

Abstract
Understanding how the brain functions differently as one learns to read may shed light on the controversial nature of the reading ability of human being. Logographic writing system such as Chinese has been found to rely on specialized neural substrates beyond the reading network of alphabetic languages. The ability to read in Chinese has also been proposed to rely on writing skills. However, it was unclear whether the learning-related alteration of neural responses was language specific or resulted from the more reliance on writing practice during acquisition. This study investigated whether the emergence of typical logographic-specific regions relied on learning by writing. We taught proficient alphabetic language readers Chinese characters and used pre-test and post-test to identify changes in two critical stages of reading, namely, orthographic processing and orthographic-to-phonological mapping. Two typical left hemispheric areas for logographic reading showed increased responses to characters in the brains of proficient alphabetic readers after learning, regardless of whether the learning strategy involved writing practice. Moreover, learning strategy modulated the response magnitude or multivoxel patterns in the left superior parietal lobule, left middle frontal gyrus, and right fusiform gyrus, some of which were task dependent. The findings corroborated a limited role of writing in the emergence of logographic-specific reading network and suggested the heterogeneous nature of different brain regions in this network.
Significance Statement
There has been debate on whether the development of skills for reading logographic characters depends on the skill of writing. We examined the emergence of typical Chinese-reading neural substrates when learners were taught character with and without training on writing. Behavioral and neural functional alterations were identified after proficient alphabetic readers learned to read Chinese with or without training on writing. Altering the responses in the left superior parietal lobule and middle frontal gyrus to Chinese characters did not require a writing-based learning, but writing modulated the responses in these areas.
Introduction
Learning to read is one of the most remarkable capabilities of human being, the nature of which remains controversial. On the one hand, as a recent invention and a commonly mastered skill bound to language faculty, reading is proposed to be derived from preexisting functions and thus have universal neurobiological basis regardless of writing system (Paulesu et al., 2000; Dehaene et al., 2001, 2010; Cohen et al., 2002; Bolger et al., 2005; Dehaene and Cohen, 2007; Nakamura et al., 2012; Rueckl et al., 2015; Feng et al., 2020; Verhoeven and Perfetti, 2021). On the other hand, writing system is a product of culture and literacy acquisition is a product of education. Culture-specific views posit that reading is attuned to the characteristics of the specific languages (Tan et al., 2001, 2005a; Kuo et al., 2004; Siok et al., 2004, 2009; Hu et al., 2010).
Comparison between Chinese and alphabetic language reading has been a major arena for the debate. The writing system of Chinese is largely logographic: the orthography does not imply pronunciation. Chinese readers cannot rely on rules of orthographic-phonological mapping to decode the sound of a written word like one might do in alphabetic language reading. The critical role of phonological awareness in English reading development has been well documented (Wagner et al., 1997; Eden and Moats, 2002; Temple et al., 2003; Vellutino et al., 2004; Ziegler and Goswami, 2005; McArthur et al., 2018), whereas the reading development of Chinese as first or second language has been found to benefit much less from phonological skills (Meade, 2020) but more from the writing ability and orthographic awareness (Tan et al., 2005b; Ye et al., 2021; Wong and Zhou, 2022). Meta-analyses have consistently recognized the roles of several regions in orthographic processing and orthographic-phonological mapping during Chinese reading, including intraparietal sulcus/superior parietal lobule (IPS/SPL; Tan et al., 2005a; C.Y. Wu et al., 2012), left middle frontal gyrus (MFG; Bolger et al., 2005; Tan et al., 2005a; C.Y. Wu et al., 2012), and right ventral occipital-temporal areas (Bolger et al., 2005). Orthographic transparency affects the between-language similarity in brain activation patterns (Kim et al., 2016; Dong et al., 2021). For readers who are proficient at alphabetic language, successful learning of Chinese characters activates typical logographic processing areas (Y. Liu et al., 2007a; Nelson et al., 2009).
Regarding the original question on the nature of reading, the parallel cognitive and neural evidence of between-language difference does not address whether the neural activational differences result from the differences in the learning/processing strategy or the differences in the writing system itself. The covariant learning hypothesis proposes that the neural substrates for processing certain kind of stimulus are developed by associating the stimulus with the cognitive and learning processes (Kochunov et al., 2003; Tan et al., 2005a). The form of writing system affects the learning strategy, which affects the functional neuroanatomy of reading (Tan et al., 2005a). Thus, comparing how different learning strategies affect the processing of the same language is an effective approach to dissociating the effect of learning from that of the language per se. Several studies have investigated the effects of different learning strategies or cognitive processes on a given language (Y. Liu et al., 2007a; Nakamura et al., 2012; Cao et al., 2013, 2017; Cao and Perfetti, 2016; Lagarrigue et al., 2017). However, these studies have shown a mixed picture on whether the so-called logographic-specific areas are the results of training on writing, which might be because of the diversity in the training procedure, levels of processing, and the second language background of participants. Moreover, interpretation of the learning effect was also difficult in the absence of a prelearning functional neuroimaging measure when participants had no knowledge of the target language.
The present study investigated how the brain functions differently when proficient alphabetic language readers learned a novel logographic system in different strategies. We asked whether learning Chinese characters for a 7-d training elicited spontaneous neural responses in the typical Chinese reading areas and whether the emergence of these regions, particularly the SPL/IPS and MFG, relied on writing tutoring and practicing. Learning effect was examined using a pre-post test paradigm. We randomly assigned participants to two strategy groups and used a passive viewing task to examine the automatic processing of characters. We investigated two critical processes in reading acquisition at the very early stage, namely, character recognition (visual) and orthographic-phonological mapping (visual-auditory modality).
Materials and Methods
Screening of participants on language background and cognitive abilities
Language History Questionnaire (P. Li et al., 2014) was used to screen and recruit participants (1) whose age of first exposure to English was before 6, (2) whose self-evaluated proficiency of English was “very good” or “excellent,” (3) who were native speakers of Germanic or Romantic languages (Haspelmath et al., 2008; Lewis et al., 2014; Skirgård et al., 2017), and (4) whose experience with Chinese was minimal.
To ensure that participants were at very basic levels of Chinese, we presented a list that contained the 150 most-frequent Chinese characters (Cai and Brysbaert, 2010) and the real single-word characters used in the main experiment on a paper to each participant. Participants were asked to mark a character if they knew its pronunciation or meaning. Participants who marked over 10 characters were excluded from further study.
Handedness of participants was measured using a revised version of the Edinburgh Inventory (https://www.brainmapping.org/shared/Edinburgh.php, adapted from Oldfield, 1971). Only right-handed participants were included in further study.
The following tests were applied to ensure participants who were assigned to the two groups of learning strategies were of comparable language and cognitive abilities.
Multilingual Naming Test (MINT; Gollan et al., 2012). Participants were asked to name the black-and-white line-drawings in English one at a time. Accuracy was measured.
English vocabulary test. The test was originally developed for Dutch (Keuleers et al., 2015) and revised for testing English vocabulary (vocabulary.ugent.be/). The vocabulary set consisted of real words in American spelling and pseudowords. One word showed on the screen at a time and participants determined whether they knew this word or not by pressing the key “J” or “F.” The performance was indicated by hit rate − false alarm rate, i.e., the proportion of real words that were correctly recognized minus the proportion of pseudo-words that were mistakenly recognized as known words.
One Minute Reading Test (1MRT; Transvaal Education Department, 1987). Participants were asked to read across a page of English words out loud, from left to right, line by line, carefully but as fast as possible, for 1 min. All the words are one to two syllables.
Rapid Automatic Naming Test (RAN). The subtests of color-naming and digit-naming asked the participant to name the color or digit on the screen as fast as possible, and press any key to proceed to the next trial. Performance was measured by the reaction time on the correct trials.
Matrix span. The test was used to measure the visuospatial working memory (Stone and Towse, 2015). A five-by-five grid was shown on the screen in each trial. Some cells randomly changed color one by one. In the recall phase, participants were presented with the grid again and were asked to click on the cells in the order as they appeared. The number of cells to be remembered increased if the recall was correct. Memory span was the largest number of cells that could be correctly remembered.
Two-back verbal working memory test. In the English version, participants saw one word at a time and decided whether the stimulus in the Nth trial was the same as the one in the N–2 trial. In the Chinese version of the task, the stimulus was individual Chinese characters. The ratio of “same” to “different” trials was 1:3. Performance was measured by accuracy.
Raven test. Participants took the short version of Raven test with 12 questions (Raven, 2017).
Participants
Among the 81 adults from the ECNU and NYU-Shanghai community who volunteered to participate, 43 were excluded after screening: 35 for mismatched language background, seven for being left-handed, and one for early history of dyslexia. Among the 38 who participated in the study, three quit halfway, three fell asleep during fMRI scans, and the data of two were not fully recorded because of technical problems. This resulted in 30 participants in total (14 females) for the following analyses. All the participants were right-handed, age from 18 to 35, reported normal or correct-to-normal vision and normal hearing status, and had no history of neurologic disease or language impairment. Participants were native speakers of Germanic or Romantic languages and were proficient at English. All native English speakers rated their English proficiency as 7/7. For those whose native language was not English, the mean self-reported proficiency at English was 6.4/7 (very proficient or excellent), and the mean proficiency at their native language was 6.7/7. Fifteen participants were bilingual or multilingual, but none had experience with languages other than Germanic or Romantic languages. Their proficiency of Chinese was at very basic level (knew no more than 10 characters). They had started to learn the principles of using pinyin to code pronunciation of Chinese. This study was approved by the University Committee on Human Research Protection of East China Normal University (Approval Number: HR-0502017).
Materials
The characters, pseudo-characters and scrambled characters or a subset of them were used in the learning session, behavioral pre-test and post-test, and the prelearning and postlearning fMRI tasks.
Real characters
One hundred and thirty-two Chinese characters were selected. These characters were of high frequency (Cai and Brysbaert, 2010; Table 1), each denoting to a concrete noun. There was no homophone in the stimuli. The characters were assigned to lists A and B, each containing 66 words. Character in the two lists were balanced on frequency (Cai and Brysbaert, 2010), frequency of their English equivalents (Brysbaert and New, 2009), age of acquisition, imageability (Y. Liu et al., 2007b), and stroke count (all ps > 0.05; Table 1). The same radical never appeared in both lists.
Mean and SDs of real characters
Pseudo-characters
One hundred and thirty-two pseudo-characters were produced based on the real characters used in this study. Radicals of the characters within a list were randomly shuffled and paired with the component of a different character using Truetype. We manually revised the stimulus if the generated one happened to be a real character.
Scrambled characters
Strokes of each pseudo-character was scrambled to create 132 scrambled characters using Truetype. We purposely adjust some scrambled characters to ensure that the structure did not follow orthography principles of Chinese.
Evaluations of pseudo-characters and scrambled characters
An independent group of 23 English native speakers with Chinese-learning experience over one year were recruited to assess the character-ness of pseudo-characters and scrambled characters. The raters saw the stimuli in a randomized order, one at a time, and rated “To what extent do you think this is a Chinese character” on a five-point scale. The mean resemblance score of pseudo-characters was 4.05 (SD = 0.51) and the mean of scrambled characters was 1.04 (SD = 0.08). The mean rating of the pseudo-characters was significantly greater than that of the scrambled characters (one-tailed t = 67.27, p < 0.00001), suggesting good validity of the stimuli.
Auditory stimuli
Characters were read by a Mandarin Chinese native speaker. The recorded audios were equated on loudness, frequency band and bit rate using Adobe Audition. One hundred and thirty-two nonverbal sound was created by reversing the audio of each character. Audios of tones at 500, 600, and 700 Hz were created.
Design and overall procedure
Participants were assigned to one of the two groups of learning strategy, fifteen in each group, after the screening tests. Each participant went through the prelearning behavioral test and fMRI scan on day 1 and went through the postlearning test and scan on day 9 (Fig. 1). On days 2–8, they learned 66 Chinese characters by either a pinyin-based strategy or a pinyin + writing strategy according to the group assignment. According to the screening tests, no significant between-group difference was found in the performance of any cognitive ability test, English vocabulary, MINT, or RAN-digit test (all ps > 0.05). The pinyin group showed higher accuracy in the 1MRT English reading test (t = 2.46, p = 0.02) and shorter reaction time in the RAN-color naming test (t = −2.11, p = 0.04), likely because there were 11 English monolinguals in the pinyin group and only 4 in the other.

A, Paradigms of character learning in two strategy groups. B, Timeline of the study for each participant.
Learning session
Each participant studied all the 66 real characters in one of the real-character lists (Fig. 1). In each strategy group, half of the participants learned list A and the other half learned list B. The learning session lasted for seven consecutive days, including five acquisition phases and two review phases. The first review phase was on day 4 and the second was on day 8. In each acquisition phase, participant received a list of 13 or 14 new characters. Experimenter first went over the pinyin of each character with participants to ensure that they were able to pronounce the characters using pinyin. For the pinyin + writing group, experimenter also taught participants the basics of character writing, including identifying the subcomponent of a character, writing individual strokes, and writing with correct stroke order. Then the participant went over the characters on a program implemented in E-prime 2.0. A character and its English translation were shown on the screen for 1 s while the pronunciation was played once. The next slide presented the character and its pinyin. The pinyin group were asked to spell the pinyin and pronounce the character. The pinyin + writing group were asked to additionally write down the character. The practice slide was learner-paced and repeated for three times. At the end of an acquisition phase and the beginning of a new acquisition phase, participants took a spelling test, during which they wrote down the pinyin of a heard character that was learned in this/the previous phase. The pinyin + group was asked to additionally wrote down the character.
In the two review phases, participants took the spelling and dictation test of same paradigm as those at the end and beginning of an acquisition phase, except that the target characters in the review-phase tests included all the words that had been learned.
Prelearning and postlearning behavioral tests
Character recognition paradigm
On day 1 and day 9, participants performed character recognition tasks in visual, auditory, or visual-auditory modality. In the visual task, participants judged whether they knew the character on the screen. For each participant, materials were the 66 learned real characters and 66 derivative pseudo-characters. In the auditory task, participants judged whether the pronunciation refers to a Chinese word. Stimuli were the pronunciation of learned characters for each participant and the corresponding reversed audios. The auditory task was irrelevant to the aim of the present study and was not considered in further analyses. In the visual-auditory task, participant judged whether the character on the screen matched the auditorily presented speech sound. Stimuli were the 66 learned characters, each paired with either the correct pronunciation or the pronunciation of another character. In each trial, the target stimulus was presented for 600 ms, followed by a 2000-ms blank screen, during which participant responded by pressing buttons. Accuracy was recorded. Trials within each modality were randomized and separated into two blocks with equal number of trials. The presentation order of the blocks was randomized. Participants could take break between blocks.
Behavioral data analysis
Two-way mixed-design ANOVA was performed to examine the effect of learning on accuracy of character recognition test and the effect of strategy on the learning effect. The within-subject factor was the stage of learning (prelearning vs postlearning) and the between-subject factor was the learning strategy (pinyin + writing vs pinyin). The tests were performed on different stimulus modalities separately. Note that we only examined the recognition rate of real characters, because the decision on pseudo-character did not reflect a learning effect.
Prelearning and postlearning task in fMRI
The fMRI task used a block design. Participants read or listened to the learned characters and other stimulus (see above, Materials) and performed a perceptual detection task while being scanned. Because each participant learned only half of the 132 words, characters in the unlearned list were used as the novel characters for the participant. The main experiment implemented a mixed design. The learning strategy (pinyin + writing vs pinyin) was a between-subject variable. The stage of learning (prelearning vs postlearning) and the type of stimuli were the within-subject variables. There were twelve types of stimulus (Fig. 2A): visually presented learned character (Vl), novel character (Vn), pseudo-character (Vp), and scrambled character (Vs); auditorily presented pronunciation of learned character (Al), novel character (An), reversed speech sound of learned word (Ab), and tone (At); learned character and its pronunciation (VA_match), learned character and pronunciation of another word (VA_mis), learned character and the reversed speech sound (VlAb), and pseudo-character and pronunciation of learned word (VpAl). A stimulus trial was formed of a 600 ms stimulus and a 200 ms blank (Fig. 2B). The presentation sequence of the 66 stimulus trials in each condition were pseudo-randomized and grouped into 11 miniblocks, six trials per block. Sequences of the miniblocks of all the conditions were determined using OPTSEQ (https://surfer.nmr.mgh.harvard.edu/optseq/). A fixation cross was presented between miniblocks, the duration of which was jittered, ranging from 1.625 to 6.675 s. Twenty-one out of the 66 fixations were presented in red and randomly distributed through the task. Participants were asked to passively view and listen to the stimulus, and press a button as soon as a red fixation appeared. The task was separated into two runs and took around 20 min in total.
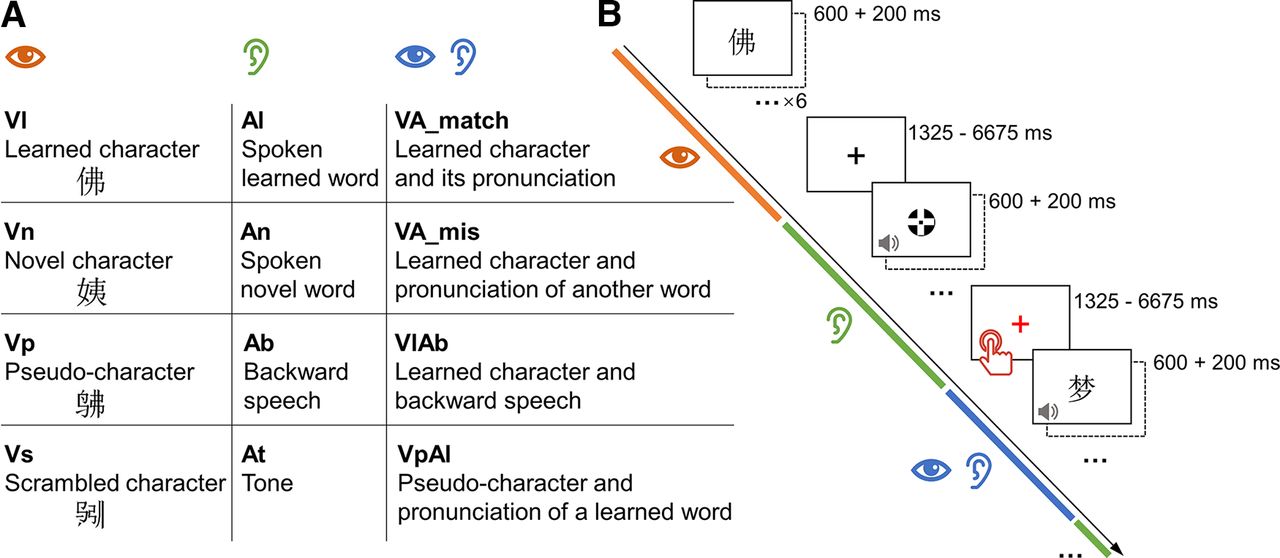
Conditions (A) and paradigm (B) of the fMRI language task. Sequence of the visual (orange), auditory (green), or integrated (blue) miniblocks were randomized. Participants were asked to press the button when a fixation was shown in red.
MRI acquisition
Subjects were scanned in a 3T MRI scanner (Siemens Prisma; Siemens) using a 20-channel head coil. Functional images were acquired using a single-shot T2*-weighted gradient-echo echo planar imaging pulse sequence [TR = 2450 ms, TE = 30 ms, flip angle (FA) = 81°, each volume comprising 40 slices, matrix 64 × 64, field of view (FoV) = 192 mm × 192 mm2, voxel size = 3 × 3 × 3 mm3, interleaved acquisition]. T1-weighted anatomic image was acquired using a multiecho MPRAGE sequence (TR = 2300 ms, TE = 2.32 ms, FA = 8°, matrix 256 × 256, FoV = 240 × 240 mm2, slice thickness = 0.9 mm).
Image preprocessing
Image preprocessing and uni-voxel analyses were performed using SPM8 (Wellcome Centre for Human Neuroimaging, London; https://www.fil.ion.ucl.ac.uk/spm/). The first four volumes of each session were excluded to allow for magnetic saturation. Functional images were corrected for slice timing and head motion, normalized to MNI space using the segmentation-based procedure, smoothed using a Gaussian filter [full-width at half-maximum (FWHM) = 5 mm], and filtered with a 128-s high-pass filter. The moderate kernel size was applied so that the local multivoxel patterns were retained.
Whole-brain uni-voxel analysis
The main effect of learning and the effect of strategy on the learning effect were examined in the two-stage random-effect analyses using general linear model (GLM). Subject-specific responses to each of the 12 types of stimuli were estimated in the prelearning and postlearning scans separately using general linear models, regressors of which were constructed as a boxcar function convolved with the canonical hemodynamic response function. Trials of participant responses and the six rigid-body motion parameters were modeled as covariates.
To examine the effect of learning, the within-subject effect of the post > prelearning contrast was first estimated in the first-level GLM for each participant. Second-level analysis was performed on the contrast images over all participants using one-sample t test against zero. To examine the interaction between strategy and learning stage, specifically, the effect of strategy on the effect of learning, the first-level contrasts of post > prelearning were supplied to the second-level between-group test, where the pinyin versus pinyin + writing groups were compared using independent sample t contrast. Each group-level contrast map was thresholded at a cluster-wise corrected α of 0.05 using the AlphaSim algorithm implemented in NeuroElf (https://neuroelf.net/). Significance of the clusters was determined jointly by the voxel-wise p of 0.05 and the minimum cluster size determined by a 2000-iteration simulation.
Uni-voxel region of interest (ROI) analysis
We performed ROI analyses to further investigate the effect of strategy on learning, specifically, whether involving a writing-based learning strategy will lead to response differences in the regions that have been consistently identified to be specific to Chinese reading. Four coordinate-based a priori regions of interests (ROIs) were selected based on previous meta-analyses on Chinese character reading: left SPL, left MFG, and the right and left FG. The left SPL, left MFG and right FG were considered Chinese-specific and reliably identified in multiple subcomponents of character processing (see Introduction). Because the fusiform ROI was the only ROI in right hemisphere and because it was part of the language-general reading network, we additionally included the left FG as a left hemispheric benchmark to the right ROI. Each ROI was constructed as a 12-mm radius sphere centered at the peak coordinate reported by the meta-analyses. The coordinates of the left MFG ([−46, 18, 28]) and SPL ([−36, −42, 48]) were from Tan et al. (2005a), because this was the meta-analysis that revealed these areas by directly contrasting the Chinese reading against alphabetic reading. The coordinate of the right FG (converted to MNI coordinate from Talairach coordinate [33, −67, −14]) was from Bolger et al. (2005), because this was the meta-analysis that identified the right FG in Chinese reading, and this was the review that proposed the right occipitotemporal cortex was more involved in Chinese reading. While the left FG was a universally identified area for reading, we used the coordinate identified for Chinese character processing ([−32, −54, 6]) in the meta-analysis by Tan et al. (2005a). The first-level contrast of post > prelearning was averaged across voxels within each ROI within participant. The mean signals were compared between two strategy groups over participants using independent sample t contrast. Because reading development of Chinese has been found to behaviorally benefit from writing ability (Tan et al., 2005b; Ye et al., 2021; Wong and Zhou, 2022; see Introduction), and because the selected ROIs have been suggested specific to Chinese reading, we examined whether the additional writing training resulted in increased activations in these regions, by performing the pinyin + writing > pinyin group contrasts.
Multivoxel pattern analysis in a priori ROIs
We performed classification analyses to examine whether the learning strategy affected the multivoxel patterns associated with character processing in each of the four ROIs. Training and testing were performed in a cross-validation procedure that iterated over participants. In each cross-validation fold, all but one participant’s data were used for training and the left-out participant’s data were used for testing. The training exemplars were the voxel patterns of the post > prelearning contrast within each ROI of the training participants. The training label/target was the group membership of each participant, i.e., the learning strategy (pinyin or pinyin + writing). Support vector machine classifiers were trained to learn the neural signatures associated with each of the two strategies. The trained classifiers were applied to predict the group membership of the left-out participant. The mean accuracy over all folds, i.e., participants, indicated whether the neural signatures of learning effect were systematically altered by the learning strategy so that they could be used to predict the learning strategy used by an individual whose data were previously unseen by the model. The significance level of the accuracy was determined by 2000-iteration random permutations, in which all the procedure and data remained the same as the actual analyses, except that the labels of the test exemplars were randomly shuffled within each fold.
Language background classification
The different number of monolingual and bilingual participants in the two groups was a potential confounder to the effect of strategy. Therefore, we performed classification on the language background of participants using exactly the same data of interests as in the main analysis, namely, the a priori ROIs and the clusters identified by the whole-brain analysis on strategy effect. The same procedure of multivoxel pattern analysis of strategy classification was applied, except that the training label/target was the language background of a participant, i.e., being monolingual versus bilingual. If the classification accuracy was not significantly different from the chance level, it suggested that the neural signatures associated with the tasks between monolingual and bilingual participants were not distinguishable, hence, the group difference could not be attributed to the language background. The data and scripts will be shared on request.
Results
Behavioral results on character recognition test
In the visual modality of the character recognition test, ANOVA revealed a main effect of the stage of learning (F(1,28) = 125.55, p = 6.14 × 10−12) on recognition rate. Learning improved the performance from a mean recognition rate of 0.21–0.88 (Fig. 3). No significant effect of learning stage × strategy interaction (F(1,28) = 12.06, p > 0.05) or main effect of strategy (F(1,28) = 1.91, p > 0.05) was found. In the visual-auditory test, ANOVA also revealed a main effect of the stage of learning (F(1,28) = 277.09, p = 4.70 × 10−16) on the performance. Learning improved the recognition rate from a mean of 0.10–0.71 (Fig. 3). No significant effect of learning stage × strategy interaction (F(1,28) = 3.63, p > 0.05) or main effect of strategy (F(1,28) = 0.39, p > 0.05) was found. Thus, different learning strategies did not cause different learning gains in the recognition test. The postlearning recognition rate over all participants in both tasks ranged from 0.20 to 0.98, suggesting that the performance did not reach a theoretical ceiling.

Behavioral results of character recognition rate.
Effects of learning: whole-brain uni-voxel GLM results
Effect of learning on orthographic processing
For visually presented characters that were studied during the learning session (Vl), the main effect of learning (postlearning > prelearning contrast over both strategy groups) was found in wide cortical areas in the bilateral SPL that extended to middle occipital gyrus (MOG), the left inferior gyrus (IFG) that included pars opercularis and pars triangularis and extended to MFG, the supplementary motor areas (SMA), and the right insula-IFG (Fig. 4A; Table 2; cluster-wise corrected p = 0.05, cluster size determined by voxel-wise p of 0.05; df = 29). By contrast, activational difference between postlearning and prelearning scans for the scrambled characters were observed in one cluster at the calcarine-precuneus area (peak coordinate x = 0, y = −61, z = 19; peak Z = 4.49, K = 314 voxels; cluster-wise corrected p = 0.05, cluster size determined by voxel-wise p of 0.05; df = 29).
Regions presenting the main effect of learning
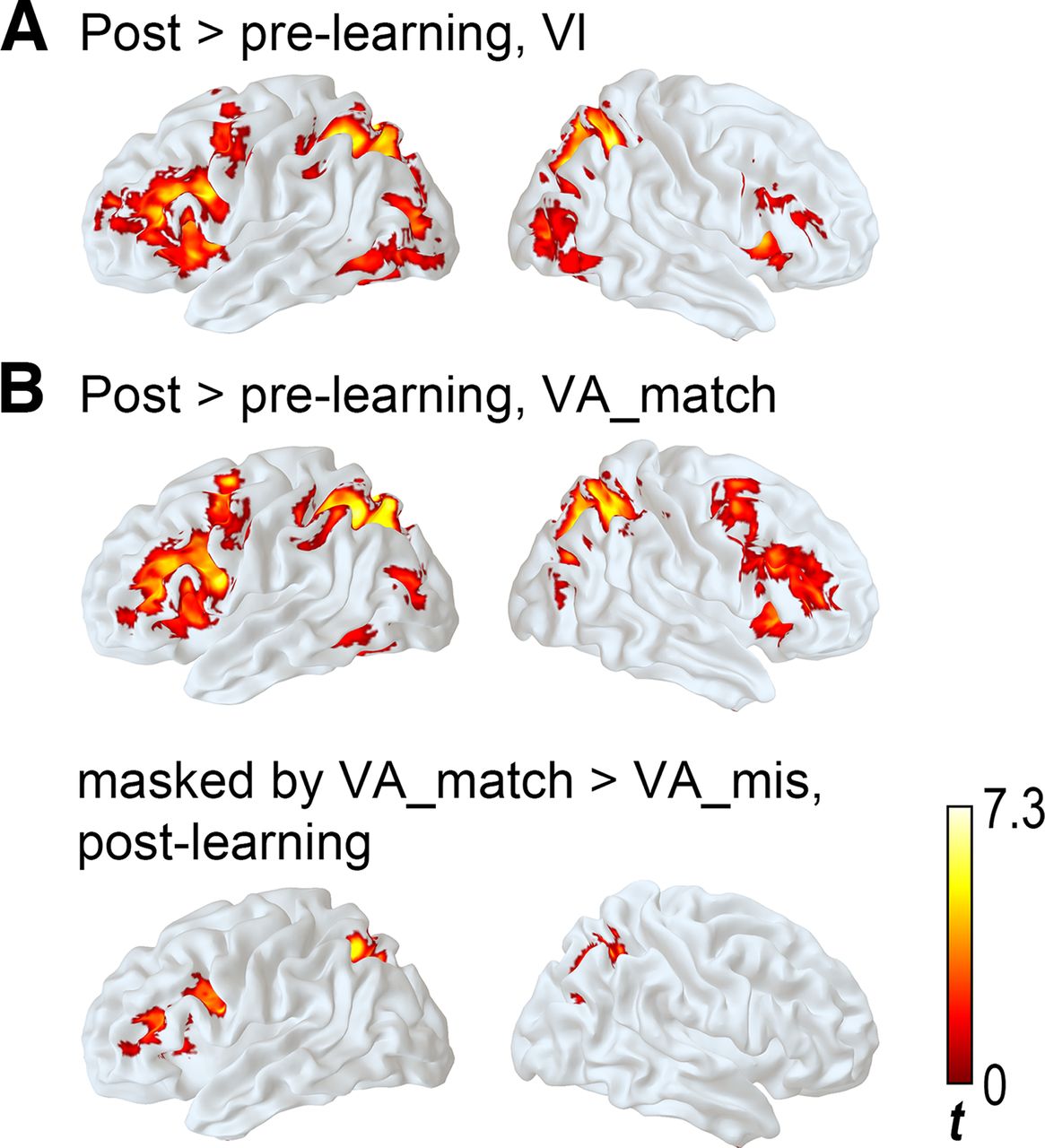
Whole-brain uni-voxel results of learning effect. A, Results of the visual processing. B, Results of visual-auditory processing.
Effect of learning on visual-auditory processing
When the character and its pronunciation were presented simultaneously (VA_match), the post > prelearning effects across strategy groups were found in the left IFG extending to MFG, the bilateral SPL-superior occipital gyri, the left inferior temporal gyrus-FG, the right IFG, and the SMA (Fig. 4B; Table 2; cluster-wise corrected p = 0.05, cluster size determined by voxel-wise p of 0.05; df = 29).
To examine whether the identified areas were sensitive to the correct orthographic-phonological mapping, or whether they just reflected a general effect of multimodal processing, we performed the VA_match versus VA_mis contrast in the postlearning session, thresholded the map using the same cluster-wise corrected p at 0.05, and masked the results of the learning effect with the VA_match versus VA_mis contrast results. Significant effects were identified in the left IFG-MFG and bilateral SPL-MOG (Fig. 4B), suggesting that these areas were sensitive to the correct speech-print association. That is, among the regions showing the main effect of learning, the SMA, left inferior temporal gyrus, and right IFG were not found to respond differently to matched versus mismatched speech-print pairing.
Effects of strategy: whole-brain uni-voxel GLM results
Effect of strategy on orthographic processing
The between-group contrast revealed that in Vl processing, the learning effect for the pinyin + writing group was greater than that for the pinyin group in two adjacent clusters in the right supramarginal gyrus (SMG) and postcentral gyrus (Fig. 5A; Table 3; cluster-wise corrected p = 0.05, cluster size determined by voxel-wise p of 0.01; df = 28). Post hoc analysis revealed that the effects in both clusters were contributed by a decrease in activation after learning (postlearning < prelearning) of the pinyin group (Extended Data Fig. 5-1).
Regions presenting difference between strategy groups (pinyin + writing > pinyin) on the learning effect

Effect of strategy on learning effect. A, Whole-brain uni-voxel GLM results for Vl processing. B, Whole-brain uni-voxel GLM results for the VA_match processing. Post hoc analysis of each cluster was shown in Extended Data Figure 5-1. C, ROI uni-voxel analysis results. The four ROIs defined based on meta-analyses were shown in surface rendering of the brain. Mean and SE of the β estimates for each ROI in each condition were shown in the bar graphs. Markers above a bar indicated the mean was significantly greater than zero according to a one-sample t test. Markers between two bars indicated significant mean difference between groups. ***p < 0.001, *p < 0.05, #p < 0.1. The overlap of the a priori ROIs and results of the whole-brain analysis was shown in Extended Data Figure 5-2. D, Accuracy of classifying participant’s group membership. Note that the critical values of accuracy at p of 0.05 were determined based on random permutation of each set of data independently, but all the critical values turned out to be the same (displayed as a dashed straight line), which was not surprising when the numbers of cross-validation folds and the numbers of test exemplars per fold were the same across all the classification tests. The results of classifying participant’s language background using the same procedure were as shown in Extended Data Figure 5-3.
Extended Data Figure 5-2
Overlap of a priori ROIs (in blue) and results of the whole-brain analysis of strategy effect (pinyin + writing > pinyin; in red). The overlapped areas are shown in pale pink. Download Figure 5-2, TIF file.
Extended Data Figure 5-3
Accuracy of classifying participant’s language background. Download Figure 5-3, DOC file.
Extended Data Figure 5-1
Post hoc analysis of the effect of strategy on the learning effect. Mean and SE of each cluster in the two tasks for the pinyin (P) and pinyin + writing (P + W) groups was plotted. Download Figure 5-1, TIF file.
Effect of strategy on visual-auditory processing
The between-group contrast revealed that in VA_match, learning effect for the pinyin + writing group was greater than the pinyin group in the left precentral gyrus (PrC) that extended to the MFG, the left IPS/SPL, the right MFG-PrC, and right angular gyrus (cluster-wise corrected p = 0.05, cluster size determined by voxel-wise p of 0.05; df = 28; Table 3; Fig. 5B). Post hoc analysis revealed that for all but one cluster, the pinyin + writing group showed an increase in activation after learning, whereas the pinyin group did not show substantial learning effect in these areas. The only exception was the cluster centered at the right PrC, in which the pinyin + writing group had a marginally significant positive effect, and the pinyin group showed a significant decrease in activation (Extended Data Fig. 5-1).
Effects of strategy: uni-voxel ROI analysis
The effect of learning strategy was further investigated in coordinate-based predefined ROIs. The clusters revealed by the whole-brain analyses spatially overlapped with the predefined ROIs of left SPL and MFG (Extended Data Fig. 5-2). In the VA_match task, the left SPL presented greater activations for the pinyin + writing than the pinyin group (t = 2.06, two-tailed p = 0.048), and a marginal effect was found in the left MFG (t = 1.79, two-tailed p = 0.084; Fig. 5C). By contrast, no ROI showed group difference when the presented word form and pronunciation were mismatched.
In the Vl task, no ROI showed greater learning effect for the pinyin + writing group than the pinyin group. However, a tendency of greater learning effect for the pinyin group compared with the pinyin + writing group was found in the right FG (pinyin versus pinyin + writing, t = 1.92, two-tailed p = 0.066; Fig. 5C). Similar patterns were observed during the scrambled word processing (Vs) in the bilateral fusiform gyri: The pinyin + writing group showed a decreased activation after learning, and learning resulted in greater responses for the pinyin group than the pinyin + writing group (post > prelearning in the versus processing, right FG: Mp+w = −0.33, Mp = 0.10, SDp+w = 0.51, SDp = 0.28, pinyin versus pinyin + writing, t = 2.73, two-tailed p = 0.011; left FG: Mp+w = −0.14, Mp = 0.02, SDp+w = 0.22, SDp = 0.12, pinyin versus pinyin + writing, t = 2.27, two-tailed p = 0.031). The SPL or MFG did not reveal a group difference during the versus processing.
Effects of strategy: multivoxel pattern analysis within ROI
The group membership of each participant, namely, the pinyin or the pinyin + writing group, was identified based on the multivoxel patterns of other participants. The classifications resulted in mean accuracies of 0.73 and 0.70 when using the multivoxel patterns within left SPL during the VA_match and Vl processing respectively, both of which were significantly higher than the chance-level accuracy of 0.5 (Fig. 5D; random permutation-based significance tests, p < 0.05). In the left MFG, only the multivoxel patterns during VA_match processing showed a marginally significant mean accuracy of 0.63 (p = 0.05). Patterns of these two regions during the scrambled character processing or mismatched sound-print processing were not distinguishable between the groups of participants (all ps > 0.05): the accuracies were 0.50 (VA_mis in SPL), 0.60 (vs in SPL), 0.23 (VA_mis in MFG), and 0.3 (vs in MFG), respectively.
Classification accuracy within the right or left FG was not significantly different from chance level (Fig. 5D). However, during scrambled character processing, the patterns in the right FG resulted in a marginally significant accuracy of group identification at 0.63 (p = 0.05).
Results of language background classifications
Based on the multivoxel patterns in either ROI during either Vl or VA_match processing, the accuracies of classifying individual participants as being monolingual or bilingual were not significantly above chance level, ranging from 0.33 to 0.57 (Extended Data Fig. 5-3). Based on the multivoxel patterns in the clusters that presented the effect of strategy in the whole-brain analysis, the mean accuracy of classifying language background over participants was 0.53 (SD = 0.50) for the Vl processing and was 0.47 (SD = 0.50) for the VA_match processing, neither being significantly above chance level. These results suggested the effect of strategy group identified in the previous analyses was not an effect of bilingualism.
Discussion
This study investigated the neural functional alterations associated with learning to read Chinese as a second language. By using the pre-test and post-test paradigm, we observed spontaneous changes in two critical stages of reading, namely, the orthographic processing and the orthographic-to-phonological mapping. Typical areas for logographic reading in the brains of proficient alphabetic readers became more responsive in superficial processing of Chinese inputs after a week of learning. Although the behavioral learning effect was not strategy dependent, whether or not involving the writing practice modulated the neural responses of the left superior parietal cortices, left middle frontal gyrus, and the right fusiform gyrus, and these modulations were observed in different tasks associated with character processing.
Learning altered the neural responses in some of the areas that have been found to be commonly activated in reading in different languages, including the left IFG, left insula, the SMA and the adjacent anterior cingulate cortex, the left fusiform gyrus, and the bilateral extrastriate cortices (Tan et al., 2005a; Maisog et al., 2008; Richlan et al., 2009; Rueckl et al., 2015). Among these regions, the IFG-insula has been identified as one of the language-general speech-print convergence regions, in that this area showed correlated responses to visual and auditorily presented words in multiple languages, including Chinese (Rueckl et al., 2015). Moreover, the logographic-specific areas in the left MFG and left SPL also showed increased activation after learning. Activation of the typical alphabetic reading-related areas in the left posterior temporal gyrus and left angular gyrus were not found altered by reading Chinese. The above-mentioned results have been observed during both the visual and the visual-auditory processing. These findings were consistent with the view that reading Chinese as a second language showed an accommodation pattern (Nelson et al., 2009; Ma et al., 2020). The present results further revealed that the accommodation appeared in broader reading networks in addition to the visual perceptual processing areas.
Although the postlearning improvement of behavioral performance was not affected by learning strategy, the interaction of strategy and learning stage in neural response suggested some logographic-specific areas were modulated by whether the learning involved writing practice. Some previous behavioral studies have shown the effect of handwriting practice on literacy acquisition (Wiley and Rapp, 2021) or the correlation between the two abilities (Tan et al., 2005b), while others have found a dissociation between writing and reading in Chinese. For instance, knowledge of how a character was written did not influence character processing in proficient readers (Zhai and Fischer-Baum, 2019). Patient with left temporoparietal lesions presented complete writing deficits and poor orthographic awareness, but was able to perform perfectly in Chinese reading task (Bi et al., 2009). The present results showed the effect of writing on reading was limited from a neural perspective: For the passive viewing process, uni-voxel analyses revealed that learning by pinyin decreased the activations in the right supramarginal gyrus, which contributed to the group difference in the learning effect (Fig. 5A; Extended Data Fig. 5-1), whereas the multivoxel analysis showed that writing also altered the activation patterns of the left SPL. Although the right SMG is not considered part of the canonical reading network, it has been identified as a cross-language speech-print convergence area, where the response magnitudes to written and spoken words are correlated over participants (Rueckl et al., 2015). An inference according to this view is that learning characters by associating the visual form with pinyin has resulted in less reliance on this universal sound-print association area, and potentially more reliance on the Chinese-specific neural substrates, such as the left MFG and SPL as identified in the visual-auditory processing.
During visual-auditory processing, the effects of learning strategy were driven by the greater postlearning increase of the pinyin + writing group compared with the pinyin group. The identified brain areas can thus be viewed as areas to which the additional writing practice has brought additional response increase. These results have shown spatial overlap with the meta-analysis-determined logographic-specific ROIs (Extended Data Fig. 5-1) in the left SPL and MFG. The joint results of uni-voxel and multivoxel analyses on strategy effect have suggested that (1) both strategies have increased the sensitivity of left SPL to characters; (2) the responses of L SPL to characters are modulated by learning strategy; (3) involving writing practice during learning tends to slightly increase the MFG response only during the orthographic-phonological mapping; and (4) the right FG is modulated by strategy during the uni-modal visual processing, but this effect is not orthographic-specific, which differ from the response of the left FG. We discuss these implications by ROIs below.
Left superior parietal lobule
First, learning led to greater responses of the left SPL in both the orthographic (Vl) and the orthographic-phonological (VA_match) processing for both of the strategy groups. Second, adding writing-based learning caused greater responses during the orthographic-phonological processing as compared with the pinyin-only strategy. Third, while the uni-voxel analyses revealed no group difference in the response magnitude during the orthographic processing, the between-group differences in multivoxel patterns associated with the orthographic processing were reliable enough to predict the learning strategy used by individual learners according to the multivoxel patterns of other learners. Forth, this area was unable to identify participant’s learning strategy when the displayed character was paired with a wrong pronunciation, suggesting the strategy effect was sensitive to the congruency of multimodal input, or in other words, the strategy effect was manifested based on participants’ knowledge about character identity. Therefore, the activities in left SPL during both orthographic processing and orthographic-phonological mapping tasks relied on how the characters were learned. The findings concur with the proposal for IPS/SPL being part of the reading network that is specialized for fine-grained visuospatial analysis and motor gesture inference (Kuo et al., 2004; Siok et al., 2009; Nakamura et al., 2012). The IPS/SPL has been found to be sensitive to visual distortion of word (Nakamura et al., 2012), activate less in a size judgment task (Siok et al., 2009), and show decreased resting-state functional connectivity with the left MFG (Zhou et al., 2015) in Chinese dyslexic children compared with the nondyslexics, suggesting the role of SPL in normal reading might not be just correlational. Our results further suggest that writing-based learning strengthens the involvement of this dorsal visuomotor pathway in reading, even when there was no explicit cue or demand of the visuomotor encoding.
Left middle frontal gyrus
The uni-voxel results of learning and strategy in the left MFG were similar to the SPL: MFG showed increased responses after learning in both tasks, except that the effect for the pinyin group during VA_match task was only marginal. The pinyin + writing strategy resulted in a slightly stronger learning effect than the pinyin group only during the processing of orthographic-phonological mapping task. Unlike the SPL, the MFG was not found to display cross-participant consistent, strategy-specific multivoxel patterns during orthographic-only (Vl) processing. Thus, our interpretation of the results with caution was that writing-based learning seemed to increase the sensitivity of left MFG to print-to-sound matching, rather than orthographic processing per se Multiple roles have been proposed for MFG in logographic reading, such as visuospatial analysis (L. Liu et al., 2009; C.Y. Wu et al., 2012), orthography-semantics association (Siok et al., 2004; J. Wu et al., 2015), representing addressed phonology of Chinese words (Tan et al., 2005a; Booth et al., 2006; Kwok et al., 2019; A. Li et al., 2022), or encoding writing gestures (Nakamura et al., 2012). The present results showed that a sign of learning effect of the left MFG was observed during orthographic-to-phonological mapping process but not passive reading, which was consistent with insights from a previous meta-analysis that the activity in MFG is task dependent (Zhao et al., 2017). Because the joint presentation of written form and sound requires the processing of character identity, we speculate that the tendency of increased activity in left MFG reflects the development of knowledge about the sound-print association of a character. Because the learning effect in MFG was slightly amplified when participants learned to write, we speculate that the writing has provided additional assistance, which might be the increased orthographic awareness, for the learners to establish the sound-print association.
Right fusiform gyrus
The responses in the right FG were distinct from the left SPL and MFG. Unlike the left FG, SPL, or MFG, R FG did not show an overall increase in the responses after learning. Moreover, for both the scrambled character and the learned real character, the R FG showed increased activations in the second scan, and this effect was reliably shown only in the pinyin group, only when the task was the unimodal visual processing (Fig. 5C). These findings suggest that the R FG is not specialized for recognizing scripts but for recognizing domain-general complex visual layout. A week of exposure to characters might increase its sensitivity to complex layout in general, but the multimodal pinyin-and-writing-based learning had downplayed the engagement of this area in character processing. Similarly, when the paradigm explicitly required multimodal knowledge of the characters, the R FG played less of a role as the other areas (MFG and SPL) took over the task of orthographic-phonological mapping.
One limitation of the study was that involving writing led to additional practice for that group of participants. The dilemma is that the control of workload means relatively less pinyin-based practice in the pinyin + writing group, the same amount of pinyin-based practice means more overall practice for the pinyin + writing group, whereas a writing-only learning procedure is unnatural and unlikely to be adopted in a realist situation for typically developed learners. We choose to examine the effect of additional writing practice, which results in unbalanced workload between groups. Future study is required to address whether the effect identified in the present work is writing-specific or just an effect of multimodal training, or even just an effect of more practice. It also remains an open question whether the activities of the so-called “logographic-specific” areas are also modulated by learning strategy if the target language is an alphabetic language. There has been evidence that a left IFG-premotor area (centered at [−42, 6, 20], close to the center of the MFG cluster used in the present study at [−46, 18, 28]) is sensitive to the correct moving trajectories of writing, and the effect seemed to be consistent in French words and Chinese characters (Nakamura et al., 2012). Such finding suggests that it might be the common processing of handwriting that results in the specialization of these seemingly language-specific areas. Future studies are required to directly investigate the effect of learning strategy on alphabetic languages.
Another limitation of the study was that the uni-voxel effects were only present at cluster level, indicating low spatial specificity. On the other hand, multivoxel patterns showed a strategy-related, cross-subject consistent patterns in the a priori regions. These findings might suggest a more distributed, i.e., less spatially specific, patterns for representing characters for the L2 learners.
Two groups of participants showed different learning effects in brain activations in the absence of behavioral differences. Previous study has observed neural differences in processing word and nonwords on adult L2 learners after several hours of learnings, when the behavioral performance was still at chance level (McLaughlin et al., 2004). Given the role of writing in Chinese learning, it is possible that neural group difference is a harbinger of overt behavioral differences.
Overall, the present study has revealed the emergence of logographic reading network after a week of learning in adult alphabetic reader’s brain. The learning effect in logographic-specific areas was not entirely dependent on, but modulated by the learning strategy. The present finding on group differences has suggested the additional effects of writing-based learning.
Footnotes
The authors declare no competing financial interests.
This research was supported by National Natural Science Foundation of China Grants 31970987 (to Q.C.) and 32100857 (to J.W.), the Basic Research Project of Shanghai Science and Technology Commission Grant No. 19JC1410101, and the “Flower of Happiness” Fund Pilot Project of East China Normal University Grant No. 2019JK2203.
This is an open-access article distributed under the terms of the Creative Commons Attribution 4.0 International license, which permits unrestricted use, distribution and reproduction in any medium provided that the original work is properly attributed.
References
In this issue

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}



