

Abstract
Cognitive flexibility, attributed to frontal cortex, is vital for navigating the complexities of everyday life. The mediodorsal thalamus (MD), interconnected to frontal cortex, may influence cognitive flexibility. Here, male rats performed an attentional set-shifting task measuring intradimensional (ID) and extradimensional (ED) shifts in sensory discriminations. MD lesion rats needed more trials to learn the rewarded sensory dimension. However, once the choice response strategy was established, learning further two-choice discriminations in the same sensory dimension, and reversals of the reward contingencies in the same dimension, were unimpaired. Critically though, MD lesion rats were impaired during the ED shift, when they must rapidly update the optimal choice response strategy. Behavioral analyses showed MD lesion rats had significantly reduced correct within-trial second choice responses. This evidence shows that transfer of information via the MD is critical when rapid within-trial updates in established choice response strategies are required after a rule change.
- anterior cingulate
- cognitive control
- cognitive flexibility
- decision making
- medial prefrontal cortex
- orbitofrontal cortex
Significance Statement
We demonstrate for the first time that rodent mediodorsal thalamus (MD) is a critical node when choice response strategies need to change rapidly after a within-session rule change during reward-guided learning. MD interactions with orbitofrontal cortex (OFC) are critical for value-based learning, while MD interactions with medial prefrontal cortex (PFC) are critical for rapid within-trial updating of optimal choice response rules. MD interactions with the OFC are not always necessary for reversal learning. These results indicate that the MD also contributes to attentional set-shifting. Our evidence advocates for studies investigating deficits in updating optimal choice response strategies because of potentially disrupted cortico-thalamocortical transfer of information in people diagnosed with Alzheimer’s disease (AD) combined with more frontal pathology, Parkinson’s disease (PD), or schizophrenia.
Introduction
Cognitive flexibility describes our ability to quickly and selectively switch our thoughts, responses, or behavior to everyday dynamic situations. This capacity to rapidly update or alter one’s actions conveys evolutionary benefit and is key to survival (Koechlin et al., 2003; Rougier et al., 2005). Cognitive flexibility shows marked changes or declines in neurodegenerative disorders, like Parkinson’s disease (PD), in patients with ventral prefrontal cortex (PFC) damage, and in neurodevelopmental diseases, like schizophrenia (Clark et al., 2004; Andrews et al., 2006; Barch et al., 2009; Giraldo-Chica et al., 2018).
Cognitive flexibility is not unique to primates. Studies show that rats can readily switch between attentional sets to optimize reward outcome (Birrell and Brown, 2000; McAlonan and Brown, 2003; Newman and McGaughy, 2011). Typically, these studies have focused on various frontal cortex subregions. Yet, it is becoming increasingly clear that the cortex does not function in isolation, but rather, relies heavily on subcortical and peripheral inputs supplied by the thalamus (Sherman and Guillery, 2002; Guillery and Sherman, 2011; Halassa and Kastner, 2017). The mediodorsal thalamus (MD) has a critical role in functions of frontal cortex during higher order cognitive processes across mammalian species (Mitchell and Dalrymple-Alford, 2005; Mitchell et al., 2007a, b; Mitchell and Gaffan, 2008; Parnaudeau et al., 2013; Browning et al., 2015; Chakraborty et al., 2016, 2019; Miller et al., 2017; Schmitt et al., 2017; Alcaraz et al., 2018; Ferguson and Gao, 2018; Pergola et al., 2018; DeNicola et al., 2020; Perry et al., 2021). This evidence highlights that the MD and PFC are working in partnership, contributing different but complementary roles to goal-directed behaviors (Mitchell, 2015; Perry et al., 2021).
In mammals, the MD is densely interconnected with areas of frontal cortex involved in various aspects of cognitive flexibility. Recent work shows MD projections to frontal cortex allow for the simultaneous sharing of information across multiple cortical regions. This distributed pattern of frontal innervation by the MD appears to be especially true for the central (MDc) and medial (MDm) subdivisions, which have projections to the orbitofrontal cortex (OFC), medial PFC, and frontal association areas (Groenewegen, 1988; Dajani and Uddin, 2015; Alcaraz et al., 2016; Kuramoto et al., 2017; Perry et al., 2021). Although phylogenetic differences exist in the organization of rodent and primate frontal cortex (Preuss, 1995), similar functional contributions to aspects of cognitive flexibility are observed in monkeys and rodents after perturbations to similar areas of agranular frontal cortex (Dias et al., 1996, 1997; Birrell and Brown, 2000; McAlonan and Brown, 2003; Bissonette et al., 2008). Given there are similarities in neuroanatomical connectivity between agranular frontal cortex and MD in rodents and primates, it seems that cognitive flexibility and rapid updating deficits observed in monkeys with excitotoxic MD lesions (Mitchell et al., 2007a; Mitchell and Gaffan, 2008; Browning et al., 2015; Chakraborty et al., 2016) may also extend to rodents.
Thus, in the current study, rats with excitotoxic lesions to the MD or MD sham controls were run on a well-established test of cognitive flexibility, the intradimensional (ID)/extradimensional (ED) attentional set-shifting task (Birrell and Brown, 2000). To test this premise and investigate any deficits in choice responding, we used the standard 7-stage version comprising of multiple subtasks all conducted within a single testing session (Birrell and Brown, 2000; Chase et al., 2012). In rats, the ID/ED task has produced dissociable deficits after permanent cortical or limbic thalamic lesions, or neuropharmacological manipulations (McGaughy et al., 2008; Tait et al., 2014, 2018; Wright et al., 2015; Linley et al., 2016).
In the current experiment, we predicted MD lesion rats would show deficits during the initial sensory acquisition (simple discrimination; SD) and the ED shift. Both the SD and ED subtasks require rapid, within trial learning or updating of choice response strategies. Therefore, impaired SD and ED performance would be consistent with the selective learning deficits observed in other rodent studies after MD perturbations (Mitchell and Chakraborty, 2013; for review, see Hunt and Aggleton, 1998; Mitchell and Dalrymple-Alford, 2005; Ferguson and Gao, 2018; Courtiol et al., 2019; Perry et al., 2021). In addition, we predicted our MD lesion rats may show deficits during reversal subtasks. MD lesions in rodents can produce reversal learning deficits, although the evidence is mixed (for review, see Mitchell and Chakraborty, 2013). However, given the reciprocal direct MD connectivity with the OFC (Krettek and Price, 1977; Groenewegen, 1988; Ray and Price, 1992), and the selective deficits observed in rodents and monkeys with OFC manipulations performing the reversal subtasks during the ID/ED attentional set-shifting task (Dias et al., 1996; McAlonan and Brown, 2003), it remained an open question.
Materials and Methods
Animals
Twenty-five Lister hooded male rats weighing between 420 and 480 g at time of surgery (Charles River) were group housed in a temperature and humidity-controlled environment (21 ± 1°C). The housing and husbandry compiled with the ARRIVE guidelines of the European Directive (2010/63/EU) for the care and use of laboratory animals. Testing was conducted in the light phase of a 12/12 h light/dark cycle (lights on at 7 A.M.s). The rats were maintained on a controlled feeding schedule (20 g/rat/d) with water freely available in the home cage. All experimental procedures were performed in compliance with the United Kingdom Animals (Scientific Procedures) Act of 1986. A project license authorized all procedures after review by the university animal care and ethical review committee and Home Office Inspectorate.
Surgery
Rats were anesthetized by isoflurane and oxygen mix (4% induction, 1.8–2% maintenance), and given analgesia in subcutaneous injections of 0.05 mg/kg buprenorphine (Vetergesic; 0.3 mg/ml) and 1 mg/kg Metacam (Meloxicam; 5 mg/ml). Rats were secured in a stereotaxic frame (Kopf) with atraumatic ear bars and the nose bar set to +3.3 mm to achieve a level skull. A subcutaneous injection of 2 mg/kg bupivacaine (Marcaine; 2.5 mg/ml) was administered into the scalp in the location of the midline incision. Viscotears was applied to keep the eyes moist. During surgery, each rat was placed on a heat pad and covered in bubble wrap with an internal rectal thermometer probe to monitor and maintain normal body temperature. Warmed sterile saline (1 ml/100 g) was administered subcutaneous into the scruff of the neck after 1 h. A midline incision was performed. Bregma and λ coordinates were determined. A dental drill with a trephine head was used to create a craniotomy over the midline. To maximize lesion accuracy, anterior and posterior injection sites were calculated according to the bregma–λ distance in each rat. Coordinates were for anterior MD injection: anterior-posterior (AP) = – 0.395 mm, medial-lateral (ML) = – 0.1 (avoiding the superior sagittal sinus, which was visible within the dura inside the craniotomy), and dorsal-ventral (DV) = 0.56 mm (from dura), volume of excitotoxin = 0.20 μl; for posterior MD: AP = – 0.435 mm, ML = + 0.1 and DV = 0.57 mm (from dura), volume of excitotoxin = 0.18 μl. Fifteen rats (MD lesion) received 0.12 m NMDA dissolved in phosphate buffer (pH 7.20) in each hemisphere from a 1 μl-gauge Hamilton bevelled-tip syringe. The injections were performed manually, taking 3 min each, and after injection the needle was left in situ for a further 3 min to allow diffusion and to limit wicking of the NMDA. A further 10 rats (MD sham controls) received injections of sterile phosphate buffer instead of NMDA using the same injection coordinates. Injections were given bilaterally in the MD. Upon completion of surgery, wounds were sutured with Vicryl 4.0. Rats were housed singly during recovery for up to 3 d and were then returned to their presurgery housing cohort. Postoperative oral analgesia, 1 mg/kg Metacam (Meloxicam, 1.5 mg/ml) dissolved and set in jelly was provided in individual bowls to each rat daily for 3 d. Behavioral and physiological evidence showed that all rats recovered well, with normal eating and presurgery weights returning within 24 h. Food regulation (20 g/d/rat) started again 10d after recovery. Postoperative testing began at least 15d after surgery. The researcher was blind to the lesion group of each animal until all tests were completed.
ID/ED attentional set-shifting paradigm
Apparatus
The test chamber consisted of a modified Plexiglas home cage (40 × 70 × 18 cm) with Perpex dividers separating the cage into a large start chamber (40 × 46 cm) and two identically sized (24 × 20 cm) choice chambers to which access was controlled by removable Perspex doors. The digging bowls were ceramic (internal diameter 7 cm, depth 4 cm) and were placed within the choice chambers. The bowls were filled with digging media of different textures, and the digging media were scented with different spices (see Table 1 for examples). The odor or digging media discriminations, pairs of stimuli used, and the correct stimulus within pairs were counter-balanced across subtasks and matched between groups. The bowls were baited with Honey Nut Cheerios (Nestle), each bowl contained a hidden (securely fixed under a metal grid) Honey Nut Cheerio to serve as an odor mask. During testing, only one of the bowls was baited and rats determined which bowl was baited using either the texture of the digging medium or the odor as cues. Before testing, rats were taught to dig in bowls filled with home-cage bedding material to retrieve one half of a Honey Nut Cheerio. The task was performed as previously described in Birrell and Brown (2000) and divided into two phases administered on two consecutive days.
Sensory dimensions (odors and digging media) and stimulus features (O1–O6; M1–M6) used in the different subtasks of the ID/ED attentional set-shifting task
Preoperative and postoperative training day
On the day before testing, each rat learnt one simple two-choice discrimination (SD) using either of the two sensory dimensions: odor (mint vs oregano), or digging media (shredded paper vs cotton pads), to a criterion of six consecutive correct trials. The rewarded odors or digging media were counterbalanced across the two groups, and these exemplars were not used again throughout testing. Digging was defined as active digging with both front paws or active foraging with the snout in the digging media. Sniffing or touching the media with the paws was not scored as a dig.
Preoperative and postoperative testing paradigm
The following day, each rat was given a series of seven two-choice discriminations (subtasks) in the following order: a new SD using either of two sensory dimension (odor or digging media); a compound discrimination (CD) using the same rewarded sensory dimension and two-choice discrimination as the SD, combined with the other irrelevant sensory dimension; first reversal (REV1), in which the exemplars remained the same as in the CD but the correct (rewarded) and incorrect exemplars were reversed; ID shift, in which new exemplars were used, but the relevant sensory dimension remained the same as in the previous subtasks; second reversal (REV2), where exemplars in the ID remained the same but the reward contingencies of the two exemplars were reversed; ED shift, where new exemplars were used and the previously irrelevant sensory dimension becomes relevant; and a third reversal (REV3), where exemplars in the ED subtask remained the same but the reward contingencies of the two exemplars were reversed.
The task is self-paced and relies on the natural foraging behaviors of the rats. The first five two-choice discrimination subtasks (SD, CD, REV1, ID, REV2) must be solved by discriminating between exemplars from the same sensory dimension. In this stage of the task, the rat is rewarded for choices based on specific perceptual features of the stimuli, while ignoring other features that also distinguish the stimuli. After acquiring each set of two-choice discriminations to a performance criterion of six correct consecutive trials, the rats encounter a reversal of the reward contingencies associated with the exemplars (a reversal subtask: REV1 and REV2). For the final two subtasks (ED and REV3), exemplars from the previously irrelevant sensory dimension become relevant. Therefore, the rat must shift its attention (and adjust its choice response strategy) to the previously irrelevant sensory dimension and perceptual feature to receive reward (ED subtask). The attentional set-shifting cost is calculated by comparing trials to criterion during the ID shift, where the sensory dimension is consistent with previous subtasks, and the ED shift, where the now relevant sensory dimension had been previously ignored and thus requires a shift to optimize continuing to retrieve rewards.
Odor detection test
At the completion of ID/ED testing, the olfactory abilities of the rat to smell the odor of the buried half-Honey Nut Cheerio were assessed. This task consisted of 10 consecutive trials where the rat was exposed to bowls containing similar bedding. The rat was placed in the waiting area of the set-shifting apparatus. Similar bedding-filled bowls were placed, one in each of the choice chambers, only one bowl contained half a Honey Nut Cheerio at the bottom (pseudorandomly assigned to avoid the rat developing a response strategy). The barrier was raised, allowing the rat access to both bowls. The rat had to chose to dig into one bowl and had up to 15 min to make a digging response on each trial. If the rat dug in the correct bowl, the trial was recorded as correct and the rat was returned to the waiting area and the barrier lowered to block access to both choice chambers. If the rat dug in the incorrect bowl, the trial was marked as incorrect. The rat was permitted to continue to explore the incorrect bowl; the trial was terminated when the rat returned to the waiting area. In all cases, the ability of each rat to detect the reward location was no different to chance guessing.
Perfusion and histology
At the end of all experimental testing, rats were deeply anaesthetized with sodium pentobarbital (Euthanol 1.0 ml/rat, 200 mg/ml: Merial), and perfused transcardially with 0.9% (w/v) saline and 4% (w/v) paraformaldehyde in 0.1 m PBS. Brains were then removed and postfixed overnight (4°C), then incubated using 30% (w/v) sucrose in 0.1 m PBS over 48 h (4°C). Free floating sections (40 μm) were cut on a freezing microtome, the slices were then preserved in cryoprotectant (0.1 m PBS containing 24% (v/v) glycerol and 24% (v/v) ethylene glycol) and stored at −20°C until required. MD thalamus lesion locations were histologically verified from cresyl violet-stained brain sections.
Lesion extent
Photomicrographs of the Nissl-stained MD sections were captured using a camera mounted on a freestanding Leica DMR microscope (Leica Microsystems) using a 1.6× objective so that the whole section could be visualised in the photomicrograph. MD volumes were measured using the NIH software ImageJ (http://rsbweb.nih.gov/ij/). In each rat, the intact MD volume from both the left and the right hemisphere was assessed. The final reading was calculated according to the Cavalieri principle (Regeur and Pakkenberg, 1989) and expressed as the percentage of MD sham controls.
Behavioral data analysis
In the behavioral experiments, all rats performed a preoperative and postoperative session, and they completed the seven two-choice discrimination subtasks in an identical order. Data are expressed as mean and standard deviation. Preoperative and postoperative mean trials to criterion, shift cost, percent correct second choice within trial responses, and latency to dig (seconds) were analyzed using repeated measures ANOVAs with Stage (comprising the seven subtasks) as the within-group repeated measure and Lesion_type (MD lesion or MD sham control) as the between-group factor. For significant interactions, post hoc simple main effects analyses were performed using additional two-way ANOVAs for repeated measures (e.g., ID/ED shift) or independent t tests for relevant subtasks (corrected for multiple comparisons). Univariate ANOVA was used to determine whether the Shift between the two sensory dimensions between the MD lesion or MD sham control had an effect on the number of errors performed in the ED subtask. All statistical analyses were calculated using SPSS 24 software. The significance level was set at p < 0.05.
Results
MD lesion site
Of the 25 adult rats involved in this experiment, 15 sustained MD lesions and 10 were MD sham controls (for details, see Materials and Methods). All 15 MD lesion rats had extensive excitotoxic (NMDA) lesion damage (Fig. 1) within the medial and central subdivisions of the MD, as intended. In all cases the lesion affected between 80% and 90% of the central and medial MD, sparing only the lateral portions. The MD was consistently shrunken, and at the lesion site there was significant cell loss. In all cases, the adjacent anterior thalamic nuclei (ATN) were spared. In two cases, there was unilateral (right-sided) damage in the lateral habenulae (LHb) and some damage to the central medial nucleus lying underneath the MD.

A, Photomicrographs of the thalamus in a MD sham control rat (top) and a MD lesion rat (bottom). B, A series of coronal schematics throughout the MD showing the area of cell loss in the MD lesion group. Numbers refer to the distance from bregma (Paxinos and Watson, 2008). 3V: third ventricle; dLGN: dorsal lateral geniculate nucleus; LHb: lateral habenula; LD: laterodorsal thalamus; MD: mediodorsal thalamus; MHb: medial habenula; PV: paraventricular nucleus; VPM: ventral posterior medial thalamus.
The MD and attentional set-shifting
Preoperatively, rats were pseudo-randomly assigned to either of two groups: MD lesion or MD sham control, based on the number of trials needed to reach the learning criterion (six correct consecutive trials in nine trials) during the SD subtask. Preoperative data from one MD sham control rat had to be discarded. An independent samples t test confirmed there was no significant difference between the two groups during preoperative learning of the SD, t(22) = 0.095, p > 0.05, with rats from both groups requiring a similar number of trials to reach criterion, MD lesion (mean (M) = 9.67, SD = 2.44) and MD sham control (mean = 9.78, SD = 3.31). Overall trials to criterion during the preoperative training session are presented in Figure 2A.
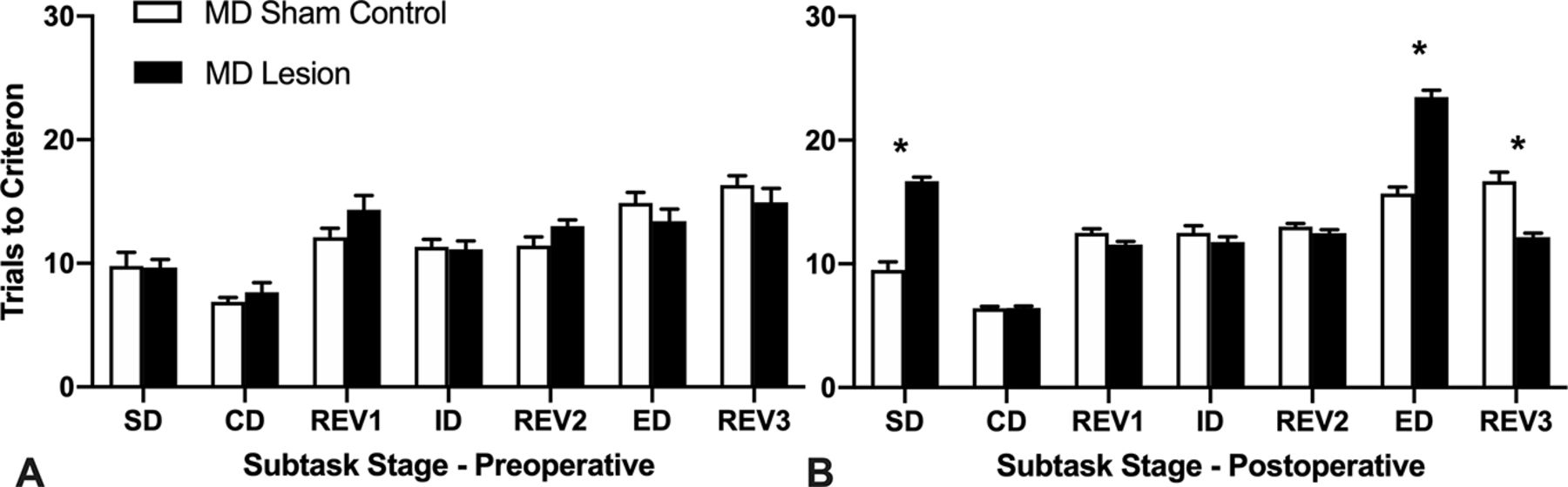
Mean (+SEM) trials to criterion data for the attentional set-shifting task. A, Preoperative test session. B, Postoperative test session. MD lesion rats were significantly slower to learn the SD and the ED subtasks compared with MD sham controls (*p < 0.001). MD sham control rats, like MD lesion rats, needed more trials to learn the ED, as expected. However, the MD lesion rats required significantly fewer trials to criterion to learn REV3, the reversal occurring immediately after the ED, compared with the MD sham controls (*p < 0.001). Also see Extended Data Figure 2-1 showing boxplots of the mean trials to criterion for individual rats during the postoperative session of the attentional set-shifting task.
Extended Data Figure 2-1
Boxplots of the mean trials to criterion for individual rats during the postoperative session of the attentional set-shifting task. MD lesion rats took significantly more trials to learn the SD and the ED subtasks compared to MD sham controls (*p < 0.001). MD sham control rats also needed more trials to learn the ED, as expected. The MD lesion rats required significantly fewer trials to learn REV3, the reversal occurring immediately after the ED, compared to the MD sham controls (*p < 0.001). Box shows 1st and 3rd quartile and whiskers are the minimum and maximum trials to criterion for individual rats. Download Figure 2-1, TIF file.
Postoperatively, a repeated measures ANOVA with Stage (7 subtasks: SD, CD, REV1 ID, REV2, ED, REV3) as the within-group factor and Lesion_type (MD lesion vs MD sham control) as the between-group factor revealed a significant interaction, Stage × Lesion_type, F(6,138) = 63.65, p < 0.001, a significant main effect of Stage, F(6,138) = 184.73, p < 0.001, and a significant main effect of Lesion_type, F(1,23) = 20.22, p < 0.001 (Fig. 2B). To explore the interaction effect, post hoc comparisons of the simple main effects of Stage (corrected for multiple comparisons, p < 0.007; α = 0.05 divided by seven tests) were computed. For the SD subtask, MD lesion rats required more trials to criterion (M = 16.67, SD = 1.23) compared with MD sham controls (M = 9.50, SD = 2.07), indicating that the rats with the MD lesion were slower to acquire the new sensory discrimination. An independent samples t test confirmed this difference was significant, t(23) = 10.88, p < 0.001. However, once the rats with MD lesion acquired the rewarded sensory dimension (to respond either to the digging media, or to the odor), they were unimpaired in subsequent subtasks that maintained the same rewarded sensory dimension, namely, concurrent discrimination (CD): t(23) = 0.00, p =1.0, and ID shift: t(23) = 1.08, p < 0.291. MD lesion rats were also not impaired in the reversals (REV1 and REV2) of the reward contingencies associated with the learned exemplars completed after the CD subtask, REV1: t(23) = 2.31, p < 0.030, or after the ID subtask, REV2: t(23) = 1.34, p < 0.192, although, all rats were familiar with the reversal rule as they had completed the ID/ED task preoperatively (see Materials and Methods). Extended Data Figure 2-1 shows the mean and first and third quartile box plots with the smallest and largest numbers of postoperative trials to criterion (whiskers) for the individual rats during each of the subtasks.
For the ED subtask, the rats had to shift their attention to the previously irrelevant (unrewarded) sensory dimension. Thus, given the rats had acquired the attentional set strategy, as expected, all rats required more trials to criterion to learn the ED subtask. MD sham controls required (M = 15.70, SD = 1.64) more trials to learn the ED when compared with trials required to learn the ID (M = 12.5, SD = 1.84). Similarly, the MD lesion rats required more trials (M= 23.47, SD = 2.07) compared with trials to learn the ID (M = 11.73, SD = 1.67; see Fig. 2B). A repeated measures ANOVA computing the trials to criterion for these two subtasks (ID vs ED) showed a significant Stage × Lesion_type interaction, F(1,23) = 99.96, p < 0.001, a significant effect of Stage, F(1,23) = 306.11, p < 0.001, and a significant effect of Lesion_type, F(1,23) = 32.83, p < 0.001 (Fig. 2B). The interaction occurred as the MD lesion rats required significantly more trials to criterion to consistently implement a new choice strategy to learn which of two stimuli was rewarded from the previously ignored sensory dimension compared with the MD sham controls, t(23) = 10.00, p < 0.001.
Interestingly, for REV3, the reversal subtask that occurred after the ED shift, MD lesion rats required fewer trials to criterion (M = 12.13, SD = 1.25) compared with the MD sham controls (M = 16.7, SD = 2.26; see Fig. 2B). An independent samples t test confirmed this difference was significant, t(23) = 6.51, p < 0.001, suggesting a facilitation in performance as a consequence of over-training experienced during the ED subtask (see Discussion for interpretation).
The additional repeated measures ANOVA of session (preoperative vs postoperative) shift cost (Fig. 3A) between Lesion_type computed as the number of trials to criterion for the ED subtask minus the number of trials to criterion for the ID subtask for each session showed a significant Session × Lesion_type interaction, F(1,22) = 44.06, p < 0.001, a significant effect of Session, F(1,22) = 38.27, p < 0.001, and a significant effect of Lesion_type, F(1,22) = 15.46, p =0.001 (Fig. 3A). The significant interaction occurred as there was a small shift cost for both groups during the preoperative test session. However, during the postoperative test session only the MD lesion rats showed a significant shift cost as a consequence of the task demands changing that required an adjustment to the previously well-established response strategy (p < 0.001). By contrast, the MD sham controls showed a similar shift cost to their preoperative performance.
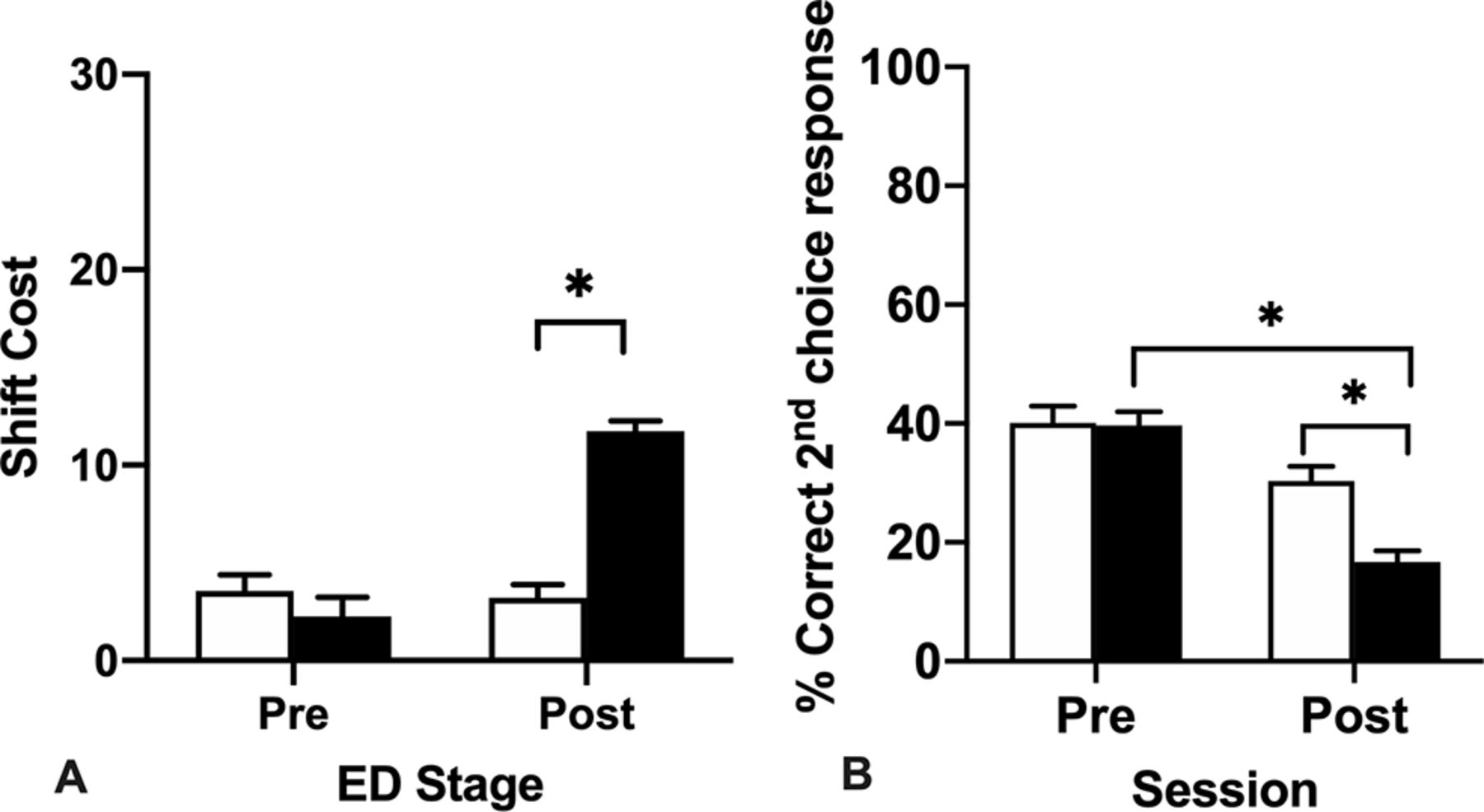
A, Mean (+SEM) trials to criterion shift cost data for the ED stage for preoperative (Pre) and postoperative (Post) testing sessions. The shift cost is computed as the number of trials to criterion in the ED minus the number of trials to criterion in the ID. Both groups of rats showed a shift cost during the ED, when they had to learn to attend to the previously irrelevant dimension. However, the shift cost for the MD lesion rats in the postoperative test session was significantly different to the MD sham controls. B, Mean (+SEM) percent correct second choice within trial responses made during the preoperative and postoperative performance sessions of the attentional set-shifting task. MD lesion rats made fewer correct second choice within trial responses than MD sham controls (*ps = 0.001).
Within trial choice responses as a measure of rapid adaptability after MD lesion
Given the observed deficits in the MD lesion group, we investigated the behavioral responses made in the MD lesion or MD sham control rats. Given the task design and a criterion of six consecutive correct responses before switching to the next stage, overall during testing, the rats do not complete many trials. Nevertheless, we could determine whether the rats rapidly adapted their choice responses in the here and now (within trial choice responses) as measured by the number of correct second choice within trial responses made, i.e., when the rat dug from the correct bowl only after visiting the incorrect bowl first but without digging in it, divided by the total number of correct responses (correct first and second choice responses combined) for each session. During each subtask of the session, we recorded whether the rat made a correct choice, either on the first attempt (“first choice response”), or on the second attempt (“second choice within trial response”). We did not include error trials in this analysis.
A repeated measures ANOVA of total percent correct second choice within trial responses was conducted with session (preoperative vs postoperative) as the repeated measure × Lesion_type revealed a significant interaction of Session × Lesion_type, F(1,21) = 9.40, p = 0.006, a significant main effect of Session, F(1,21) = 57.06, p < 0.001, and a significant main effect of Lesion_type, F(1,21) = 7.18, p = 0.014 (Fig. 3B). To investigate the interaction effect, post hoc comparisons of the simple main effects showed that the MD lesion rats (M = 28.18, SD = 7.61) made fewer correct second choice within trial responses compared with the MD sham controls (M = 35.23, SD = 8.46). This deficit suggests the MD lesion rats had a diminished ability to rapidly bind together their previous choice about what stimuli are worth sampling to update their choice response strategy within the trial, rather than a deficit in perseverative responding, or an inability to respond to negative feedback.
In addition, we analyzed the rats’ errors made reaching criterion during the ED subtask, as a consequence of whether they experienced a sensory dimension shift from odor to digging medium or digging medium to odor, which indicated that the increase in errors occurs regardless of the direction of the attentional shift. The univariate ANOVA with Lesion_type and Sensory Shift as the between-subject factors and errors to criterion for the ED subtask as the dependent measure revealed a main effect of Lesion_type, F(1,21) = 106.03, p < 0.001, but no main effect of Sensory Shift, F(1,21) = 3.84, p = 0.063 and no interaction effect, F(1,21) = 0.19, p = 0.666.
Latency to dig changes
Latency to dig (in seconds) for error trials and correct trials were also computed for each rat in preoperative (Fig. 4A,C) and postoperative (Fig. 4B,D) analyses. Separate repeated measures ANOVAs were used to assess the latency to dig for the preoperative or postoperative sessions for either error trials or correct trials with Stage (seven subtasks: SD, CD, REV1 ID, REV2, ED, REV3) as the within-group factor and Lesion_type (MD lesion vs MD sham control) as the between-group factor.
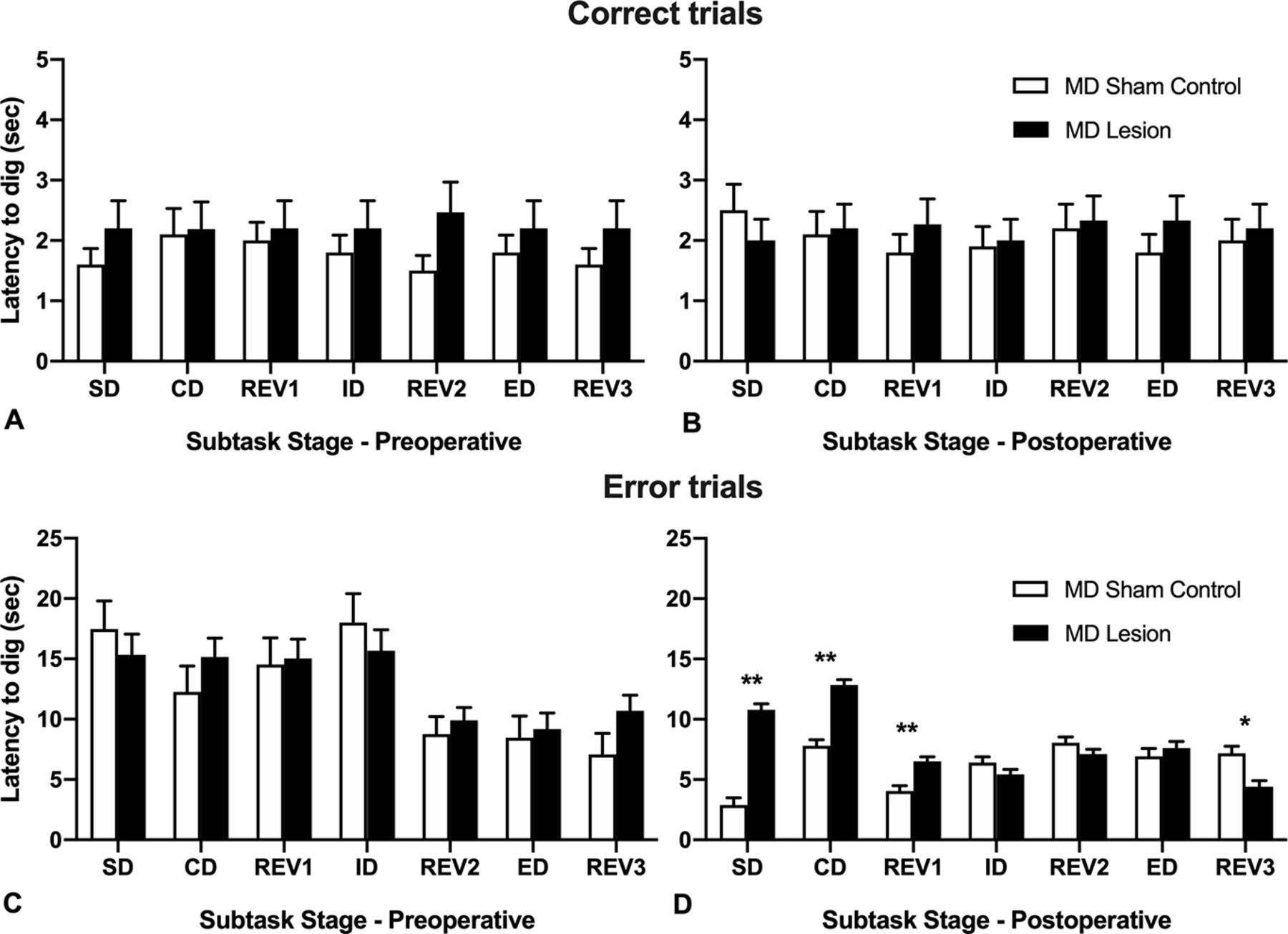
Mean (+SEM) latencies to dig (seconds) for MD sham controls and MD lesion rats during each subtask of the ID/ED attentional set-shifting task. A, Preoperative correct trials. B, Postoperative correct trials. C, Preoperative error trials. D, Postoperative error trials (**p < 0.001, *p = 0.002).
For error trials (Fig. 4C), the repeated measures ANOVA for the preoperative session revealed a significant main effect of Stage, F(1,21) = 23.99, p < 0.001, but no effect of lesion, and no interaction, Fs < 1.0. This result indicates there was no difference in response times between the pseudo-randomly assigned groups (MD lesion and MD sham controls) preoperatively and that all rats responded quicker on errors trials as they moved through subsequent subtasks within the session.
In contrast, the repeated measures ANOVA of latency to dig during postoperative error trials (Fig. 4D) revealed a significant interaction of Stage × Lesion_type, F(6,132) = 26.81, p < 0.001, a significant main effect of Stage, F(6,132) = 22.45, p < 0.001, and a significant main effect of Lesion_type, F(1,22) = 57.15, p < 0.001. To investigate the interaction effect, post hoc comparisons of the simple main effects (corrected for multiple comparisons) were computed. Postoperatively, MD lesion rats responded slower on error trials during the SD subtask, t(22) = 10.09, p < 0.001, the CD subtask, t(22) = 7.65, p < 0.001, and the REV1 subtask, t(22) = 4.44, p < 0.001. However, for the final reversal subtask, REV3, completed after the ED shift, MD lesion rats were quicker on the error trials than MD sham controls, t(22) = 3.53, p = 0.002. This result in combination with the reduced trials to criterion required to complete this subtask (Fig. 2A) suggests that the MD lesion rats may have experienced overtraining because of the number of additional trials to criterion required to adapt their choice responding in the ED subtask (Dhawan et al., 2019). For correct trials, repeated measures ANOVAs for the preoperative or postoperative session showed no significant main effects and no significant interaction (ps > 0.05).
Discussion
This set of experiments investigated the active influence of rat MD in cognitive flexibility. More specifically, we examined whether rats with excitotoxic lesions of the central and medial MD were impaired in performing an attentional set-shifting task that measures ID and ED shifts in attention to sensory discriminations (digging media or odors). Our new results indicate that rats with MD lesions required more trials to criterion to learn the optimal response strategy during the initial SD. Although, once this stable response strategy had been learnt, MD lesion rats were able to attain an attentional set strategy as they acquired further two-choice discriminations involving the same sensory dimension during the CD and ID shifts, and during reversals (REV1 and REV2) of the reward contingencies associated to the exemplars. However, when the relevant sensory dimension was switched, for the ED subtask, the MD lesion rats were markedly impaired and required many more trials to update their choices using a new response strategy, as measured by the significant ED shift cost. Further, for REV3, the third reversal performed after the ED shift, the MD lesion rats showed facilitated learning, suggesting they had received “overtraining” as a consequence of completing many more trials to reach criterion in the ED subtask (see below). The analyses of behavioral responses indicated that the MD lesion rats made significantly fewer correct second choice within-trial responses suggesting that information transfer via the MD is critical for the rapid, within-trial monitoring and updating of an optimal choice response. Interestingly, similar updating deficits in choice responding have been previously reported in rhesus macaques with magnocellular MD excitotoxic lesions (Chakraborty et al., 2016).
Learning and updating
These behavioral data accord somewhat with previous studies conducted in different species. For example, marmosets with lateral PFC lesions (Dias et al., 1996) or rats with medial PFC lesions (equivalent frontal regions; Birrell and Brown, 2000) are also impaired at the ED subtask, showing a similar ED shift cost as our MD lesion rats. However, unlike our MD lesion rats, these marmosets and rats did not need more trials to reach criterion during the SD subtask. Instead, the MD lesion rats’ learning deficit during the SD subtask accords with rats that had ATN lesions, who similarly required more trials to acquire the SD subtask (Wright et al., 2015), although these ATN lesion rats were not impaired during the ED subtask. Instead, and in contrast to our MD lesion rats, rats with ATN lesions continued to be impaired during the CD subtask and subsequent ID shifts (Wright et al., 2015) indicating that they never properly learnt the discrimination rule, which suggests an intact ATN (a brain structure adjacent to the MD but with markedly different cortico-thalamo-cortical connectivity) is important for supporting the formation of an attentional set strategy.
As already indicated, with more trials, our rats with MD lesions were able to learn the SD subtask to criterion and apply the optimal rule during subsequent subtasks. For the SD subtask, trials revert back to presenting only one sensory dimension and two stimulus exemplars. Other studies also confirm MD lesion rats are impaired in acquisition of an initial task rule (Hunt and Aggleton, 1998). Other rodent studies causing temporary perturbations to the MD also show impaired discrimination learning (Ferguson and Gao, 2018; Courtiol et al., 2019). Thus, these results from across species reinforce the notion that the MD is not simply mimicking the PFC (Mitchell et al., 2007a, 2014; Mitchell and Gaffan, 2008; Miller et al., 2017; DeNicola et al., 2020). Consequently the influence of other corticocortical interactions that also indirectly transfer information to the cortex via the MD and other thalamic structures, e.g., those PFC interactions with limbic structures in the temporal lobes and sensory association areas are also critical during learning sensory discriminations (Floresco and Grace, 2003; Mitchell, 2015; Bueno-Junior and Leite, 2018; Chakraborty et al., 2019; Banerjee et al., 2020; Pelekanos et al., 2020).
However, during the ED subtask, our MD lesion rats were markedly impaired at rapidly adapting this now well-established choice response strategy when they had to attend to the previously irrelevant (nonrewarded) sensory dimension. The demands on cognitive flexibility, response inhibition, and adapting behavioral responses after negative feedback are increased when the animals must make an ED shift (Dhawan et al., 2019). The increased effort involved in adapting to these changes in attention is measured by a shift cost, and it is expected that more trials to criterion are required during learning the ED than during learning the ID shift. While the ID/ED shift cost was higher for both groups (Fig. 2B), it was significantly increased for the MD lesion rats compared with the MD sham controls. As indicated, increased shift costs have also been observed in marmosets and rats with lesions to the comparative PFC regions (Dias et al., 1996; Birrell and Brown, 2000). Surprisingly though, rats with damage to the nucleus reuniens (Re), another thalamic structure located near the MD, which interconnects the medial PFC and hippocampus (Hoover and Vertes, 2007, 2011; Vertes et al., 2015) do not cause deficits in the ED subtask or produce a significant ED shift cost. Instead, rats with NRe damage are impaired at acquiring the attentional set strategy, similar to rats with ATN lesions (Linley et al., 2016). Thus, our MD thalamus results are unique.
Evidence from studies in mice suggests the MD supports the frontal cortex to sustain intracortical attentional control without transferring categorical information about a particular task rule (Schmitt et al., 2017), or that medial PFC-MD projections are important for behavioral flexibility but not task engagement (Marton et al., 2018; Nakayama et al., 2018). Further, the MD has been shown to have a critical role in maintaining the balance between excitation and inhibition in dorsomedial PFC via its influence on interneurons (Ferguson and Gao, 2018; Anastasiades et al., 2021) and pyramidal neurons (Delevich et al., 2015; Collins et al., 2018). Our results are supportive of these circuit level interactions, although our deficits indicate the influence of MD on frontal circuits is specific to certain aspects of the attentional set-shifting task, e.g., linked to the ED shift and learning the optimal choice response strategy, but not to the ID shift or completing reversals. However, our study involved permanent MD lesions, which are likely to have caused some adaptations across the network, while the studies in mice likely involve short-term changes as temporary inactivation of the MD was employed. Nevertheless, it may be proposed that after MD perturbations, while the animal is still able to detect a change in the task demands (i.e., they are responsive to negative feedback), they are impaired at rapidly adapting their behavior and coordinating correct choice responses within a trial after a change to the already established choice response strategy (Hunt and Aggleton, 1998; Block et al., 2007; Chakraborty et al., 2016). Interestingly though, in both these rodent studies (Hunt and Aggleton, 1998; Block et al., 2007), the learning deficits were attributed to increased perseverative responding. Others have also observed increased perseverative responding after MD perturbations (Ferguson and Gao, 2018). In contrast, our current rodent work and previous work in rhesus macaques has shown an intact MDmc is necessary for rapid reward guided learning of complex discriminations (Mitchell et al., 2007a; Chakraborty et al. 2016, 2019), although the deficits linked to learning in monkeys are attributed to increased response switching (Mitchell et al., 2007a; Chakraborty et al., 2016). These congruent results across species after MD damage suggest that cortical information transfer via the MD is particularly important when rapid, within a trial changes in choice response strategies linked to establishing a new rule are required.
Reversal learning
For the reversal subtasks (REV1 and REV2), somewhat surprisingly, our MD lesion rats were unimpaired during reversals in the reward contingencies. Rats and marmosets with comparable medial PFC damage are also not impaired in reversal learning subtasks (Dias et al., 1996; Birrell and Brown, 2000). Instead, OFC perturbations cause deficits in the reversal subtasks, while leaving performance intact during the SD and ED subtasks of the ID/ED task (Dias et al., 1997; McAlonan and Brown, 2003; Chase et al., 2012). These observations accord with a large body of research indicating that separate mammalian PFC subregions differentially contribute to learning, memory, and other cognitive functions (Clark et al., 2004; Rudebeck et al., 2008; for review, see Uylings et al., 2003; Dalley et al., 2004; Izquierdo et al., 2017).
MD is reciprocally connected to the OFC and medial PFC (Groenewegen, 1988; Ray and Price, 1992; Preuss, 1995). Thus, this double dissociation in deficits between these reciprocally interconnected MD-OFC and MD-medial PFC brain regions needs to be reconciled. First, it must be noted that the MD rats had experienced all of the subtasks (and thus were familiar with the concept of reversal learning and that it can appear in the task structure) during their preoperative test session, which may have reduced or eliminated the reversal learning deficit (Costa et al., 2015; Jang et al., 2015). Further, for the MD, while previous evidence of reversal learning deficits are reported for some tasks after MD perturbations (Hunt and Aggleton, 1998; Chudasama et al., 2001; Block et al., 2007; Ostlund and Balleine, 2008; Parnaudeau et al., 2013, 2015; Ferguson and Gao, 2018), the evidence is mixed (Beracochea et al., 1989; Alcaraz et al., 2018; Fresno et al., 2019). These mixed effects may be the consequence of differences in task structure and the way that the reversal is introduced. Other factors, including the extent of disruption caused to thalamic structures adjacent to the MD may also be a factor (Mitchell and Chakraborty, 2013; Wolff and Vann, 2019). Indeed, one recent study has identified that another thalamic structure nearby to the MD, the submedius thalamus, may contribute a role in reversal learning, instead of the MD (Fresno et al., 2019). Consequently, our current results and this above evidence highlights that the information transfer between OFC and MD is not always critical for reversal learning per se Taken together, it is clear that the MD is not simply mimicking the behavioral deficits observed after PFC damage, and instead shows that the MD and different subregions of PFC are supporting different aspects of cognitive flexibility. At least in monkeys, OFC-MDmc interactions are critical for adaptive, value based decision-making (Mitchell et al., 2007b; Izquierdo and Murray, 2010; Browning et al., 2015), while in rodent studies, the OFC-striatal part of the cortico-striatal-thalamic neural circuits have been implicating in reversal learning (Yin et al., 2005; Gremel and Costa, 2013; Izquierdo et al., 2017; Nakayama et al., 2018; Yang et al., 2021) or OFC-submedius thalamus circuits (Fresno et al., 2019). Other OFC-striatal-thalamic and OFC-thalamic interactions are potentially more involved in supporting reversal learning. In addition, thalamic inputs from the intralaminar thalamic nuclei and motor thalamus to the striatum contribute a selective role in inhibitory control and behavioral flexibility (Saund et al., 2017).
Intriguingly, our additional analyses used to investigate the types of behavioral deficits occurring after MD lesions showed that our rats made fewer correct second choice within-trial responses. Given that our MD lesion rats required more trials to criterion to learn the SD and ED subtasks, this change in behavioral responses suggests that without the MD thalamus, our rats could not rapidly adapt their choice responses as the trial was progressing (within-trial) after the changed task demands. However, this reduced correct second choice within-trial responding does not suggest the rats adopted perseverative responding (they would have potentially been impaired in reversal learning if they had) or were not learning about the stimulus and associated rewards. We can conclude this because, after acquiring the ED shift, the MD lesion rats required fewer trials to criterion to learn the final reversal subtask, REV3. This facilitation of learning during the final reversal subtask suggests that the MD lesion rats had experienced “overtraining” on all of the stimulus features associated with each sensory dimension as they required more trials to learn the ED subtask (Dhawan et al., 2019). It is well established that overtraining rats during discrimination learning can eliminate any reversal learning deficits (for review, see Reid, 1953; Lovejoy, 1966; Nilsson et al., 2015). As Pearce and Mackintosh (2010) indicate, until learning is fully consolidated, all stimulus features continue to be attended to and this seemed to be so for our rats, even with the MD lesion. Thus, we propose, in accord with others, that through experiencing these extra trials during the ED subtask, the MD lesion group may have developed a greater understanding of the rewarded and unrewarded stimulus dimensions, thus increasing the salience of the predictive cues and reducing/eliminating the number of factors that can led to an error. In essence, these extra trials in the ED subtask helped our MD lesion rats to slowly learn the optimal response strategy so when the final reversal was encountered, they were readily able to continue to implement this optimal response strategy for the sensory dimension while simply reversing the two stimulus exemplars (something they were already well-practiced at doing). An MD lesion does not render an animal or human densely amnesic or cause them to be insensitive to the receipt of rewards (Mitchell, 2015). Instead, we are proposing that after the loss of the MD, an animal or human is unable to rapidly learn a new choice response strategy or rapidly update a well-established optimal one.
However, in the Birrell and Brown (2000) study, rats with medial PFC lesions did not show a facilitation of learning during the final reversal subtask after showing a similar ED shift cost to our MD lesion rats. We cannot be certain why medial PFC lesion rats also did not show a facilitation of learning in the final reversal subtask, although exploring whether they experience overtraining is worthy of further investigation. What is known is that the medial PFC is proposed to contribute to learning via supporting the processing of error feedback related to adapting stimulus-action selection. For example, during instrumental learning, medial PFC lesions in rats contribute to the acquisition but not the expression of goal-directed actions (Ostlund and Balleine, 2005).
There may be other explanations for the SD and ED impairments observed in our MD lesion rats. With the ID/ED task, it is important to establish that the animals understand the relevant sensory dimension that is rewarded across the related subtasks within the session. Fortunately, in our animals, this transfer of knowledge was evident as the MD lesion rats showed reliable learning in the subtasks, CD, REV1, ID, and REV2, that required the implementation of choice responses (follow the same rule) to the same sensory dimension as the SD to receive reward. Further, both sham controls and MD lesion rats showed similar numbers of trials to criterion during the REV1 and REV2 subtask, which indicates that they did not favor one feature of the stimulus dimension more than the other or that they were insensitive to negative feedback. However, we only used male rats in this current study so the results might not transfer to female rats. Additionally, the MD is a subcortical node in the olfactory neural circuitry that also includes the OFC (Courtiol and Wilson, 2015; Veldhuizen et al., 2020). However, evidence collected from rodents with MD perturbations or humans with strokes affecting the MD indicates that changes to the MD do not impair olfactory discriminations (Tham et al., 2011; Courtiol et al., 2019). Moreover, our MD lesion rats showed similar levels of olfactory discriminations as the MD sham controls (for further details, see Materials and Methods).
Finally, the diffuse influence of the MD thalamocortical inputs to several frontal cortex structures (Mitchell, 2015; Alcaraz et al., 2016; Kuramoto et al., 2017; Wolff and Vann, 2019) supports previous findings implicating medial PFC-MD connectivity in flexible behaviors (Marton et al., 2018; Nakayama et al., 2018). Furthermore, the MD along with the OFC provides additional evidence for the contribution of these areas in supporting online, “here and now” reward-guided learning and decision-making (Wallis, 2007; Schoenbaum et al., 2011; Gardner et al., 2017, 2019). Intact OFC connectivity is essential to the animals’ ability to perform reward-guided learning in order that other interconnected neural networks, likely including corticocortical, cortico-striatal, and cortico-thalamo-cortical connectivity, can determine the optimal choice responses and implement the appropriate actions (Rushworth et al., 2011; Browning et al., 2015; Chakraborty et al., 2016; Nakayama et al., 2018; Perry et al., 2021).
The ID/ED task is analogous to the Wisconsin Card Sorting Test (WCST) in humans. In healthy humans performing the WCST during neuroimaging, the MD is activated during responding to negative feedback after the choice has been executed (Monchi et al., 2001). Unfortunately, thus far, humans with stroke damage in the MD have cognitive deficits that are clinically very poorly defined (Pergola et al., 2018). However, people diagnosed with Alzheimer’s disease (AD), PD, or schizophrenia show impaired attentional set-shifting performance (Owen et al., 1993; Monchi et al., 2004; Barch et al., 2009). Neuroimaging and postmortem studies show marked changes in the MD and/or ATN in these diseases (Hornberger et al., 2012; Pergola et al., 2015; Ouhaz et al., 2018; Perry et al., 2019). Our current evidence advocates for studies investigating cortico-thalamocortical transfer of information in people diagnosed with AD combined with more frontal pathology, or PD, or schizophrenia.
To conclude, excitotoxic damage to the rodent central and medial MD selectively increased the trials to criterion on the ED subtask, producing a shift cost. This selective performance deficit is similar to monkeys with lateral PFC inactivation (Dias et al., 1996, 1997) and rats with medial PFC lesions (Birrell and Brown, 2000). Further, this deficit after MD lesions contrasts with monkeys or rats with perturbations to the OFC, who are selectively impaired on reversal learning, but not on ED shifts (Dias et al., 1997; McAlonan and Brown, 2003; Chase et al., 2012). As evidenced, the frontal cortex is critically involved in value-based decision-making and reward-guided learning (Miller, 2000; Wallis, 2007; Rushworth et al., 2011). However, cortico-thalamocortical connections also contribute a role (Izquierdo and Murray, 2010; Mitchell et al., 2014; Browning et al., 2015; Chakraborty et al., 2016; Pelekanos et al., 2020). In rodents, the MD and the medial PFC together appear crucial for binding reward information and behavior (Corbit et al., 2003; Bradfield et al., 2013), as inhibition of dorsomedial PFC-projecting MD neurons results in rats having difficulties with tracking changes in action-outcome contingencies (Alcaraz et al., 2018). In the current study, we show that rat medial and central MD are critical for rapidly updating an optimal choice response strategy. That is, when the MD is damaged, there is a diminished ability to rapidly learn (within a trial) a choice response strategy, as well as update a well-established one as task demands change. Behaviorally, our MD lesion rats made less correct second choice within-trial responses, suggesting they could not rapidly alter their choice response strategy when they encountered the unrewarded stimuli on a given trial.
Acknowledgments
Acknowledgements: We thank Professor Verity Brown and Dr. David Tait, who taught Z.O. to perform the ID/ED attentional set-shifting task during an extended visit to their research lab at St Andrews University.
Footnotes
The authors declare no competing financial interests.
Z.O. was supported by the Royal Society Newton International Fellowship NF160862. A.S.M. was supported by the Wellcome Trust Grant WT110157/Z/15/Z.
This is an open-access article distributed under the terms of the Creative Commons Attribution 4.0 International license, which permits unrestricted use, distribution and reproduction in any medium provided that the original work is properly attributed.
References
In this issue

{kind=link}
{kind=link}
{kind=link}
{kind=link}



