

Article Figures & Data
Figures
- Figure 1.
scRNA-seq data analysis reveals gene expression of EAAT2 in all mouse retinal BCs and co-expression with EAAT5 in several BC types. A, Gene expression patterns of EAATs in different types of BCs. The protein that each gene encodes is given in parentheses. The size of each circle represents the percentage of cells in the group (PercExp) in which the gene expression is detected. The color represents the average transcript count in expressing cells (AvgExp). BC, bipolar cell; RB, rod bipolar cell. B, Co-expression of EAAT2 and EAAT5 in cone and rod photoreceptors.
- Figure 2.
Co-expression of EAAT2 and EAAT5 in mouse retinal RBs is confirmed by scRT-PCR analysis. A, scRT-PCR analyses of EAAT2 and EAAT5 mRNA expression in a single RB cell from a P17 mouse retina and the other RB from an adult mouse retina. Co-expression of EAAT2 and EAAT5 could be seen in both RBs. A ladder with DNA fragments between 100 and 1000 bp is shown on the left. B, The percentages of EAAT2 and EAAT5 expression in individual RBs from both P17 and adult mice. See also Table 2. C, Schematic diagrams illustrating the molecular heterogeneity for EAAT2 and EAAT5 expression in individual RBs from P17 and adult mice. The percentages of either subtype and a combination of EAAT2 and EAAT5 are shown, and the exact cell numbers are given in parentheses. See also Table 3.
- Figure 3.
EAAT2 is located near ribbons in the axon terminals of RBs. A, Confocal images showing immunofluorescence triple labeling of EAAT2 (green), RIBEYE (magenta), and PKCα (blue) in a frozen mouse retinal section. EAAT2 was expressed strongly in the OPL, and moderately in the INL, IPL, and GCL. Note that, in the INL, EAAT2 was expressed in the somata of some cone bipolar cells but not RB cells labeled by PKCα (asterisks). ONL, outer nuclear layer; OPL, outer plexiform layer; INL, inner nuclear layer; IPL, inner plexiform layer; GCL, ganglion cell layer. Scale bar: 20 μm. B, Magnification of the images in the dashed line frames of A. EAAT2 was expressed in RB axon terminals at sites near ribbons (arrows). Scale bar: 2.5 μm.
- Figure 4.
EAAT2 regulates signal transmission at RB→AII ribbon synapses. A, A schematic diagram showing the optogenetic study of neurotransmission between RBs and AII amacrine cells. ChR2 was expressed predominantly in RBs by cre-dependent recombination in adult Pcp2-cre::Ai32 mouse retinas. With all the synaptic transmission between photoreceptors and BCs is blocked pharmacologically, brief flashes of 470-nm LED light can directly activate ChR2+ RBs and induce postsynaptic responses in AIIs, which mainly reflect neurotransmitter release from RBs. The electrical coupling between ChR2+ ON cone bipolar cells and AIIs is negligible under this experimental condition (Liang et al., 2021). R, rod. B, The EPSCs recorded in AIIs, which were evoked by 470-nm LED light stimulation, were enhanced by 200 μm DHK, a selective EAAT2 blocker. Vhold = −80 mV. C, The ChR2-evoked EPSCs were reduced by 10 μm GT949, a positive allosteric modulator of EAAT2. D, E, Summary data showing the effects of GT949 (n = 10) and DHK (n = 7) on the peak amplitude of AII EPSCs. F, DHK reduced the time to peak of EPSCs slightly, but not significantly (n = 7, p = 0.0531). G, DHK reduced the rise time of EPSCs (n = 7). H, DHK did not change the decay time (tau) of EPSCs (n = 7). I–L, DHK did not affect the frequency, amplitude, rise time, or tau of mEPSCs recorded in AIIs (n = 7). mEPSCs, miniature EPSCs. M, The voltage changes in ChR2+ RBs, which were evoked by brief flashes of 470-nm LED light, were increased by 200 μm DHK. N, DHK increased the voltage changes in RBs evoked by light flashes (n = 6). O, DHK did not influence the resting membrane potentials of RBs (n = 6). The data were represented as mean ± SEM. Wilcoxon signed-rank test or Student’s t test was used where appropriate. *p < 0.05, **p < 0.01; ns, not significantly different. See also Table 4.
- Figure 5.
Pharmacological blockade of all EAATs has a significant effect on signal transmission at RB→AII synapses. A, The EPSCs recorded in AII amacrine cells, which were evoked by activating ChR2+ RBs with 470-nm LED light stimulation, were enhanced by 50 μm TBOA, a nonselective blocker of all EAATs. Vhold = −80 mV. R, rod; RB, rod bipolar cell; ChR2, channelrhodopsin-2. B, TBOA increased the peak amplitude of ChR2-evoked EPSCs (n = 7). C, The relative effects of TBOA on the peak amplitude and current integral of AII EPSCs (n = 7). The peak amplitudes/integrals were normalized to the peak amplitude/integral under control condition in each cell before averaging across cells. D, Comparison of the relative effects of DHK (n = 7), a selective EAAT2 blocker, and TBOA (n = 7) on the peak amplitude of AII EPSCs. E–G, TBOA changed the time to peak, rise time and tau of EPSCs (n = 7). H–K, TBOA did not influence the frequency, amplitude, or rise time of mEPSCs recorded in AIIs while increasing the tau slightly (n = 7). mEPSCs, miniature EPSCs. L, The voltage changes in ChR2+ RBs, which were evoked by brief flashes of 470-nm LED light, were increased by 50 μm TBOA. Note that, in the presence of TBOA, a large, long-lasting AHP (arrow) could be recorded in each RB following the light-evoked depolarization. M, TBOA increased the initial voltage changes in RBs evoked by light flashes (n = 5). N, Comparison of the relative effects of DHK (n = 6) and TBOA (n = 5) on light-evoked voltage changes in RBs. The data were represented as mean ± SEM. Wilcoxon signed-rank test or Student’s t test was used where appropriate. *p < 0.05, **p < 0.01, ***p < 0.001; ns, not significantly different. See also Table 5.
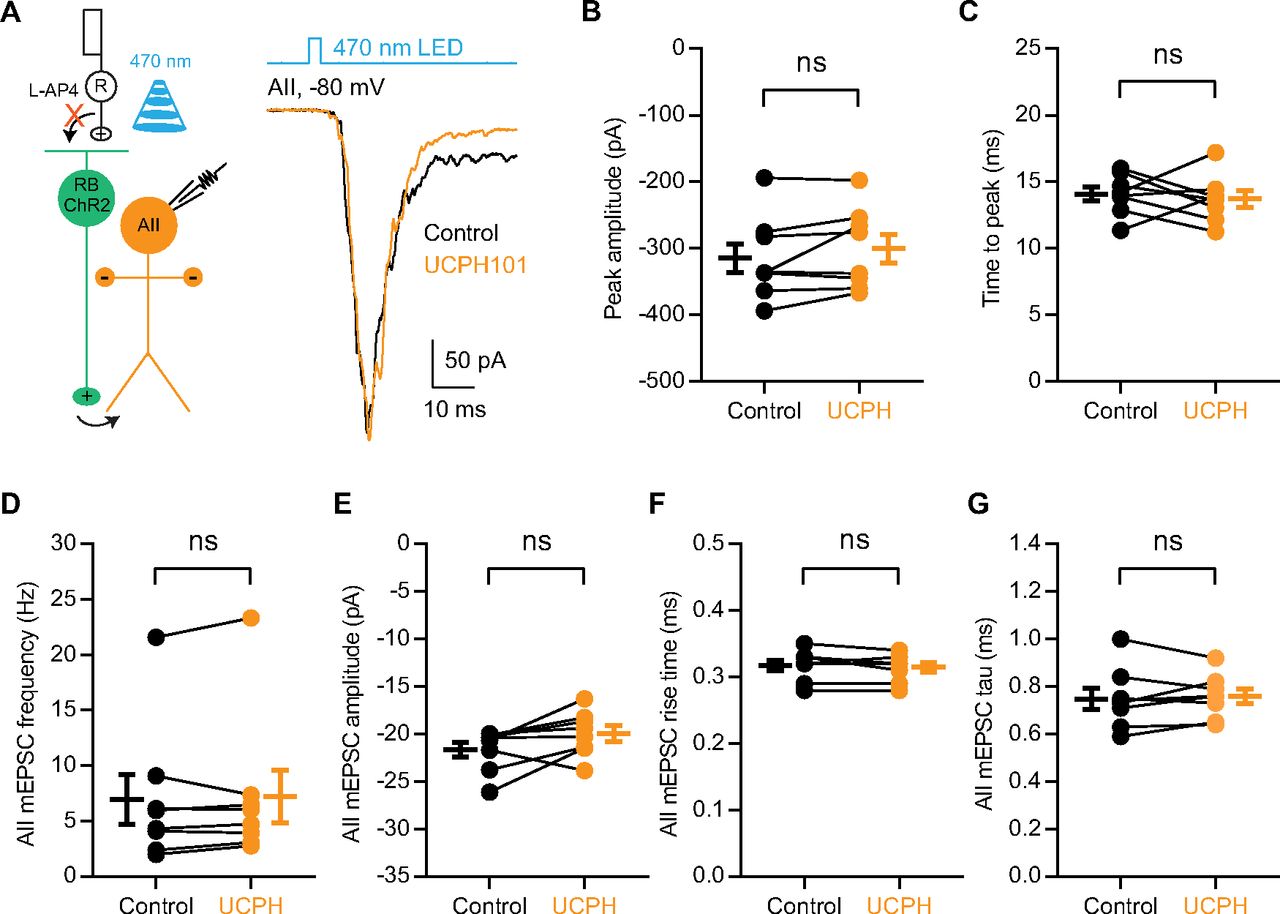
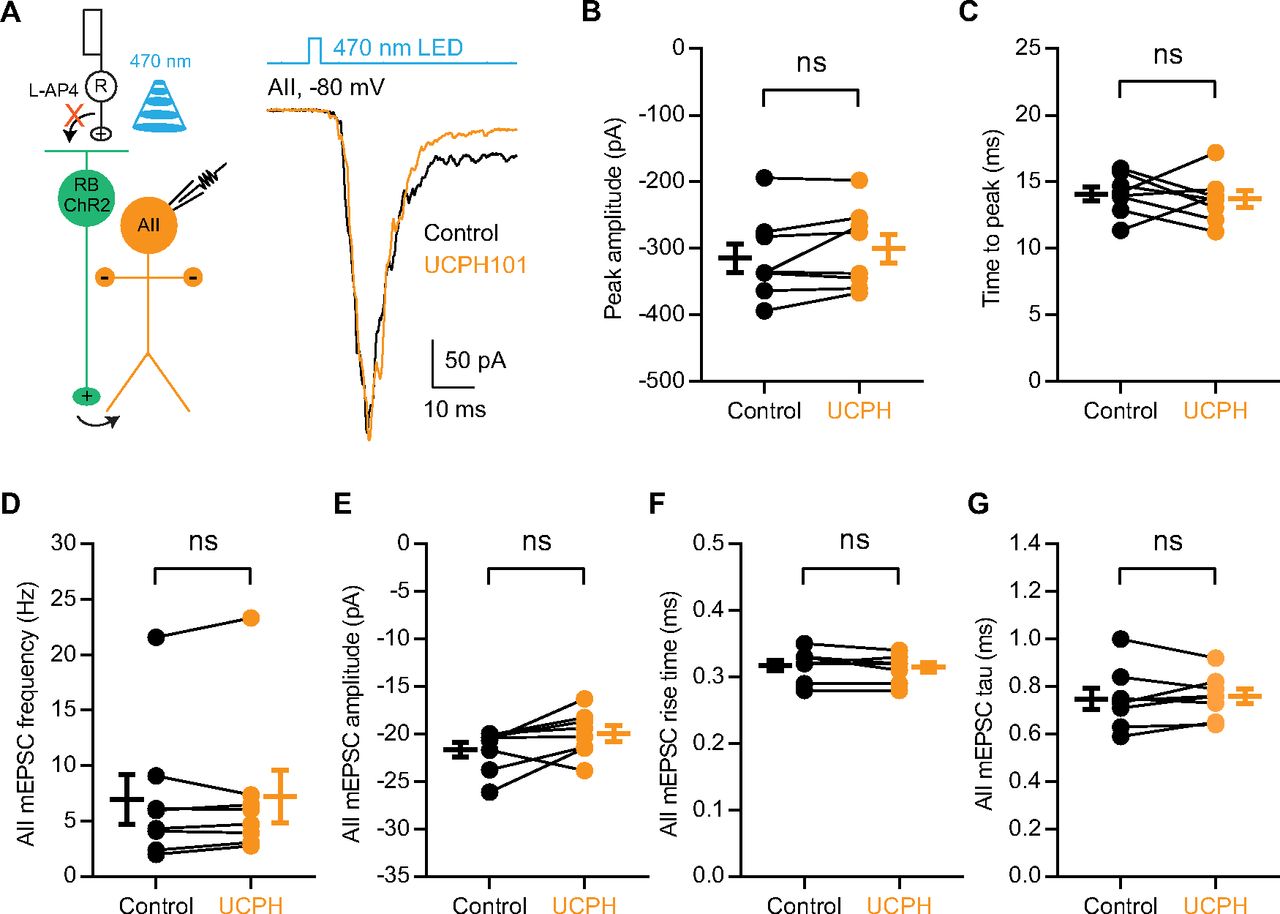
- Figure 6.
EAAT1 in Müller cells does not influence neurotransmission at RB→AII synapses. A, The EPSCs recorded in AII amacrine cells, which were evoked by activating ChR2+ RBs with 470-nm LED light stimulation, were not affected by 50 μm UCPH101, a selective blocker of EAAT1 expressed exclusively in Müller cells. Vhold = −80 mV. R, rod; RB, rod bipolar cell; ChR2, channelrhodopsin-2. B, C, UCPH101 did not change the peak amplitude or time to peak of ChR2-evoked EPSCs (n = 8). D–G, UCPH101 did not influence the frequency, amplitude, rise time, or tau of mEPSCs recorded in AIIs (n = 8). mEPSCs, miniature EPSCs. The data were represented as mean ± SEM. Wilcoxon signed-rank test or Student’s t test was used where appropriate. ns, not significantly different. See also Table 6.
- Figure 7.
EAAT5 plays a predominant role in regulating neurotransmission at RB→AII synapses. A, The EPSCs recorded in AII amacrine cells, which were evoked by 470-nm LED light stimulation of ChR2-expressing RBs, were increased slightly by co-application of DHK (200 μm) and UCPH101 (50 μm; n = 8), selective blockers of EAAT2 and EAAT1, respectively, and then enhanced more strongly by application of TBOA (50 μm; n = 3), a nonselective blocker of all EAATs. Vhold = −80 mV. R, rod; RB, rod bipolar cell; ChR2, channelrhodopsin-2. B, Magnification of the traces shown in A.
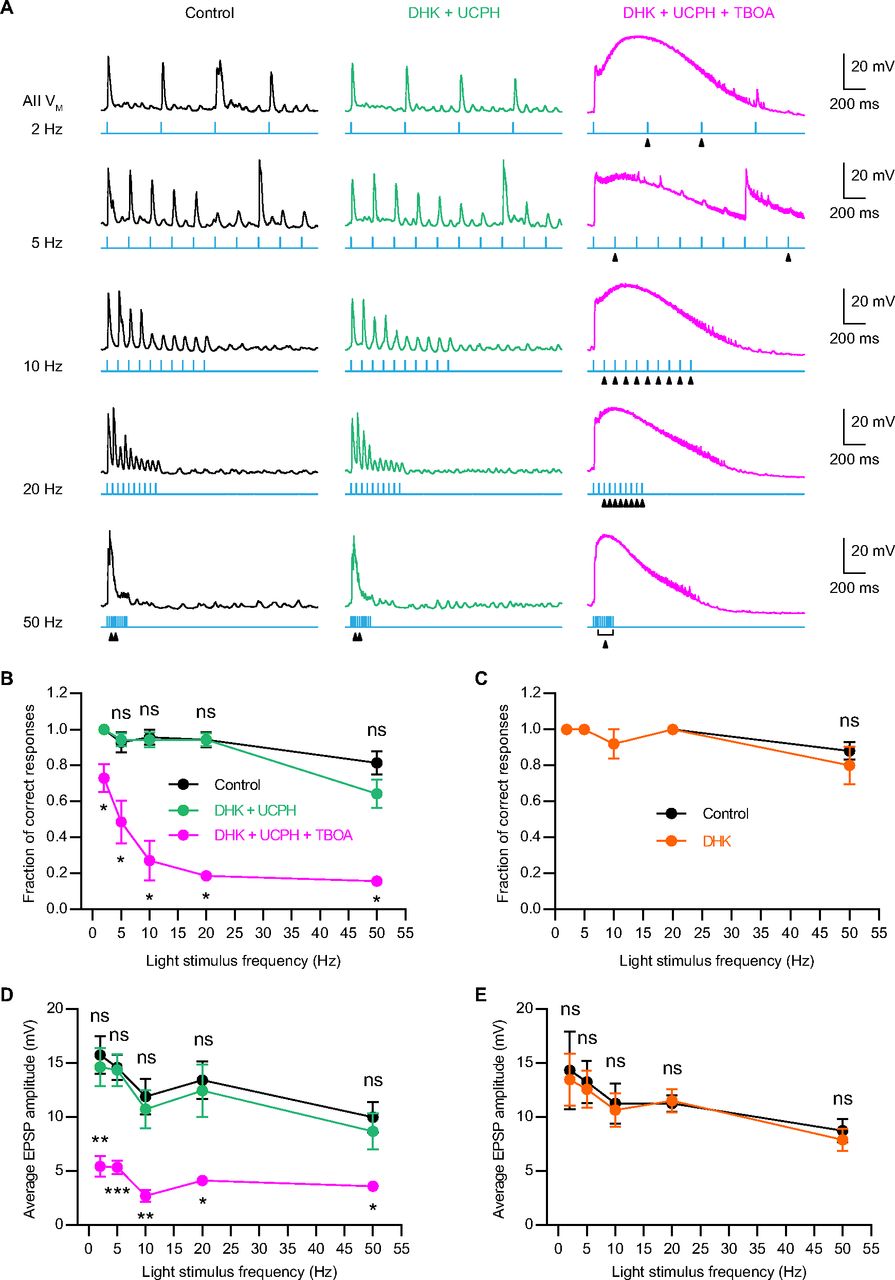
- Figure 8.
Blockade of presynaptic EAAT5 but not EAAT2 reduces temporal resolution at RB→AII synapses. A, Representative traces showing the EPSPs recorded in AII amacrine cells, which were evoked by activating ChR2-expressing RBs with 470-nm LED light stimulation. The frequencies of light stimulation were in the range of 2–50 Hz. Under the control condition, the membrane potentials of AIIs could follow the 10 consecutive flashes very well even at stimulus frequency as high as 25 Hz (left panel). Co-application of 50 μm UCPH101, a selective EAAT1 blocker, and 200 μm DHK, a selective EAAT2 blocker, did not significantly influence AIIs’ responses to flashes (middle panel). But in the presence of 50 μm TBOA, a nonselective blocker of EAATs, AIIs failed to response to some individual flashes (marked by triangles; right panel), especially at stimulus frequencies higher than 10 Hz. B, Summary data showing the fractions of correct responses for AIIs under three different experimental conditions. The fraction of correct responses was plotted as a function of light stimulus frequency. Application of 50 μm TBOA significantly reduced the fraction of correct responses at various stimulus frequencies (n = 7). C, Summary data showing the fractions of correct responses for AIIs under control and DHK conditions. Application of 200 μm DHK did not significantly change the fraction of correct responses (n = 5). D, Summary data showing the average amplitudes of AII EPSPs under three different experimental conditions. The average EPSP amplitude was plotted as a function of light stimulus frequency. Application of 50 μm TBOA significantly reduced the average EPSP amplitude at various stimulus frequencies (n = 7). E, Summary data showing the average amplitudes of AII EPSPs under control and DHK conditions. Application of 200 μm DHK did not significantly change the average EPSP amplitude (n = 5). The data were represented as mean ± SEM. Wilcoxon signed-rank test or paired t test was used for comparison. *p < 0.05, **p < 0.01, ***p < 0.001; ns, not significantly different. See also Tables 7 and 8.






Tables
Gene Protein Forward primer (5′—3′) Reverse primer (5′—3′) Slc1a2 EAAT2 CTGATGTGGTCATGTTGATAGCC AACTGGAGATGATAAGAGGGAGG Slc1a7 EAAT5 TGGCATACTACCTGTGGACTAC CTTGGTGCGGTACTGTTTGAA Prkca PKCα GTTTACCCGGCCAACGACT GGGCGATGAATTTGTGGTCTT EAAT2, excitatory amino acid transporter 2; EAAT5, excitatory amino acid transporter 5; PKCα, protein kinase C α.
EAAT2 EAAT5 P17 mice Total RB number 37 37 Positive RB number 30 26 Negative RB number 7 11 Percentage of expression 81.08% 70.27% Adult mice Total RB number 32 32 Positive RB number 28 21 Negative RB number 4 11 Percentage of expression 87.50% 65.63% EAAT2, excitatory amino acid transporter 2; EAAT5, excitatory amino acid transporter 5; P17, postnatal day 17; RB, rod bipolar cell.
P17 mice Adult mice RB number Percentage RB number Percentage EAAT2 alone 7 21.21% 8 27.59% EAAT5 alone 3 9.09% 1 3.45% EAAT2 and EAAT5 23 69.70% 20 68.97% Total 33 29 EAAT2, excitatory amino acid transporter 2; EAAT5, excitatory amino acid transporter 5; P17, postnatal day 17; RB, rod bipolar cell.
Data Data structure Type of test Power Mean ± SEM Number
of cellsEffect of DHK on AII EPSC amplitude (unit: pA) Control Normal distribution −295.40 ± 24.45 7 200 μm DHK Normal distribution −350.70 ± 36.26 7 a Control vs DHK Paired Student’s t test p = 0.0111 7 Effect of GT949 on AII EPSC amplitude (unit: pA) Control Normal distribution −347.50 ± 24.07 10 10 μm GT949 Normal distribution −274.40 ± 24.73 10 b Control vs GT949 Paired Student’s t test p = 0.0004 10 Relative effects of DHK and GT949 on AII EPSC amplitude Control Non-normal distribution 1.00 ± 0.00 200 μm DHK Normal distribution 1.18 ± 0.04 7 10 μm GT949 Normal distribution 0.78 ± 0.03 10 c Control vs DHK Wilcoxon signed-rank test p = 0.0313 7 d Control vs GT949 Wilcoxon signed-rank test p = 0.0020 10 Effect of DHK on AII EPSC time to peak (unit: ms) Control Normal distribution 12.53 ± 0.44 7 200 μm DHK Normal distribution 11.87 ± 0.44 7 e Control vs DHK Paired Student’s t test p = 0.0531 7 Effect of DHK on AII EPSC rise time (unit: ms) Control Normal distribution 3.98 ± 0.43 7 200 μm DHK Normal distribution 3.09 ± 0.48 7 f Control vs DHK Paired Student’s t test p = 0.0050 7 Effect of DHK on AII EPSC tau (unit: ms) Control Normal distribution 6.73 ± 0.80 7 200 μm DHK Normal distribution 6.03 ± 0.58 7 g Control vs DHK Paired Student’s t test p = 0.3519 7 Effect of DHK on AII mEPSC frequency (unit: Hz) Control Normal distribution 8.25 ± 1.20 7 200 μm DHK Normal distribution 8.97 ± 1.07 7 h Control vs DHK Paired Student’s t test p = 0.1120 7 Effect of DHK on AII mEPSC amplitude (unit: pA) Control Normal distribution −19.47 ± 1.30 7 200 μm DHK Normal distribution −19.23 ± 0.95 7 i Control vs DHK Paired Student’s t test p = 0.6892 7 Effect of DHK on AII mEPSC rise time (unit: ms) Control Normal distribution 0.33 ± 0.01 7 200 μm DHK Non-normal distribution 0.34 ± 0.00 7 j Control vs DHK Wilcoxon signed-rank test p = 0.3750 7 Effect of DHK on AII mEPSC tau (unit: ms) Control Non-normal distribution 0.88 ± 0.06 7 200 μm DHK Normal distribution 0.90 ± 0.03 7 k Control vs DHK Wilcoxon signed-rank test p = 0.5781 7 Effect of DHK on RB light-evoked voltage change (unit: mV) Control Normal distribution 9.62 ± 0.39 6 200 μm DHK Normal distribution 11.14 ± 0.56 6 l Control vs DHK Paired Student’s t test p = 0.0036 6 Effect of DHK on RB resting membrane potential (unit: mV) Control Normal distribution −55.28 ± 1.04 6 200 μm DHK Normal distribution −57.21 ± 1.76 6 m Control vs DHK Paired Student’s t test p = 0.1574 6 DHK, a selective EAAT2 blocker; GT949, a positive allosteric modulator of EAAT2; AII, AII amacrine cell; RB, rod bipolar cell.
Data Data structure Type of test Power Mean ± SEM Number
of cellsEffect of TBOA on AII EPSC amplitude (unit: pA) Control Normal distribution −237.40 ± 16.21 7 50 μm TBOA Non-normal distribution −547.20 ± 61.50 7 a Control vs TBOA Wilcoxon signed-rank test p = 0.0156 7 Relative effects of TBOA on AII EPSC amplitude and integral Control Non-normal distribution 1.00 ± 0.00 7 50 μm TBOA (amplitude) Normal distribution 2.31 ± 0.20 7 50 μm TBOA (integral) Non-normal distribution 11.22 ± 1.70 7 b Control vs TBOA (amplitude) Wilcoxon signed-rank test p = 0.0156 7 c Control vs TBOA (integral) Wilcoxon signed-rank test p = 0.0156 7 d TBOA (amplitude) vs TBOA (integral) Wilcoxon signed-rank test p = 0.0156 7 Relative effect of DHK on AII EPSC amplitude 200 μm DHK Normal distribution 1.18 ± 0.04 7 e DHK vs TBOA (amplitude) Unpaired Student’s t test p = 0.0001 Effect of TBOA on AII EPSC time to peak (unit: ms) Control Normal distribution 14.16 ± 0.62 7 50 μm TBOA Normal distribution 26.21 ± 2.12 7 f Control vs TBOA Paired Student’s t test p = 0.0004 7 Effect of TBOA on AII EPSC rise time (unit: ms) Control Normal distribution 3.34 ± 0.75 7 50 μm TBOA Normal distribution 10.23 ± 1.99 7 g Control vs TBOA Paired Student’s t test p = 0.0032 7 Effect of TBOA on AII EPSC tau (unit: ms) Control Non-normal distribution 64.54 ± 20.61 7 50 μm TBOA Normal distribution 325 ± 68.02 7 h Control vs TBOA Paired Student’s t test p = 0.0050 7 Effect of TBOA on AII mEPSC frequency (unit: Hz) Control Normal distribution 6.19 ± 1.14 7 50 μm TBOA Normal distribution 6.19 ± 1.19 7 i Control vs TBOA Paired Student’s t test p = 0.9854 7 Effect of TBOA on AII mEPSC amplitude (unit: pA) Control Normal distribution −19.98 ± 1.55 7 50 μm TBOA Normal distribution −20.13 ± 1.57 7 j Control vs TBOA Paired Student’s t test p = 0.8856 7 Effect of TBOA on AII mEPSC rise time (unit: ms) Control Non-normal distribution 0.32 ± 0.01 7 50 μm TBOA Normal distribution 0.32 ± 0.01 7 k Control vs TBOA Wilcoxon signed-rank test p > 0.9999 7 Effect of TBOA on AII mEPSC tau (unit: ms) Control Normal distribution 0.75 ± 0.04 7 50 μm TBOA Normal distribution 0.81 ± 0.06 7 l Control vs TBOA Paired Student’s t test p = 0.0392 7 Effect of TBOA on RB light-evoked voltage change (unit: mV) Control Normal distribution 8.69 ± 0.39 5 50 μm TBOA Normal distribution 9.64 ± 0.51 5 m Control vs TBOA Paired Student’s t test p = 0.0192 5 Effect of TBOA on RB resting membrane potential (unit: mV) Control Normal distribution −52.92 ± 2.01 5 50 μm TBOA Normal distribution −52.91 ± 2.95 5 n Control vs TBOA Paired Student’s t test p = 0.9931 5 Relative effects of DHK and TBOA on RB light-evoked voltage change 200 μm DHK Normal distribution 1.16 ± 0.03 6 50 μm TBOA Normal distribution 1.11 ± 0.03 5 o DHK vs TBOA Unpaired Student’s t test p = 0.3226 TBOA, a non-selective EAAT blocker; DHK, a selective EAAT2 blocker; AII, AII amacrine cell; RB, rod bipolar cell.
Data Data structure Type of test Power Mean ± SEM Number
of cellsEffect of UCPH on AII EPSC amplitude Control Normal distribution −315.00 ± 22.05 8 50 μm UCPH Normal distribution −300.60 ± 21.52 8 a Control vs UCPH Paired Student’s t test p = 0.1588 8 Effect of UCPH on AII EPSC time to peak (unit: ms) Control Normal distribution 14.08 ± 0.53 8 50 μm UCPH Normal distribution 13.69 ± 0.63 8 b Control vs UCPH Paired Student’s t test p = 0.6306 8 Effect of UCPH on AII mEPSC frequency (unit: Hz) Control Normal distribution 6.95 ± 2.24 8 50 μm UCPH Normal distribution 7.22 ± 2.37 8 c Control vs UCPH Paired Student’s t test p = 0.4685 8 DHK effect on AII mEPSC amplitude (unit: pA) Control Normal distribution −21.62 ± 0.78 8 50 μm UCPH Normal distribution −19.94 ± 0.82 8 d Control vs UCPH Paired Student’s t test p = 0.0707 8 Effect of UCPH on AII mEPSC rise time (unit: ms) Control Non-normal distribution 0.32 ± 0.01 8 50 μm UCPH Normal distribution 0.32 ± 0.01 8 e Control vs UCPH Wilcoxon signed-rank test p = 0.8125 8 Effect of UCPH on AII mEPSC tau (unit: ms) Control Normal distribution 0.75 ± 0.04 8 50 μm UCPH Normal distribution 0.76 ± 0.03 8 f Control vs UCPH Paired Student’s t test p = 0.6382 8 UCPH101, a selective EAAT1/GLAST blocker; AII, AII amacrine cell.
- Table 7
The effects of EAAT-related drugs on the fraction of correct responses in AIIs at various light stimulus frequencies
Data Data structure Type of test Power Mean ± SEM Number
of cellsEffects of DHK, UCPH101 and TBOA on the fraction of correct responses in AIIs at various stimulus frequencies 2-Hz stimulus frequency Control Non-normal distribution 1.00 ± 0.00 7 200 μm DHK +
50 μm UCPH101Non-normal distribution 1.00 ± 0.00 7 200 μm DHK +
50 μm UCPH101 + 50 μm TBOANormal distribution 0.73 ± 0.08 7 a Control vs
DHK + UCPHWilcoxon signed-rank test N/A 7 b DHK + UCPH vs DHK + UCPH + TBOA Wilcoxon signed-rank test p = 0.0313 7 5-Hz stimulus frequency Control Non-normal distribution 0.93 ± 0.06 7 200 μm DHK +
50 μm UCPH101Non-normal distribution 0.94 ± 0.04 7 200 μm DHK +
50 μm UCPH101 + 50 μm TBOANormal distribution 0.49 ± 0.12 7 c Control vs
DHK + UCPHWilcoxon signed-rank test p > 0.9999 7 d DHK + UCPH vs DHK + UCPH + TBOA Wilcoxon signed-rank test p = 0.0313 7 10-Hz stimulus frequency Control Non-normal distribution 0.96 ± 0.04 7 200 μm DHK +
50 μm UCPH101Non-normal distribution 0.94 ± 0.04 7 200 μm DHK +
50 μm UCPH101 + 50 μm TBOANon-normal distribution 0.27 ± 0.11 7 e Control vs
DHK + UCPHWilcoxon signed-rank test p > 0.9999 7 f DHK + UCPH vs DHK + UCPH + TBOA Wilcoxon signed-rank test p = 0.0156 7 20-Hz stimulus frequency Control Non-normal distribution 0.94 ± 0.04 7 200 μm DHK +
50 μm UCPH101Non-normal distribution 0.94 ± 0.04 7 200 μm DHK +
50 μm UCPH101 + 50 μm TBOANormal distribution 0.19 ± 0.03 7 g Control vs
DHK + UCPHWilcoxon signed-rank test p > 0.9999 7 h DHK + UCPH vs DHK + UCPH + TBOA Wilcoxon signed-rank test p = 0.0156 7 50-Hz stimulus frequency Control Normal distribution 0.81 ± 0.06 7 200 μm DHK +
50 μm UCPH101Normal distribution 0.64 ± 0.08 7 200 μm DHK +
50 μm UCPH101 + 50 μm TBOANon-normal distribution 0.16 ± 0.03 7 i Control vs
DHK + UCPHPaired t test p = 0.0533 7 j DHK + UCPH vs DHK + UCPH + TBOA Wilcoxon signed-rank test p = 0.0156 7 Effects of DHK alone on the fraction of correct responses in AIIs at various stimulus frequencies 2-Hz stimulus frequency Control Non-normal distribution 1.00 ± 0.00 5 200 μm DHK Non-normal distribution 1.00 ± 0.00 5 k Control vs DHK Wilcoxon signed-rank test N/A 5 5-Hz stimulus frequency Control Non-normal distribution 1.00 ± 0.00 5 200 μm DHK Non-normal distribution 1.00 ± 0.00 5 l Control vs DHK Wilcoxon signed-rank test N/A 5 10-Hz stimulus frequency Control Non-normal distribution 0.92 ± 0.08 5 200 μm DHK Non-normal distribution 0.92 ± 0.08 5 m Control vs DHK Wilcoxon signed-rank test N/A 5 20-Hz stimulus frequency Control Non-normal distribution 1.00 ± 0.00 5 200 μm DHK Non-normal distribution 1.00 ± 0.00 5 n Control vs DHK Wilcoxon signed-rank test N/A 5 50-Hz stimulus frequency Control Non-normal distribution 0.88 ± 0.05 5 200 μm DHK Normal distribution 0.80 ± 0.10 5 o Control vs DHK Wilcoxon signed-rank test p = 0.5000 5 TBOA, a non-selective EAAT blocker; UCPH101, a selective EAAT1/GLAST blocker; DHK, a selective EAAT2 blocker; AII, AII amacrine cell; N/A, not applicable.
- Table 8
The effects of EAAT-related drugs on the average amplitude of AII EPSPs at various light stimulus frequencies
Data Data structure Type of test Power Mean ± SEM Number
of cellsEffects of DHK, UCPH101 and TBOA on the average amplitude of AII EPSPs at various stimulus frequencies 2-Hz stimulus frequency Control Normal distribution 15.75 ± 1.77 7 200 μm DHK +
50 μm UCPH101Normal distribution 14.62 ± 1.77 7 200 μm DHK +
50 μm UCPH101 + 50 μm TBOANormal distribution 5.44 ± 0.97 7 a Control vs
DHK + UCPHPaired t test p = 0.1704 7 b DHK + UCPH vs DHK + UCPH + TBOA Paired t test p = 0.0013 7 5-Hz stimulus frequency Control Normal distribution 14.58 ± 1.16 7 200 μm DHK +
50 μm UCPH101Normal distribution 14.36 ± 1.48 7 200 μm DHK +
50 μm UCPH101 + 50 μm TBOANormal distribution 5.35 ± 0.62 7 c Control vs
DHK + UCPHPaired t test p = 0.7810 7 d DHK + UCPH vs DHK + UCPH + TBOA Paired t test p = 0.0007 7 10-Hz stimulus frequency Control Normal distribution 11.90 ± 1.66 7 200 μm DHK +
50 μm UCPH101Normal distribution 10.74 ± 1.79 7 200 μm DHK +
50 μm UCPH101 + 50 μm TBOANormal distribution 2.70 ± 0.52 7 e Control vs
DHK + UCPHPaired t test p = 0.0746 7 f DHK + UCPH vs DHK + UCPH + TBOA Paired t test p = 0.0054 7 20-Hz stimulus frequency Control Normal distribution 13.41 ± 1.74 7 200 μm DHK +
50 μm UCPH101Normal distribution 12.45 ± 2.42 7 200 μm DHK +
50 μm UCPH101 + 50 μm TBOANormal distribution 4.12 ± 0.29 7 g Control vs
DHK + UCPHPaired t test p = 0.4964 7 h DHK + UCPH vs DHK + UCPH + TBOA Paired t test p = 0.0116 7 50-Hz stimulus frequency Control Normal distribution 9.99 ± 1.42 7 200 μm DHK +
50 μm UCPH101Normal distribution 8.67 ± 1.65 7 200 μm DHK +
50 μm UCPH101 + 50 μm TBOANon-normal distribution 3.58 ± 0.24 7 i Control vs
DHK + UCPHPaired t test p = 0.0763 7 j DHK + UCPH vs DHK + UCPH + TBOA Wilcoxon signed-rank test p = 0.0156 7 Effects of DHK alone on the average amplitude of AII EPSPs at various stimulus frequencies 2-Hz stimulus frequency Control Normal distribution 14.33 ± 3.61 5 200 μm DHK Normal distribution 13.46 ± 2.41 5 k Control vs DHK Paired t test p = 0.6330 5 5-Hz stimulus frequency Control Normal distribution 13.25 ± 1.97 5 200 μm DHK Normal distribution 12.57 ± 1.71 5 l Control vs DHK Paired t test p = 0.5547 5 10-Hz stimulus frequency Control Normal distribution 11.26 ± 1.87 5 200 μm DHK Normal distribution 10.65 ± 1.55 5 m Control vs DHK Paired t test p = 0.4382 5 20-Hz stimulus frequency Control Normal distribution 11.26 ± 0.76 5 200 μm DHK Normal distribution 11.51 ± 1.08 5 n Control vs DHK Paired t test p = 0.8054 5 50-Hz stimulus frequency Control Normal distribution 8.74 ± 1.07 5 200 μm DHK Normal distribution 7.88 ± 1.05 5 o Control vs DHK Paired t test p = 0.0780 5 TBOA, a non-selective EAAT blocker; UCPH101, a selective EAAT1/GLAST blocker; DHK, a selective EAAT2 blocker; AII, AII amacrine cell.
In this issue

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}



