error bars represent s.d.
In summary, our data suggest a radically different model of the role of KLF2: rather than primarily influencing T-cell survival, KLF2 regulates thymocyte and T-cell trafficking. Loss of KLF2 results in defective expression of S1P1, CD62L, β7 integrin and CCR7, and KLF2 seems to have a direct role in induction of S1P1 expression. KLF2 and S1P1 show similar expression patterns in T cells, both being upregulated on thymocyte maturation, downregulated after T-cell activation and re-expressed in the late effector/memory pool4,12,13,27. Furthermore, KLF2-null and S1P1-null mice die at similar stages in gestation because of widespread haemorrhaging, probably owing to defective tunica media integrity9,11,27,30. It is interesting to speculate that KLF2 may be required for promoting S1P1 expression in endothelial cells, explaining the early embryonic lethality of KLF2-deficient mice.
Our data do not exclude additional roles for KLF2, potentially including long-term effects on T-cell quiescence or survival. Indeed, forced KLF2 expression can clearly promote cell-cycle withdrawal in tumour cell lines6,15,16. However, our findings suggest KLF2 is critical to license mature thymocytes for trafficking competence. Such a role is consistent with the action of other KLF family members, which are essential for terminal differentiation of various cell types1,2,3.
Methods
A detailed description of all materials and methods can be found in Supplementary Information.
Mice, chimaeras and adoptive transfer
Fetal livers from embryonic day 12.5 Klf2-/- and control animals were used to reconstitute irradiated Rag2-/- hosts, which were analysed after eight weeks. In some experiments, thymocytes from FLCs were adoptively transferred into C57BL/6.SJL hosts, which were then offered BrdU-laced drinking water. Donor-derived (CD45.2+) cells were analysed for tissue distribution, phenotype and BrdU incorporation by flow cytometry, using the indicated antibodies (BD Pharmingen). Secondary bone-marrow chimaeras were generated using a mixture of bone marrow from FLCs (CD45.2) plus C57BL/6.SJL (CD45.1) animals to reconstitute lethally irradiated C57BL/6.SJL hosts. After eight weeks, these animals were intrathymically injected with biotin and the appearance of FLC-donor-derived (CD45.2) and wild-type-donor-derived (CD45.1) biotin-labelled cells in the thymus and periphery monitored.
Real-time RT–PCR
Thymocyte subsets were purified by fluorescence-activated cell sorting (FACS) on a FACSVantage (Becton Dickinson), followed by RNA isolation and preparation of cDNA. Real-time RT–PCR was performed on a SmartCycler (Cepheid) using primers listed in Supplementary Information. mRNA abundance was determined relative to controls (Hprt, Gapdh and/or Ctnnb1).
Chromatin immunoprecipitation and gene reporter assays
Jurkat T cells transfected with tetracycline (tet)-inducible haemagglutinin (HA)-tagged KLF2 (refs 14, 15) were cultured with or without doxycycline for 48 h, formaldehyde-fixed, lysed, and subjected to immunoprecipitation with an anti-HA antibody or control IgG. Co-precipitated DNA was used as template for PCR of the S1P1 promoter using primer sets 1 or 2 (initiating 141 base pairs (bp) and 474 bp upstream of the indicated S1P1 transcriptional start site, respectively). For S1P1 reporter assays, normal Jurkat or HepG2 cells were transiently transfected with the indicated plasmids and luciferase activity monitored at 48 h using a Monolight 3010 Luminometer (BD Biosciences).
Statistical analysis
An unpaired two-tailed Student's t-test was applied using GraphPad software (Prism) on normal or log10-transformed data sets.
References
Dang, D. T., Pevsner, J. & Yang, V. W. The biology of the mammalian Kruppel-like family of transcription factors. Int. J. Biochem. Cell Biol. 32, 1103–1121 (2000)
Kaczynski, J., Cook, T. & Urrutia, R. Sp1- and Kruppel-like transcription factors. Genome Biol. 4, 206 (2003)
Turner, J. & Crossley, M. Mammalian Kruppel-like transcription factors: more than just a pretty finger. Trends Biochem. Sci. 24, 236–240 (1999)
Kuo, C. T., Veselits, M. L. & Leiden, J. M. LKLF: a transcriptional regulator of single-positive T cell quiescence and survival. Science 277, 1986–1990 (1997)
Kuo, C. T. & Leiden, J. M. Transcriptional regulation of T lymphocyte development and function. Annu. Rev. Immunol. 17, 149–187 (1999)
Buckley, A. F., Kuo, C. T. & Leiden, J. M. Transcription factor LKLF is sufficient to program T cell quiescence via a c-Myc-dependent pathway. Nature Immunol. 2, 698–704 (2001)
Di Santo, J. P. Lung Kruppel-like factor: a quintessential player in T cell quiescence. Nature Immunol. 2, 667–668 (2001)
Anderson, K. P., Kern, C. B., Crable, S. C. & Lingrel, J. B. Isolation of a gene encoding a functional zinc finger protein homologous to erythroid Kruppel-like factor: identification of a new multigene family. Mol. Cell. Biol. 15, 5957–5965 (1995)
Wani, M. A., Means, R. T. Jr & Lingrel, J. B. Loss of LKLF function results in embryonic lethality in mice. Transgenic Res. 7, 229–238 (1998)
Wani, M. A., Wert, S. E. & Lingrel, J. B. Lung Kruppel-like factor, a zinc finger transcription factor, is essential for normal lung development. J. Biol. Chem. 274, 21180–21185 (1999)
Kuo, C. T. et al. The LKLF transcription factor is required for normal tunica media formation and blood vessel stabilization during murine embryogenesis. Genes Dev. 11, 2996–3006 (1997)
Schober, S. L. et al. Expression of the transcription factor lung Kruppel-like factor is regulated by cytokines and correlates with survival of memory T cells in vitro and in vivo. J. Immunol. 163, 3662–3667 (1999)
Grayson, J. M., Murali-Krishna, K., Altman, J. D. & Ahmed, R. Gene expression in antigen-specific CD8+ T cells during viral infection. J. Immunol. 166, 795–799 (2001)
Wu, J. & Lingrel, J. B. Kruppel-like factor 2, a novel immediate-early transcriptional factor, regulates IL-2 expression in T lymphocyte activation. J. Immunol. 175, 3060–3066 (2005)
Wu, J. & Lingrel, J. B. KLF2 inhibits Jurkat T leukemia cell growth via upregulation of cyclin-dependent kinase inhibitor p21WAF1/CIP1. Oncogene 23, 8088–8096 (2004)
Haaland, R. E., Yu, W. & Rice, A. P. Identification of LKLF-regulated genes in quiescent CD4+ T lymphocytes. Mol. Immunol. 42, 627–641 (2005)
Mick, V. E., Starr, T. K., McCaughtry, T. M., McNeil, L. K. & Hogquist, K. A. The regulated expression of a diverse set of genes during thymocyte positive selection in vivo. J. Immunol. 173, 5434–5444 (2004)
Gabor, M. J., Godfrey, D. I. & Scollay, R. Recent thymic emigrants are distinct from most medullary thymocytes. Eur. J. Immunol. 27, 2010–2015 (1997)
Feng, C. et al. A potential role for CD69 in thymocyte emigration. Int. Immunol. 14, 535–544 (2002)
Boursalian, T. E., Golob, J., Soper, D. M., Cooper, C. J. & Fink, P. J. Continued maturation of thymic emigrants in the periphery. Nature Immunol. 5, 418–425 (2004)
Arbones, M. L. et al. Lymphocyte homing and leukocyte rolling and migration are impaired in L-selectin-deficient mice. Immunity 1, 247–260 (1994)
Wagner, N. et al. Critical role for β7 integrins in formation of the gut-associated lymphoid tissue. Nature 382, 366–370 (1996)
Wagner, N. et al. L-selectin and β7 integrin synergistically mediate lymphocyte migration to mesenteric lymph nodes. Eur. J. Immunol. 28, 3832–3839 (1998)
Cyster, J. G. Chemokines, sphingosine-1-phosphate, and cell migration in secondary lymphoid organs. Annu. Rev. Immunol. 23, 127–159 (2004)
Nakayama, T. et al. The generation of mature, single-positive thymocytes in vivo is dysregulated by CD69 blockade or overexpression. J. Immunol. 168, 87–94 (2002)
Shiow, L. R. et al. CD69 acts downstream of interferon-α/β to inhibit S1P1 and lymphocyte egress from lymphoid organs. Nature 440, 540–544 (2006)
Matloubian, M. et al. Lymphocyte egress from thymus and peripheral lymphoid organs is dependent on S1P receptor 1. Nature 427, 355–360 (2004)
Allende, M. L., Dreier, J. L., Mandala, S. & Proia, R. L. Expression of the sphingosine-1-phosphate receptor, S1P1, on T-cells controls thymic emigration. J. Biol. Chem. 279, 15396–15401 (2004)
Lo, C. G., Xu, Y., Proia, R. L. & Cyster, J. G. Cyclical modulation of sphingosine-1-phosphate receptor 1 surface expression during lymphocyte recirculation and relationship to lymphoid organ transit. J. Exp. Med. 201, 291–301 (2005)
Liu, Y. et al. Edg-1, the G protein-coupled receptor for sphingosine-1-phosphate, is essential for vascular maturation. J. Clin. Invest. 106, 951–961 (2000)
Acknowledgements
We thank J. Cyster for generous provision of CCL19–Fc fusion protein, Y. Li for developing S1P1 RT–PCR assays, M. Jenkins and M. Mescher for critical input on the manuscript, and members of the ‘Jamequist’ laboratory for helpful discussions. This work was supported by the NIH (an immunology pre-doctoral training grant to B.T.E., a grant to K.A.H., and a grant to S.C.J.), the American Cancer Society (a grant to S.C.J.), and the Cancer Research Institute (post-doctoral fellowship to C.M.C.).
Author information
Authors and Affiliations
Corresponding authors
Ethics declarations
Competing interests
Reprints and permissions information is available at npg.nature.com/reprintsandpermissions. The authors declare no competing financial interests.
Supplementary information
Supplementary Notes
This file contains Supplementary Methods with eight references, details of grant support and Supplementary Figures S1–S4 with legends. Supplementary Figure S1 details the normal in vitro survival of KLF2-/- mature thymocytes. Supplementary Figure S2 shows aberrant trafficking and phenotype of KLF2-/- SP thymocytes after adoptive transfer. Supplementary Figure S3 details real-time PCR analysis of gene expression in KLF2-/- and KLF2+/- DP and CD4 SP thymocytes. Supplementary Figure S4 shows the sequence of human and mouse S1P1 promoters, showing the CACCC sequence and the location of primers used for the CHiP assay. (PDF 453 kb)
Rights and permissions
About this article
Cite this article
Carlson, C., Endrizzi, B., Wu, J. et al. Kruppel-like factor 2 regulates thymocyte and T-cell migration. Nature 442, 299–302 (2006). https://doi.org/10.1038/nature04882
Received:
Accepted:
Issue Date:
DOI: https://doi.org/10.1038/nature04882
This article is cited by
-
FOXP1 and KLF2 reciprocally regulate checkpoints of stem-like to effector transition in CAR T cells
Nature Immunology (2024)
-
A genome-wide in vivo CRISPR screen identifies essential regulators of T cell migration to the CNS in a multiple sclerosis model
Nature Neuroscience (2023)
-
How autoreactive thymocytes differentiate into regulatory versus effector CD4+ T cells after avoiding clonal deletion
Nature Immunology (2023)
-
Tissue-resident immune cells in the pathogenesis of multiple sclerosis
Inflammation Research (2023)
-
Krüppel-like factor 2 controls IgA plasma cell compartmentalization and IgA responses
Mucosal Immunology (2022)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
