

Abstract
This study assesses the involvement in human motor learning, of the ventrolateral prefrontal cortex (BA 9/46v), a somatic region in the middle frontal gyrus. The potential involvement of this cortical area in motor learning is suggested by studies in nonhuman primates which have found anatomic connections between this area and sensorimotor regions in frontal and parietal cortex, and also with basal ganglia output zones. It is likewise suggested by electrophysiological studies which have shown that activity in this region is implicated in somatic sensory memory and is also influenced by reward. We directly tested the hypothesis that area 9/46v is involved in reinforcement-based motor learning in humans. Participants performed reaching movements to a hidden target and received positive feedback when successful. Before the learning task, we applied continuous theta burst stimulation (cTBS) to disrupt activity in 9/46v in the left or right hemisphere. A control group received sham cTBS. The data showed that cTBS to left 9/46v almost entirely eliminated motor learning, whereas learning was not different from sham stimulation when cTBS was applied to the same zone in the right hemisphere. Additional analyses showed that the basic reward-history-dependent pattern of movements was preserved but more variable following left hemisphere stimulation, which suggests an overall deficit in somatic memory for target location or target directed movement rather than reward processing per se. The results indicate that area 9/46v is part of the human motor learning circuit.
Significance Statement
Prefrontal cortex may contribute to motor learning as it is known to be involved in planning, executive control, and motivation or reward processing (Miller and Cohen, 2001). Here, we focused on ventrolateral prefrontal cortex (BA 9/46v), an area which has been shown to be linked neuroanatomically and electro-physiologically to sensorimotor regions of the brain and to circuits involved in reinforcement. Using continuous theta burst stimulation (cTBS) to this region before a reinforcement-based motor learning task, we found a significant reduction in learning. This suggests that this zone in the lateral prefrontal cortex contributes to motor learning which is mediated by reward.
Introduction
The brain structures involved in human motor learning have been studied extensively. Areas in frontal and parietal cortex, cerebellum and basal ganglia have each been shown to contribute to learning and retention, although their weighting differs between tasks. In contrast, prefrontal cortex has received little attention to date in the context of motor learning (but see Anguera et al., 2010 and Codol et al., 2020) although regions within prefrontal cortex are known to be neuroanatomically connected to sensorimotor related regions of other structures which are implicated in learning. The present study focuses on ventrolateral prefrontal cortex (BA 9/46v), a somatic region in the middle frontal gyrus directly above the ascending anterior ramus of the lateral fissure that separates BA 44 and 45, and has neuroanatomical connections to premotor, somatosensory and basal ganglia structures. By disrupting this area using magnetic brain stimulation, we test for its participation in human motor learning.
The focus on 9/46v is motivated by both electrophysiological findings and neuroanatomical connectivity. Studies in nonhuman primates have identified a homologous somatic region in the inferior bank of the principal sulcus which is interconnected with areas PF and PFG in the inferior parietal lobe (or supramarginal gyrus in humans) and second somatosensory cortex in the parietal operculum (Preuss and Goldman‐Rakic, 1989; Petrides and Pandya, 2002; for a summary, see Yeterian et al., 2012). This area also communicates with the hand area of ventral premotor cortex and likewise receives inputs from globus pallidus and substantia nigra of the basal ganglia (Middleton and Strick, 2002). In electrophysiological studies, this same region has been implicated in somatic sensory memory and decision-making (Romo et al., 1999).
We tested for the involvement of 9/46v using a reinforcement learning task. In reinforcement-based motor learning, positive feedback provides behavioral reinforcement, inducing plasticity in motor, somatic and reward-related networks (Bernardi et al., 2015; Sidarta et al., 2016). The involvement of the middle frontal gyrus in reinforced sequence learning has been demonstrated using repetitive TMS (Dayan et al., 2018). Area 9/46v involvement in both reinforcement learning (Fermin et al., 2016) and visuomotor adaptation (Anguera et al., 2010) has been observed in studies using fMRI. Other parts of the prefrontal cortex, in particular, ventromedial prefrontal cortex and orbitofrontal cortex have been implicated in reward-based learning more generally. In the nonhuman primate literature, activity in dorsolateral prefrontal cortex during a delay period was found to be related to the amount of reward received and the type of responses to be performed (Hikosaka and Watanabe, 2000; Wallis and Miller, 2003). Moreover, there is evidence that the lateral prefrontal cortex carries reciprocal projections with the midbrain dopaminergic neurons (Williams and Goldman-Rakic, 1998; Frankle et al., 2006), as well as with the orbitofrontal cortex (Barbas and Pandya, 1989).
Participants in the present study were assigned to one of three experimental conditions in which continuous theta-burst transcranial magnetic stimulation (cTBS) was applied to either left or right 9/46v, with the goal of disrupting activity in the target zone, or to a sham stimulation group. This was followed by a motor learning task in which participants performed reaching movements to a hidden target. The participants were given positive feedback when the movement was successful, that is, when it had landed in the target zone. We found that disruption of area 9/46v before learning had a detrimental effect on both learning rate and on the overall number of successful (and thus rewarded) movements. This is consistent with its participation in reinforcement-based motor learning.
Materials and Methods
Participants
Fifty-four healthy right-handed young adults (19 men, 35 women) were recruited and randomly assigned into either a left hemisphere (left 9/46v, N = 18), right hemisphere (right 9/46v, N = 18), or sham stimulation condition (sham, N = 18). Handedness was assessed using the Edinburgh handedness inventory (Oldfield, 1971). All procedures were approved by the McGill University Faculty of Medicine Institutional Review Board and participants provided written informed consent.
Experimental design
Participants held a vertical handle attached to the end of a 2-df robotic manipulandum (Interactive Motion Technologies). They were seated with their right shoulder abducted to ∼70° and the elbow supported by an air sled. A semi-silvered mirror, which served as a display screen, was placed just below eye level and blocked the vision of the arm and the robot handle (Fig. 1A). A white start circle, 20 mm in diameter, was positioned on the display screen ∼30 cm in front of the participant, on the body midline. A 1-cm white arc was shown on the left of the screen during familiarization trials (Fig. 1B). During the familiarization phase, participants were instructed to move to any point on the arc after the “Go” cue appeared and to make straight movements without corrections. A cursor, which represented the instantaneous handle position in space, was removed once the arm moved outside of the white start circle. The required movement duration was 500–700 ms, but there was no penalty if the movement did not end on time or outside the target arc. Once the movement ended, the robot brought the arm back to the start position.

Participants learned to make movements to a hidden target, and positive feedback was provided for successful movements. A, Participants made movements holding a robotic manipulandum. B, Schematic of the task. Participants made outward movements. If the movement direction fell within the hidden target zone, positive feedback was provided to indicate success. No feedback was given in the case of an unsuccessful movement. C, Experimental sequence. MEPs were elicited from the motor hot-spot in the left or right hemisphere before stimulation (cTBS to right or left 9/46v or sham stimulation). MEPs were again recorded 10 min after stimulation followed by the motor learning trials. In the no-feedback session at the end, participants were not provided with feedback on the success of the movement. D, Location of the stimulation site in representative participants from the left 9/46v and right 9/46v condition, shown in the sagittal (right panel) and coronal (middle panel) planes. The average location of the stimulation site (red circle) across participants in the MNI brain.
Following the familiarization training, the target arc was removed. The participant was instructed to move toward the now hidden arc and was told there was a target located in the arc. Then, each participant made 15 movements without receiving feedback of any kind. A target direction was then set for each subject separately to correspond to the direction of the first movement after the 15th trial that fell between 110° and 160° (second quadrant at the left). Positive feedback (an animated explosion, a pleasant tone, and a score) was provided for this movement. Participants were told that their task was to repeat the same successful movement throughout the course of training. Positive feedback was dependent solely on movement direction at peak velocity although participants were provided feedback on distance for training purposes during familiarization trials. The width of the target zone was 5°, and positive feedback was provided if the angular deviation was within ±2.5° of the center line. The width and position of the reinforced direction were fixed. Altogether, the participants completed four blocks of 50 training trials with positive feedback when successful. This was followed by 25 further movement trials with no feedback. For these trials, participants were told to aim in the direction in which they had been rewarded previously. They were also told that no reward would be given even if they were accurate. The sequence of different phases of the experiment is shown in Figure 1C.
Stimulation sites
Before the study, each participant underwent an MRI scan at the Montreal Neurologic Institute Brain Imaging Centre. Structural images were acquired with a T1-weighted 3D MPRAGE sequence as follows: TR = 2300 ms; TE = 2.98 ms; slices = 192; thickness = 1 mm (no gap); FA = 90°; and FOV = 256 × 256 mm, iPAT mode = ON (acceleration factor 2×).
The stimulation location in area 9/46v was identified for each subject separately, in the following manner. The identification starts with pars opercularis and pars triangularis in the inferior frontal gyrus, which are separated by the ascending anterior ramus of the lateral fissure (Petrides and Pandya, 2002). This ascending sulcus runs up from the lateral fissure and is almost perpendicular to the inferior frontal sulcus. The stimulation site, as shown in Figure 1D, lies in the middle frontal gyrus, medial to ascending anterior ramus of the lateral fissure and between two posterior middle frontal gyrus sulci, the posterior middle frontal sulcus (anterior) and posterior middle frontal sulcus (intermediate; Petrides, 2012). The mean stimulation location is shown in each hemisphere in standard MNI coordinates: (−46, 26, 30 mm) for the left 9/46v and (52, 26, 32 mm) for the right 9/46v. The stimulation site was marked and maintained using Brainsight (Rogue Research). The TMS coil position was tracked using a three-dimensional optical system (Polaris System, Northern Digital).
Stimulation protocol
The theta-burst magnetic stimulation magnitude was based on the resting motor threshold (RMT) in primary motor cortex. The position at which left or right motor cortex was maximally excitable in eliciting motor-evoked potentials (MEPs) in the contralateral FDI muscle was determined, using single-pulse TMS (Magstim200 stimulator). The coil was placed tangentially on the scalp with the handle pointing backward and laterally at a 45° angle away from the midline. The EMG response of the FDI muscle was recorded using Ag-AgCl surface electrodes. The RMT was defined as the minimum intensity required to elicit at least 5 MEPs (>50 mV peak-to-peak amplitude) in 10 consecutive single-pulse stimulations.
cTBS (Goldsworthy et al., 2012) was used to disrupt neural activity in left or right 9/46v before learning. cTBS was applied in two trains (10 min apart) of repetitive biphasic magnetic pulses (Magstim Super Rapid Stimulator) at 70% intensity of the RMT for the FDI muscle (based on left and right M1 separately, recorded using a Magstim 200 monophasic stimulator). Each train of cTBS comprised 600 pulses applied in bursts of three pulses at 50 Hz, with bursts repeated at a frequency of 5 Hz, corresponding to a total train length of 40 s. cTBS stimulation was delivered with the coil handle pointed downward.
To test for possible indirect effects of cTBS on motor cortex, we applied single-pulse TMS to the motor hotspot, at an intensity sufficient to evoke 20 MEPs of ∼500–1000 μV (peak-to-peak amplitude) both before stimulation and at the same intensity, 10 min after cTBS.
Statistical analysis
Directional error was quantified as the angular deviation (AD) from the true target direction (center of the target zone) at the maximum velocity. The absolute angular deviation, |AD|, was used as a measure of movement accuracy. The number of trials with positive feedback and absolute angular deviation were used to quantify learning. The rate of learning was computed through a linear fit to the absolute angular deviation as a function of learning trials. The slope of the fitted line was used as a measure of the learning rate. One-way ANOVA was performed on learning rates across experimental conditions. One-way ANOVA was also performed on mean change in absolute angular deviation from the first block to the last block of training.
We also computed a linear fit to the mean percentage of rewarded movements across participants over the course of training. One-way ANOVA was performed on changes in the percent of rewarded movements between the first and last block of training. A two-way ANOVA was performed to assess the effect of reward history on movement variability from the nth to n + 1th trial in different experimental conditions. Post hoc tests were corrected for multiple comparisons.
To evaluate possible effects of cTBS on motor cortex, MEPs recorded post-cTBS were expressed as a percentage of pre-cTBS MEPs, using mean MEP amplitude on a per subject basis. One-way ANOVA was used to test for the difference between experimental conditions.
Results
Participants held the handle of a robotic manipulandum (Fig. 1A) and made reaching movements toward a hidden target (Fig. 1B, shaded gray area) in four blocks of 50 trials each. Participants were rewarded for successful movement in the target direction. To assess the contribution of the ventrolateral prefrontal cortex to motor learning, cTBS stimulation was applied before learning in different groups of subjects in each hemisphere separately. Figure 2A shows data from a representative subject in each experimental condition. Movement paths shown in blue are for successful (rewarded) movements and those in red are for unsuccessful movements. Note that the overall direction differs in the three conditions because of individual differences in target location. The figure shows that movement paths were similar in the three experimental conditions at the beginning of training (block 1). At the end of training (block 4), participants in the sham condition moved more consistently to the target than participants who received stimulation to either left or right 9/46v (Fig. 2A).
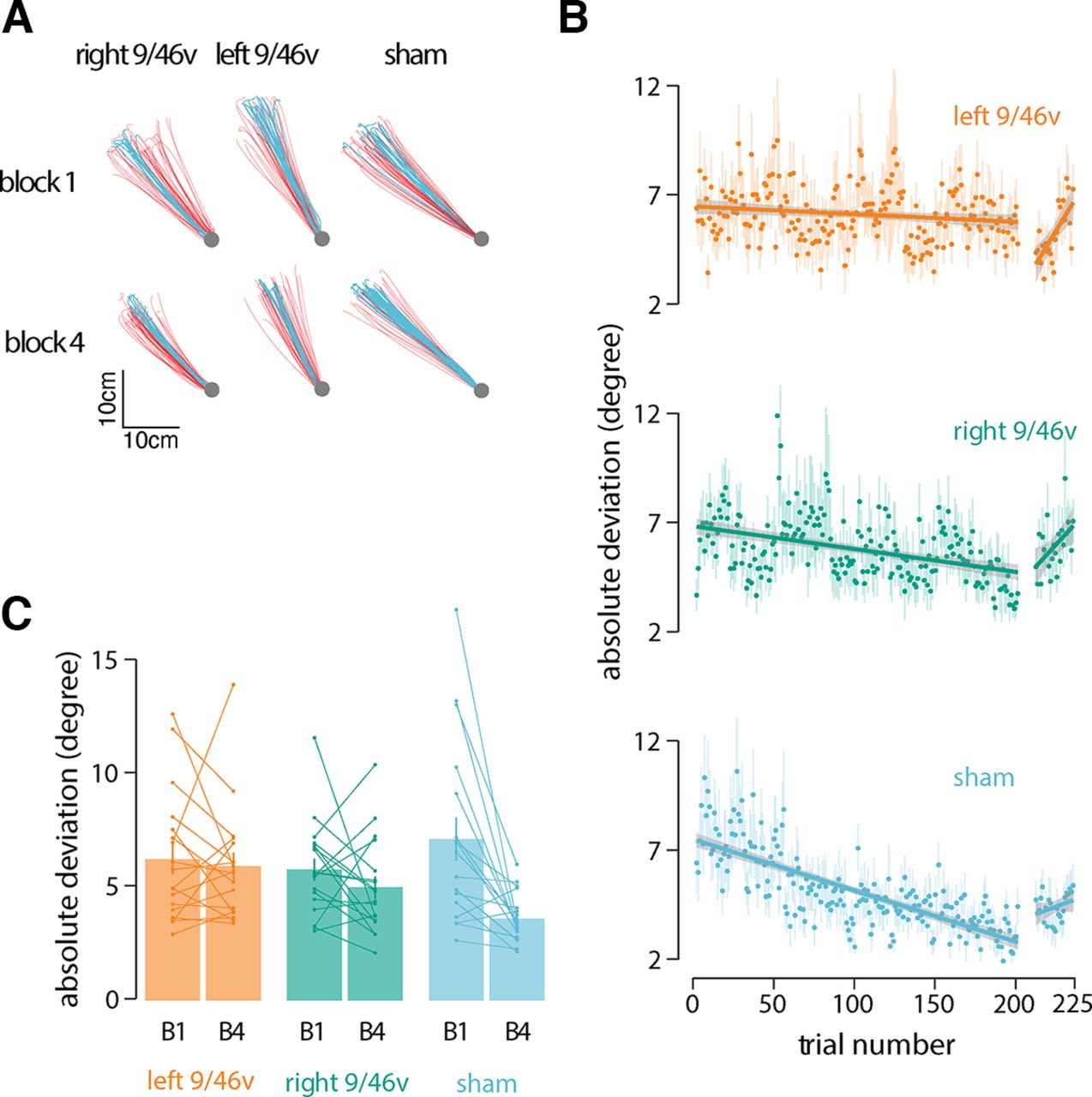
Suppression of left 9/46v using cTBS disrupts motor learning. A, Hand paths of a representative participant from each group at the start (block 1) and end of training (block 4). Hand paths shown in red are for unsuccessful movements, and those in blue are for successful movements. B, Mean absolute deviation from the center of the target zone over the course of training. The linear fit is shown across learning trials and no-feedback trials separately. The shaded region represents ±SEM. The rate of learning was less in participants who received stimulation over left 9/46v than those who received sham stimulation. C, Mean absolute deviation in the first and last block of the training. Participants in the sham stimulation condition showed a greater reduction in |AD| than participants in the left 9/46v condition.
Reduction in the angular deviation from the target direction, |AD|, over the course of the motor task provides a measure of improvement in accuracy as a result of learning (Fig. 2B). The rate of reduction in |AD| was estimated for each subject separately. ANOVA applied to the slope estimates indicated the rate of angular deviation reduction differed significantly among stimulation conditions (F(2,51) = 4.19, p = 0.02; Fig. 2B). The rate of learning was slower in participants who received stimulation over left 9/46v (slope = −0.002, 95% CI = −0.01, 0.006) than those who received sham (slope = −0.022, 95% CI = −0.033, −0.011) stimulation (p = 0.016). There was no significant difference in the learning rate between the sham and right 9/46v (slope = −0.009, 95% CI = −0.019, 0.001) conditions (p = 0.16). Another indicator of learning is the change in the |AD| from the beginning of the learning session to the |AD| at the end (Fig. 2C). The mean change in |AD| from the first to last learning block showed significant differences between conditions (F(2,51) = 4.93, p = 0.01). Post hoc tests indicated that participants in the sham stimulation condition showed a greater reduction in |AD| than participants in the left 9/46v condition (p = 0.014).
Participants also performed no-feedback trials after the initial learning session in which feedback on movement success was withheld. We found no significant difference in |AD| between conditions (F(2,51) = 0.57, p = 0.56) nor was there a significant difference in the slope between the groups in no-feedback trials (F(2,51) = 1.82, p = 0.17). The slope in these trials for the sham condition was not reliably different from zero (p = 0.56). The slopes in the left 9/46v (p = 0.003) and right 9/46v (p = 0.05) conditions were both found to be reliably greater than zero indicating a progressive reduction in accuracy for the learned target direction.
During the motor learning task, participants were instructed to maximize the number of rewarded trials. Figure 3A shows the percentage of rewarded trials over the course of learning. Participants in the sham stimulation group showed a steady increase in the number of successful movements (slope = 0.119, 95% CI = 0.091–0.146) compared with participants in the left 9/46v stimulation condition (slope = 0.011, 95% CI = −0.014–0.038). Participants in the right 9/46v condition showed values intermediate between those in the other two conditions (slope = 0.073 95% CI = 0.047–0.098). Statistical tests were conducted to assess changes in the percent of rewarded (Figure 3B) movements between the first and the last block of training. The change scores (increase from start to end of training in the percent of rewarded trials) differed significantly across conditions (F(2,51) = 6.18, p = 0.003). Post hoc tests indicated a reliable difference in reward change scores between the left 9/46v and sham stimulation conditions (p = 0.002). Specifically, participants in the sham stimulation condition received more rewards as learning progressed, whereas participants who received stimulation to left 9/46v showed no improvement at all. There was no difference in reward change scores for participants in the right 9/46v and sham stimulation conditions (p = 0.10). One sample t tests indicated that the reward change from the first to last block for participants in the left 9/46v condition was not reliably different from zero (t(17) = −0.13, p = 0.89).

Suppression of left 9/46v using cTBS leaves reinforcement learning intact. A, Mean percentage of rewarded trials over the course of training. A linear fit is shown across learning trials. The shaded region represents ±SEM. B, Mean percent of rewarded movements in the first and last block of the training. Participants in the sham stimulation condition received more rewards as learning progressed, whereas participants who received stimulation to left 9/46v showed no improvement at all. C, Mean absolute change in movement direction between the current trial (nth trial) and the subsequent trial (n + 1th trial) as a function of the history of rewarded movements. Reward history included three most recent movements (n, n–1, and n–2 trial), where at least one of these movements was rewarded. The left 9/46v group showed the same basic reward-history-dependent pattern as the other conditions but with greater change in direction overall. This suggests that the learning deficit after left 9/46v suppression is not because of inability to process reward but likely because of a deficit in memory for target direction.
One possible reason for not showing improvement over the course of training in the left 9/46v condition was that stimulation impaired the capacity to benefit from reward. To assess this possibility, we computed the absolute change in movement direction between the current trial (nth trial) and the subsequent trial (n + 1th trial) as a function of the history of rewarded movements. The analysis, shown in Figure 3C, was conducted over the three most recent movements (n, n–1, and n–2 trial), under conditions where at least one of these movements was rewarded. It can be seen that there is a graded pattern of absolute change in movement direction, which is least following three rewarded movements and greatest when only a single movement is rewarded. Thus, a normal although more variable reward-history-dependent pattern is obtained following cTBS to left 9/46v. A two-way ANOVA with reward history and the stimulation condition as the independent factors and Δm, the absolute change in movement direction, as dependent variable revealed a significant effect of reward history (F(6,306) = 53.85, p < 0.001) indicating that change in movement direction is dependent on the number of rewarded trials in the recent past. The overall magnitude of the change in direction, Δm, marginally differed across stimulation conditions (F(2,51) = 2.86, p = 0.06). Bonferroni-holm corrected post hoc tests indicated that participants in the left 9/46v stimulation condition showed greater change in direction than participants in the sham (p = 0.007) and right 9/46v conditions (p = 0.02; Fig. 3C). There was no indication that the reward-history dependent pattern differed between conditions, that is, there was no significant interaction between stimulation conditions and reward history (F(12,306) = 0.43, p = 0.94). In summary, participants in the left 9/46v group showed the same basic reward-history dependent pattern as the other conditions but with greater change in direction overall. This suggests that the learning deficit in the left 9/46v condition is not because of an inability to benefit from reward per se.
We have also assessed the possibility that 9/46v stimulation affected the movements themselves. We compared three basic movement parameters, peak velocity, movement amplitude and movement duration across stimulation conditions (Fig. 4C). There were no significant differences between conditions (left and right 9/46v and sham condition) in peak velocity (F(2,51) = 1.28, p = 0.28), movement amplitude (F(2,51) = 0.10, p = 0.90) and movement duration (F(2,51) = 2.12, p = 0.13). We also tested the possibility that 9/46v stimulation indirectly affected primary motor cortex and that deficits in learning occurred as a consequence. We assessed MEPs before and after stimulation (representative sample in Fig. 4A) and found that there were no significant differences in peak-to-peak MEP amplitude across experimental conditions (F(2,51) = 0.56, p = 0.57; Fig. 4B). Overall, this suggests that cTBS to 9/46v did not alter basic movement patterns nor did it indirectly act on primary motor cortex.

cTBS over left or right 9/46v did not alter the excitability of motor cortex or basic movement parameters. A, Mean time series of MEPs recorded from the FDI muscle pre-cTBS (blue) and post-cTBS (red) from a representative participant in each experimental condition. The TMS pulse occurs at time = 0 ms. The shaded regions are ±SEM across 20 MEPs. B, Mean change in amplitude of MEPs measured 10 min after cTBS (computed as a percentage of pre-cTBS MEPs). Error bars give the SE across participants. C, Mean movement duration, peak velocity, and movement amplitude across experimental conditions. cTBS to either left or right 9/46v did not modify the movement parameters.
Discussion
The present study used transcranial magnetic stimulation to disrupt activity in ventrolateral prefrontal cortex (9/46v) to test its involvement in human motor learning. Participants held the handle of a robot arm and made movements to a hidden target. Positive reinforcement was provided when the movement ended in the target zone. cTBS stimulation was delivered before learning either to left or right 9/46v; control participants received sham TBS. It was found that cTBS to left 9/46v all but eliminated improvements in movement as measured by changes in angular direction relative to the target. cTBS also led to a significant reduction in the number of reinforced trials in comparison to sham stimulation. The disruption of 9/46v did not adversely affect the ability to use reward as indicated by a normal, although more variable, dependence of movement direction on reward-history. As there is no visual feedback whatsoever in this task, this latter observation suggests that while a sensitivity to reward is preserved following disruption of 9/46v, there is an across-the-board deficit in somatic memory for target location or target directed movement, a result consistent with previous demonstrations of 9/46v involvement in somatic memory (Romo et al., 1999). Overall, the present results indicate that area 9/46 is part of a network that participates in human motor learning.
No feedback trials at the end of training are consistent with this conclusion. For both left and right 9/46v there is a progressive increase during no-feedback trials in angular deviation relative to the target which is suggestive of a progressive loss of information during retention testing. In contrast, the slope is not different from zero following sham stimulation indicating that retention is unimpaired when 9/46v is intact. It should be noted that while for left 9/46v stimulation retention performance appears to be initially better than that observed during learning, the values at the start of the retention test are wholly with the range of those obtained over the course of training.
Although it is in prefrontal cortex, several studies have shown that, in nonhuman primates, area 9/46v has both inputs and outputs to somatic regions of the brain, including connections to second somatosensory cortex, cortical areas PF and PFG (supramarginal gyrus) in the inferior parietal lobe (Preuss and Goldman‐Rakic, 1989; Petrides and Pandya, 2002; Gerbella et al., 2013) and also to ventral premotor cortex (Dum and Strick, 2005). In humans, an analogous pattern of connectivity between this same set of areas has been reported using resting-state fMRI and diffusion tractography (Barbeau et al., 2020) and between the ventral portion of the middle frontal gyrus and ventral premotor cortex (Catani et al., 2012). Outputs from the basal ganglia to area 9/46v have also been reported (Middleton and Strick, 2002). As such, this area is well placed to funnel both somatic (error-based) and reinforcement-based information to frontal motor areas in support of learning. The involvement of area 9/46v in somatic memory and decision-making has been documented in studies in which nonhuman primates are required to hold in memory vibrotactile information (delivered to the fingertips) and to make judgements regarding relative frequency. Neurons in this region have been found to show both memory dependent and decision-making related activity (Romo et al., 1999).
Studies of spatial working memory in humans also report activity in this same region of prefrontal cortex (D’Esposito et al., 1998; Owen et al., 2005). In strictly behavioral studies, there is evidence of a relationship between somatosensory memory and reinforcement learning (Sidarta et al., 2018) and also between visuospatial memory and sequence learning (Bo and Seidler, 2009; Bo et al., 2009). In the study by Sidarta and colleagues (Sidarta et al., 2018), using a task similar to the one in the present study, it was found that individuals with better sensory memory for their own movements also showed greater learning.
Reinforcement learning has been characterized as involving both repetition of successful movements (exploitation) or the selection of new movements following unsuccessful trials (exploration). The present results suggest that disruption of 9/46v leaves both processes intact as indicated by the finding that a normal, but more variable, dependence of movement on reward history is preserved. The deficits in learning appear instead to be memory dependent. This finding shows that it is possible experimentally to partially dissociate the contribution of brain structures involved in reward and sensory memory in motor learning. Area 9/46v involvement in human motor learning has been reported in studies involving both reinforcement and error-based learning where learning-related activity is observed in both task-based and resting-state scans (Anguera et al., 2010; Sidarta et al., 2016).
It was found that disruption of activity in right 9/46v resulted in a reduction in both the rate of learning and the number of reinforced trials. Although these effects were not statistically different from measures of the same variables when stimulation was delivered to left 9/46v, nor when sham stimulation was delivered, the results for right hemisphere stimulation are intermediate between the two. Activity in right 9/46v has been observed previously in humans in both reinforcement learning and error-based learning tasks (Anguera et al., 2010; Sidarta et al., 2016). It has also been observed previously in sensory memory tasks in nonhuman primates (Romo et al., 1999). The extent to which there is hemispheric specialization in the contribution of area 9/46v to learning is uncertain. In humans, there is substantial interhemispheric connectivity in prefrontal cortex (Zarei et al., 2006). Moreover, interhemispheric propagation of TMS stimulation in prefrontal cortex has been reported (Voineskos et al., 2010), which makes possible the idea that the partial disruption of learning which occurs when right 9/46v is stimulated occurs as a result of indirect effects on the left hemisphere.
The involvement of a somatic network in human motor learning is supported by the finding that areas which show somatic memory and decision-making activity in nonhuman primates, second somatosensory cortex, ventral premotor cortex, supplementary motor area, and ventrolateral prefrontal cortex (Romo et al., 2012) are likewise areas that show learning-related changes in functional connectivity following motor learning in humans (Vahdat et al., 2011). This somatic network which also includes inferior parietal cortex (supramarginal and angular gyrus; Barbeau et al., 2020) fits within a broader interconnected sensorimotor network which includes primary motor and somatosensory cortex, medial wall motor areas, the superior parietal lobule, basal ganglia, and cerebellum (for review, see Rizzolatti and Luppino, 2001; Bostan and Strick, 2018). While each of these areas might contribute to the learning observed in the present study, the elimination of learning following cTBS to 9/46v suggests a causal contribution of this specific area in the context of reinforcement motor learning in humans.
In summary, it was found that cTBS stimulation to area 9/46v in prefrontal cortex disrupts motor learning without affecting the movements themselves. The deficit appears to be primarily related to impaired somatic memory for target location or target directed movement; disruption of 9/46v leaves reinforcement-based learning largely intact. 9/46v is distinguished from other regions of prefrontal cortex by its significant pattern of somatosensory connectivity. Area 9/46v thus appears to be part of the human motor learning circuit.
Footnotes
The authors declare no competing financial interests.
This work was supported by the Gouvernement du Canada, Canadian Institutes of Health Research (IRSC) Grant PJT 165987 (to D.J.O.).
This is an open-access article distributed under the terms of the Creative Commons Attribution 4.0 International license, which permits unrestricted use, distribution and reproduction in any medium provided that the original work is properly attributed.
References
In this issue

{kind=link}
{kind=link}
{kind=link}
{kind=link}



