

Article Figures & Data
Figures
- Figure 1.
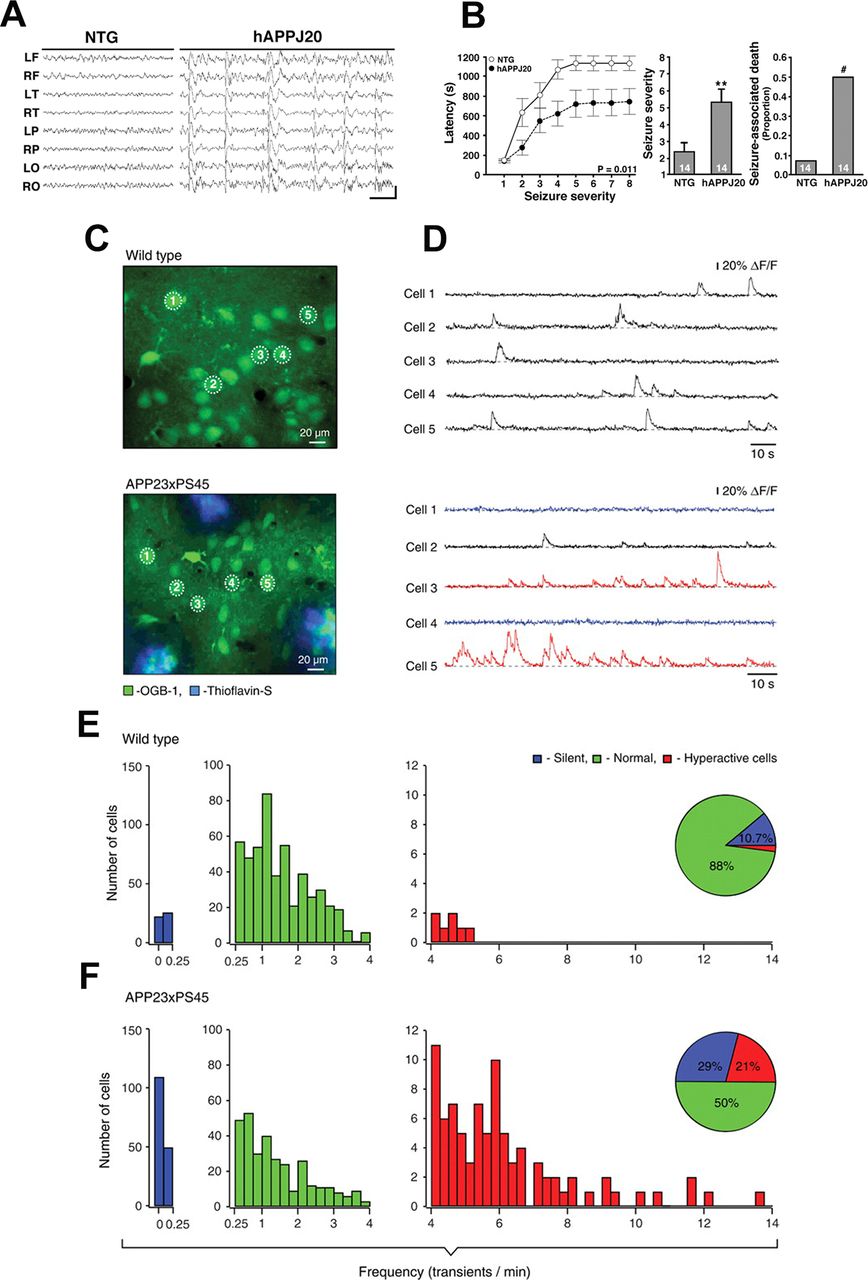
Neuronal network hyperexcitability at advanced stages of pathology in hAPP/Aβ mouse models of AD. A, B, Aberrant synchronous neuronal network activity, spontaneous nonconvulsive seizures, and increased susceptibility to PTZ-induced seizures in four- to seven-month-old hAPP-J20 mice. Reproduced from Palop et al. (2007) with permission from Elsevier. A, Chronic cortical EEG recordings performed in freely moving, untreated hAPP-J20 mice, and non-transgenic (NTG) controls. L, left; R, right; F, frontal; T, temporal; P, parietal; O, posterior-parietal, indicate the position of recording electrodes. In contrast to NTG mice, which showed normal EEG activity (left), hAPP-J20 mice exhibited frequent (5–50/min) generalized cortical epileptiform (interictal) spike discharges (right). Calibration: 1 s and 400 mV. B, Mice were injected intraperitoneally with PTZ (GABAA antagonist), behavior was videorecorded, and seizure severity was scored off-line. Compared with NTG controls, hAPP-J20 mice had shorter latencies to reach a given seizure severity (left), greater overall seizure severity (center), and more seizure-associated deaths (right); ∗∗p < 0.01 versus NTG by Student’s t test; #p < 0.05 by Fisher’s exact test. Quantitative data represent mean ± SEM. C–F, Clusters of hyperactive neurons near amyloid plaques in APP23xPS45 mice. In vivo two-photon calcium imaging from layer 2/3 cortical neurons. Reproduced with permission from Busche et al. (2008). C, D, Spontaneous Ca2+ transients (D) recorded in vivo in the corresponding neurons of the frontal cortex shown in C in a WT (top) and a APP23xPS45 (bottom) mouse. Traces in D, bottom, are color-coded to mark neurons that were either inactive during the recording period (blue) or showed an increased frequency of Ca2+ transients (red). E, F, Histograms showing the frequency distribution of Ca2+ transients in WT and APP23xPS45 mice (in both cases n = 564 cells). There is a substantial increase in the amount of silent and hyperactive neurons in APP23xPS45 mice. (Insets) Pie charts showing the relative proportion of silent, normal, and hyperactive neurons in WT (n = 10) and APP23xPS45 (n = 20) mice.
- Figure 2.
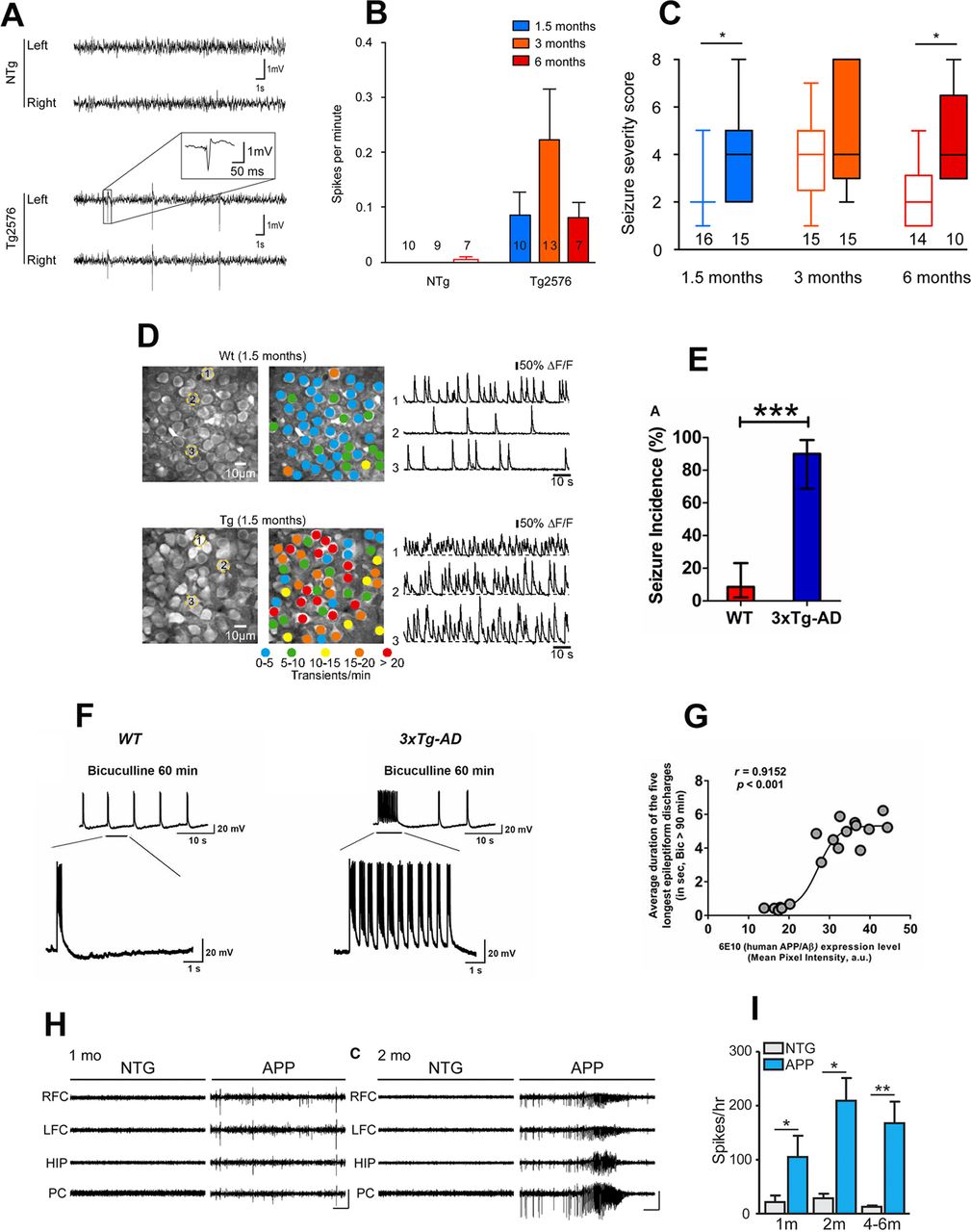
Early-onset neuronal network hyperexcitability in hAPP/Aβ mouse models of AD. A–C, Tg2576 mice exhibit spontaneous epileptiform activity and high susceptibility to pharmacologically induced seizures as young as 1.5 months of age. Reproduced with permission from Bezzina et al. (2015). A, Representative EEG traces from non-transgenic (NTg; top) and Tg2576 (bottom) mice from left and right parietal cortices. Note that only transgenic animals displayed sharp, high-voltage spikes that characterize epileptiform activity (inset). B, Quantitative analysis of the frequency of interictal spikes (mean ± SEM). Two-way ANOVA shows a significant genotype effect (p = 0.013) but no age effect (p = 0.4091) and no interaction (p = 0.3865). Numbers over the horizontal axis indicate the number of mice used in each experimental group. C, Seizure severity score of 1.5-, 3-, and 6-month-old Tg2576 male mice and NTg age-matched littermates. Whiskers boxes represent the interquartile distribution. Number of mice in each group is indicated below the boxes. Tg2576 mice exhibit more severe seizures than NTg at 1.5 and 6 months of age (Dunn’s tests: p < 0.05 for Tg2576 vs NTg at 1.5 and 6 months old). Note that only transgenic animals exhibit lethal seizures. Numbers over the horizontal axis indicate the number of mice used in each experimental group. D, Early hyperactivity of hippocampal neurons of 1.5-month-old APP23xPS45 mice (an age when no plaques are detectable). Reproduced with permission from Busche et al. (2012). Left, CA1 neurons imaged in vivo in a WT and a transgenic mouse, respectively. Center, Activity maps in which hue is determined by the frequency of spontaneous Ca2+ transients, overlaid with the anatomic image (left). Right, Spontaneous Ca2+ transients of the corresponding neurons marked (left). E–G, Early-onset seizure susceptibility and epileptiform activities in three-week-old 3xTg-AD mice (much before plaques and overt cognitive impairment). Reproduced with permission from Kazim et al. (2017). E, Incidence of convulsive seizures after audiogenic stimulation was markedly higher in three-week-old 3xTg-AD mice (blue bar) compared with WT mice (red bar). The data are presented as percent incidence with 95% confidence interval and compared using exact logistic regression stratified by litter; ***p < 0.001, compared with WT. WT (n = 35) and 3xTg-AD (n = 20) mice. F, Ictal-like epileptiform discharges in CA3 pyramidal cells of hippocampal slices from three-week-old 3xTg-AD mice. Left, CA3 intracellular recording from a WT slice after bicuculline addition (50 μm). Within 20 min, bicuculline induced rhythmic, short epileptiform discharges (≤1.5 s in duration) that were ongoing for at least 1 h of continuous recording. Membrane potential at the beginning of recording: −60 mV. Right, CA3 intracellular recording from a 3xTg-AD slice after bicuculline. Bicuculline first induced short synchronized epileptiform discharges that were similar to those in WT slices. However, continuous perfusion with bicuculline induced prolonged epileptiform (ictal-like) discharges (>1.5 s) in 3xTg-AD slice. Membrane potential at the beginning of recording: −65 mV. G, Positive correlation of intraneuronal human APP/Aβ expression in CA3 neurons and ictal-like activity in CA3 region. Correlation analyses revealed a positive relationship between intraneuronal human APP/Aβ immunoreactivity in the CA3 neurons (analyzed by 6E10, human APP/Aβ) and average duration of the five longest epileptiform discharges recorded during a 5-min period after 90 min of bicuculline application in the CA3 region of hippocampal slices from the same mice. Data from Saline-3xTg-AD (n = 9) and 6E10–3xTg-AD (n = 9) was pooled together to evaluate the correlation. The sigmoidal curve based on nonlinear regression is also shown. H, I, Early-onset epileptic activity in one- and two-month-old hAPP-J20 mice. Reproduced with permission from Fu et al. (2019). H, Representative EEG traces from NTg and hAPP-J20 mice at one and two months of age, with epileptiform spikes at one month of age and a seizure at two months of age in hAPP-J20 mice. Electrodes were in left and right frontal cortices (LFC and RFC), hippocampus (HIP), and parietal cortex (PC). Scale bars: 1 mV, 10 s. I, The number of epileptic spikes per hour in NTg or hAPP-J20 mice at one, two, and four to six months of age (n = 3−5 mice per genotype and age).
- Figure 3.
Neuronal network hyperexcitability in tau mouse models. A–G, Hyperexcitability and epileptic seizures in a mouse model of a tauopathy (FTDP-17). Reproduced from García-Cabrero et al. (2013) with permission from Elsevier. A, Intracranial recording of background activity (6–7 Hz) in control mice. B, Spindle-shaped polyspike discharge at 8–10 Hz, during an initial tonic phase (thin arrow indicates the beginning) and a short clonic phase (open arrow signals the beginning) in a FTDP-17 mouse. C, Spontaneous interictal epileptic activity in FTDP-17 mice corresponding to (c1) single spike, (c2) polyspike, (c3) slow wave, and (c4, c5) polyspike-wave discharges. D, Nonconvulsive spontaneous seizure with EEG correlates corresponding to rhythmic, spindle-shaped discharges. E, Spontaneous generalized tonic–clonic seizure in a FTDP-17 mouse manifested in the EEG record as generalized low-frequency (3–6 Hz) poyspike-wave discharge, 36 s in length. Figure shows the records from monopolar electrodes placed over the left frontal cortex with the reference electrodes implanted posterior to λ. F, G, Analysis of seizure latency and length of PTZ-induced generalized seizures in FTDP-17 mice. Mice at three different age spans (1–5, 6–14, and 15–22 months) were injected with a convulsive dose of PTZ (50 mg/kg). F, The time interval between drug administration and development of generalized tonic–clonic seizures and (G) the seizure length were measured. Data are presented as mean ± SEM. Student’s t test was performed for statistical evaluation; *p < 0.05, **p < 0.01, ***p < 0.001 (n = 15–24). H, I, Increased spontaneous synaptic activity in whole-cell patch clamp recordings of layer 3 frontal cortex pyramidal neurons of rTg4510 (hTau P301L) mice. Reproduced with permission from Crimins et al. (2011). Increased frequency of sEPSCs in TG (rTg4510) cells. H, Representative sEPSCs from non-transgenic (NT) and TG cells. I, Bar graphs of mean frequency sEPSCs in NT and TG cells. J, Increased PTZ-induced seizure susceptibility in three-month-old Tau58/4 (htau P301S) mice. Young HET (Tau58/4) mice had higher mean severity scores than WT littermates. Reproduced with permission from Van Erum et al. (2020).
- Figure 4.
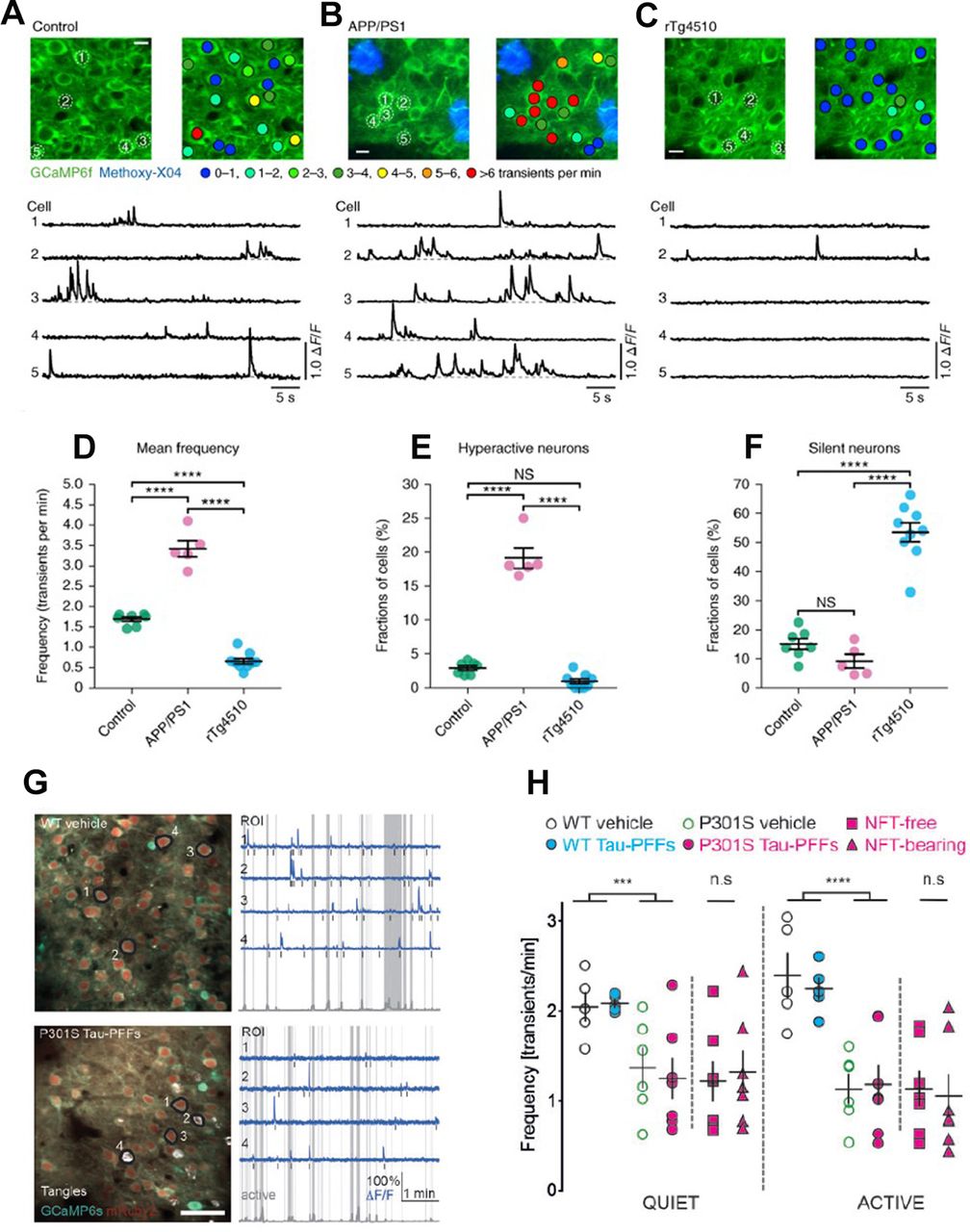
In vivo evidence of suppression of neuronal activity in tau mouse models. A–F, Neuronal silencing in rTg4510 mice as compared with neuronal hyperactivity in APP/PS1 mice. Reproduced with permission from Busche et al. (2019). A–C, top, In vivo two-photon fluorescence images of GCaMP6f-expressing (green) layer 2/3 neurons in the parietal cortex and corresponding activity maps from WT controls (A), APP/PS1 (B), and rTg4510 (C) mice. In APP/PS1 mice, plaques were labeled with methoxy-X04 (blue); in the activity maps, neurons were color-coded as a function of their mean Ca2+ transient activity. Scale bars: 10 μm. Bottom, spontaneous Ca2+ transients of neurons indicated in the top panel. D, Mean neuronal frequencies for controls (1.69 ± 0.05 transients per minute), APP/PS1 (3.42 ± 0.20 transients per minute), and rTg4510 (0.66 ± 0.07 transients per minute); F(2,18) = 171.2, p = 1.93 × 10−12. All post hoc multiple comparisons between genotypes were highly significant: p = 5.42 × 10−9 for controls versus APP/PS1, p = 1.38 × 10−6 for controls versus rTg4510, and p = 1.01 × 10−12 for APP/PS1 versus rTg4510. E, Fractions of hyperactive neurons. Controls: 2.91 ± 0.35%, APP/PS1: 19.11 ± 1.50%, rTg4510: 0.93 ± 0.35%; F(2,18) = 176.2, p = 1.51 × 10−12. Post hoc multiple comparisons were p = 2.84 × 10−11 for controls versus APP/PS1, p = 1.64 × 10−12 for APP/PS1 versus rTg4510 and not significant, p = 0.1045, for controls versus rTg4510. F, Fractions of silent neurons. Controls: 15.05 ± 1.87%, APP/PS1: 9.20 ± 2.36%, rTg4510: 53.48 ± 3.24%; F(2,18) = 77.18, p = 1.48 × 10−9. Post hoc multiple comparisons were p = 2.02 × 10−8 for controls versus rTg4510 and p = 1.08 × 10−8 for APP/PS1 versus rTg4510 and not significant, p = 0.3972, for controls versus APP/PS1. Each solid circle represents an individual animal (controls, n = 7; APP/PS1, n = 5; rTg4510, n = 9), and all error bars reflect the mean ± SEM; the differences between genotypes were assessed by one-way ANOVA followed by Tukey’s multiple comparisons test, ****p < 0.0001. NS, not significant. G, H, Neuronal activity is reduced in P301S mice independently of presence of NFTs. Reproduced with permission from Marinković et al. (2019). G, left, Representative in vivo recordings from WT vehicle and P301S tau-PFFs (tau preformed fibrils-injected) mice. AAV1 transduced neurons are labeled with mRuby2 (red) and GCaMP6s (green). NFTs are labeled with FSB (white). Images are made by averaging 450 time-series frames acquired in vivo at 410 Hz with two-photon lasers tuned to 940 nm for CGaMP6s/mRuby2 and to 750 nm for FSB. Scale bar: 50 mm. Right, Traces (blue) extracted from annotated regions of interest (black) during quiet and active (gray shade) behavioral states classified based on changes in whisking movement (gray trace in the bottom). Note that traces 2 and 4 in P301S tau-PFFs group are from NFT-bearing neurons. Black bars mark detected calcium transients. H, Mean frequency of calcium transients during quiet and active states of all neurons detectable in three or more time points. WT vehicle (black), WT tau-PFFs (cyan), P301S vehicle (green), all P301S tau-PFFs: all neurons are denoted as magenta circles, with NFT-free as magenta squares and NFT-bearing as magenta triangles. Data points represent individual mice, n = 5–7 mice per group; black lines represent mean value ± SEM; ***p < 0.001, ****p < 0.0001, WT versus P301S (two-way ANOVA, genotype factor, not significant; Student’s t test).
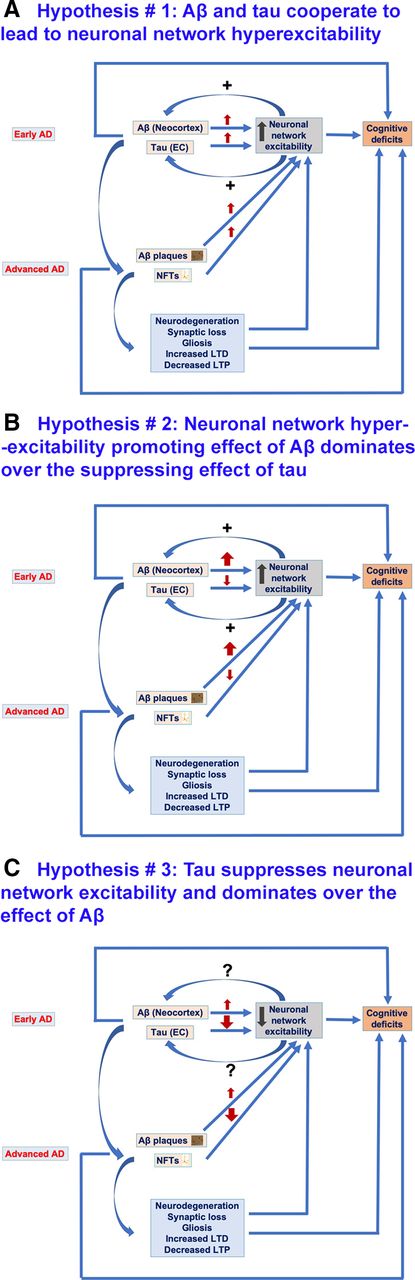
- Figure 5.
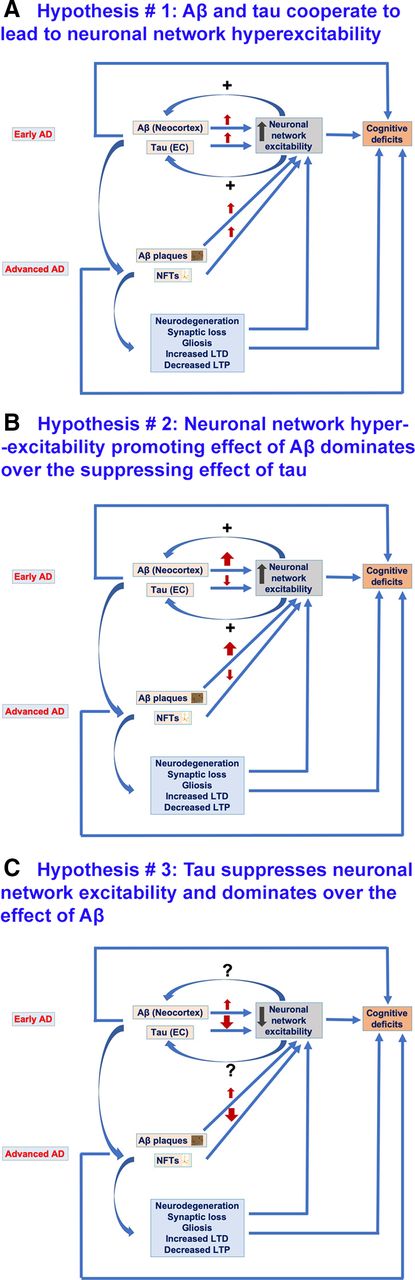
Hypotheses regarding the possible effects of Aβ and tau on neuronal network excitability in AD. A, Hypothesis #1, Aβ and tau cooperate to lead to neuronal network hyperexcitability in AD. At early stages of AD, Aβ is more abundant in the neocortex whereas tau is localized to EC. Both Aβ and tau at early AD stages promote neuronal network hyperexcitability which not only contributes to cognitive impairments but also reciprocally increases Aβ deposition and tau release and spread to other cortical areas across connected neuroanatomical circuitry. Also, at advanced AD stages, both Aβ and tau promote neuronal network hyperexcitability, thus leading to cognitive deficit. Furthermore, Aβ-induced and tau-induced neuronal and synaptic loss, gliosis, and impaired synaptic plasticity (decreased LTP and increased LTD) contribute to neuronal network hyperexcitability and to cognitive deficits, effects also at play in scenarios illustrated in B, C. B, Hypothesis #2, Aβ enhances neuronal network hyperexcitability whereas tau suppresses excitability; the overall phenotype is hyperexcitability as Aβ effect dominates over tau effect. Aβ at early AD stages promotes neuronal network hyperexcitability which not only contributes to cognitive impairments but also increases Aβ deposition and tau release and spread to other cortical areas across connected neuroanatomical circuitry. However, tau at early AD stages suppresses neuronal activity, thus leading to silencing of neuronal networks which could also contribute to AD-related network dysfunction and cognitive deficit. Also, at advanced AD stages, Aβ enhances and tau suppresses neuronal network excitability, both leading to cognitive deficits. This could also be the case in AD patients with higher Aβ deposits than NFTs in their brains. C, Hypothesis #3, tau suppresses neuronal network excitability, whereas Aβ enhances it; the overall phenotype is suppressed excitability as tau suppressive effect dominates over Aβ enhancing effect. Tau both at early and advanced AD stages suppresses neuronal excitability thus leading to silencing of neuronal networks contributing to AD cognitive deficits. Contrarily, Aβ both at early and advanced AD stages promotes neuronal network hyperexcitability however this is dominated by tau suppressive effect. However, this hypothesis cannot explain the tau spread from EC to other cortical areas as increased neuronal activity has been identified to promote propagation of tau. Nonetheless, there could be other mediators of tau spread besides neuronal network hyperexcitability.


Tables
Author(s) and publication year Mouse model/transgene(s)/promoter Age/stage of pathology Neuronal network excitability status Experimental paradigm/neuronal network excitability observation(s) Studies assessing neuronal network hyperexcitability in hAPP/Aβ mice at advanced stages of Aβ plaque pathology and cognitive impairment Palop et al. (2007) hAPP-J20
(hAPP Swedish and Indiana)
PDGF-β promoter4–7 months, Aβ plaques deposition, cognitive impairment, synaptic deficit Increased Experimental paradigm: in vivo chronic video EEG recordings; PTZ-induced seizure susceptibility; in vitro mIPSCs and fEPSPs recordings.
Findings: frequent epileptiform activity including spikes and SWDs and increased PTZ-induced seizure susceptibility in hAPP-J20 mice. Reduced LTP and PPF in hippocampal perforant pathway in hAPP-J20 mice slices. Increased dentate granule cells mIPSCs frequency in hAPP-J20 mice. Remodeling of inhibitory circuits and altered NPY expression in dentate gyrus of hAPP-J20 mice.Verret et al. (2012) hAPP-J20
(hAPP Swedish and Indiana)
PDGF-β promoter4–7 months, Aβ plaques deposition, cognitive impairment, synaptic deficit Increased Experimental paradigm: in vivo chronic video EEG recordings.
Findings: spontaneous epileptiform discharges observed during reduced γ oscillatory activity (generated by inhibitory PV cells) in hAPP-J20 mice. Decreased levels of the interneuron-specific and PV cell-predominant voltage-gated sodium channel subunit Nav1.1. Restoring Nav1.1 level in hAPP-J20 mice increased inhibitory synaptic activity and γ oscillations and reduced hyperexcitability and cognitive deficits.Sanchez et al. (2012) hAPP-J20
(hAPP Swedish and Indiana)
PDGF-β promoter4–6 months, Aβ plaques deposition, cognitive impairment, synaptic deficit Increased Experimental paradigm: in vivo chronic video EEG recordings; fEPSPs in acute hippocampal slices.
Findings: spontaneous epileptiform activity in hAPP-J20 mice. Chronic treatment with levetiracetam reversed abnormal spiking activity, hippocampal remodeling, behavioral abnormalities, synaptic dysfunction, and deficits in learning and memory in hAPP-J20 mice.Martinez-Losa et al. (2018) hAPP-J20
(hAPP Swedish and Indiana)
PDGF-β promoter7–8 months, Aβ plaques deposition, cognitive impairment, synaptic deficit Increased Experimental paradigm: in vivo EEG recordings in freely moving mice.
Findings: epileptiform spikes on cortical EEG in hAPP-J20 mice. Nav1.1-overexpressing, interneuron transplants enhanced reduced network hypersynchrony and improved cognitive functions in hAPP-J20 mice.Minkeviciene et al. (2009) APdE9
(hAPP Swedish and PSEN1: deltaE9) Prp promoter3 and 4.5 months, substantial number Aβ plaques observed in cortex, hippocampus, and amygdala Increased Experimental paradigm: in vivo video EEG recordings; patch clamp electrophysiology; extracellular field recordings in brain slices.
Findings: unprovoked seizures in APdE9 mice. Hyperexcitability in neocortical layer 2/3 pyramidal cells in APdE9 mice on patch clamp recordings. Aβ protofibrils induced neuronal network hyperexcitability in acute brain slices.Ziyatdinova et al. (2011) APdE9
(hAPP Swedish and PSEN1: deltaE9) Prp promoter4–5 months, Aβ plaques in the neocortex and hippocampus Increased Experimental paradigm: in vivo video EEG recordings.
Findings: spontaneous electrographic epileptiform discharges. Antiepileptic drugs that block sodium chan-nels, including carbamazepine, phenytoin, and valproic acid suppressed epileptiform activity in APdE9 mice with increased amyloid pathology.Gurevicius et al. (2013) APdE9
(hAPP Swedish and PSEN1: deltaE9) Prp promoter4 months; Aβ plaques in the neocortex and hippocampus Increased Experimental paradigm: in vivo EEG recordings from the hippocampus, cerebral cortex, and thalamus during movement, quiet waking, non-rapid eye movement sleep, and REM sleep.
Findings: cortical EEG power was higher in APdE9 mice than in WT mice over a broad frequency range (5–100 Hz) and during all 4 behavioral states. Thalamic EEG power was also increased but in a narrower range (10–80 Hz). While power and θ–γ modulation were preserved in the APdE9 hippocampus, REM sleep-related phase shift of θ–γ modulation was altered.Ziyatdinova et al. (2015) APdE9
(hAPP Swedish and PSEN1: deltaE9) Prp promoter4–5 months, Aβ plaques in the neocortex and hippocampus Increased Experimental paradigm: in vivo video EEG recordings.
Findings: spontaneous epileptiform discharges. Antiepileptic drug valproic acid reduced the amount of epileptiform activity, but the effect disappeared after treatment discontinuation.Nygaard et al. (2015) APdE9
(hAPP Swedish and PSEN1: deltaE9) Prp promoter10 months, Aβ plaques in the cortex and hippocampus Increased Experimental paradigm: in vivo video EEG recordings.
Findings: epileptiform activity in the form of SWDs in APdE9 mice. SWDs correlated with spatial memory impairment in these mice. Brivaracetam (a chemical analog of levetiracetam) reduced SWDs and reversed memory impairments in in APdE9 mice.Reyes-Marin and Nuñez (2017) APdE9
(hAPP Swedish and PSEN1: deltaE9) Prp promoter4–9 months, Aβ plaques in the cortex and hippocampus Increased Experimental paradigm: in vivo video EEG recordings; PTZ-induced seizure susceptibility.
Findings: higher incidence of epileptiform-like discharges, i.e., seizure events (interictal spikes, sharp waves, or polyspikes) in APdE9 than in the controls. Also, APdE9 mice showed a lower latency to PTZ-evoked seizure events than in the control animals. A correlation was also found between the frequency of epileptiform-like discharges and the number of Aβ plaques.Busche et al. (2008) APP23xPS45
(hAPP751 Swedish and PSEN1-Gly384→Ala384, G384A)
Thy-1 promoter8–10 months, Aβ plaques Increased Experimental paradigm: in vivo two-photon Ca2+ imaging of neurons in layer 2/3 of the cortex.
Findings: clusters of hyperactive neurons were found in the vicinity of Aβ plaques.Busche et al. (2012) APP23xPS45
(hAPP751 Swedish and PSEN1-Gly384→Ala384, G384A)
Thy-1 promoter6–7 months, Aβ plaques Increased Experimental paradigm: in vivo two-photon Ca2+ imaging of CA1 pyramidal neurons in the hippocampus.
Findings: hyperactive neurons were found to be located exclusively in the vicinity of Aβ plaques in the hippocampus of transgenic mice.Lalonde et al. (2005) APP23
(hAPP751 Swedish)
Thy-1 promoter24 months, Aβ plaques Increased Experimental paradigm: behavioral seizures evaluation.
Findings: 41% of APP23 mice exhibited tonic-clonic seizures; 24% displayed myoclonic jumping.Jolas et al. (2002) TgCRND8
(hAPP695 Swedish and Indiana)
Prp promoter5 months, Aβ plaques Increased Experimental paradigm: in vitro hippocampal electrophysiology recordings; evoked EPSCs and IPSCs; PTZ-induced seizure threshold.
Findings: increased synaptic excitability and increased maximum amplitude of evoked mEPSCs; consistently lower dose of PTZ was required to elicit myoclonic activity (preseizure signs) in TgCRND8 mice compared with controls.Siwek et al. (2015) 5XFAD
(hAPP Swedish, Florida, and London plus PSEN1: M146L and L286V) Thy-1 promoter16.5 months, Aβ plaques throughout hippocampus and cortex Increased Experimental paradigm: in vivo video EEG recordings from the cortex and the hippocampus.
Findings: aberrant hyperexcitability in 5×FAD mice evidenced as ictal-like discharges, such as spikes, polyspikes, and spike-waves.Davis et al. (2014) 3xTg-AD
(hAPP Swedish. htau P301L, and hPSEN1: M146V)
Thy-1.2 promoter17–18 months, Aβ plaques Increased Experimental paradigm: in vivo hippocampal electrophysiology recordings.
Findings: increased synaptic excitability in DG and CA1.Nygaard et al. (2015) 3xTg-AD
(hAPP Swedish. htau P301L, and hPSEN1: M146V)
Thy-1.2 promoter8–10 months, Aβ plaques, cognitive impairment Increased Experimental paradigm: in vivo video EEG recordings from the cortex.
Findings: SWDs in 3×Tg-AD mice which correlated with spatial memory impairments.Chan et al. (2015) Tg2576
(hAPP Swedish), Prp promoter12–14 months, Aβ plaques, cognitive impairment Increased Experimental paradigm: electrical amygdala kindling with implanted electrodes and behavioral seizures evaluation.
Findings: Tg2576 mice exhibited increased susceptibility to kindling and seizure-associated death.Studies evaluating neuronal network hyperexcitability in hAPP/Aβ mice at early stages of Aβ pathology before plaque deposition and/or cognitive impairment Westmark et al. (2008) Tg2576
(hAPP Swedish), Prp promoter2 months, before Aβ plaques deposition and cognitive impairment Increased Experimental paradigm: PTZ-induced seizure susceptibility assessment.
Findings: increased susceptibility to PTZ-induced seizures in Tg2576 mice.Westmark et al. (2010) Tg2576
(hAPP Swedish), Prp promoter3 weeks, before Aβ plaques deposition and cognitive impairment Increased Experimental paradigm: audiogenic seizure susceptibility evaluation.
Findings: increased susceptibility to audiogenic seizures in Tg2576 mice as compared with WT controls. The audiogenic seizure susceptibility in Tg2576 mice could be suppressed by passive immunization with an anti-APP/Aβ antibody or by blockade of mGluR5 with the selective antagonist, MPEP.Corbett et al. (2013) Tg2576
(hAPP Swedish), Prp promoter5–7 months, before Aβ plaques deposition Increased Experimental paradigm: in vivo EEG recordings.
Findings: presence of SWDs and abnormal EEG patterns in Tg2576 mice; these mice also exhibited longer durations of higher frequency brain activity, suggesting increased synchrony.Bezzina et al. (2015) Tg2576
(hAPP Swedish), Prp promoter1.5–2 months, before Aβ plaques deposition and cognitive impairment Increased Experimental paradigm: electrical amygdala kindling with implanted electrodes and behavioral seizures evaluation.
Findings: Tg2576 mice exhibited increased susceptibility to kindling and seizure-associated death.Duffy et al. (2015) Tg2576
(hAPP Swedish), Prp promoter2–4 months, prior to Aβ plaques deposition; soluble Aβ40 and Aβ42 detectable; impairment in object location, an EC-dependent cognitive task. Increased Experimental paradigm: ex vivo EC recordings.
Findings: increased excitability in EC recordings in slices from Tg2576 mice.Kam et al. (2016) Tg2576
(hAPP Swedish), Prp promoter5 weeks, prior to Aβ plaques deposition and cognitive impairment Increased Experimental paradigm: in vivo video EEG recordings.
Findings: synchronized large amplitude potentials resembling interictal spikes in epilepsy were observed in Tg2576 mice.Ciccone et al. (2019) Tg2576
(hAPP Swedish), Prp promoter3 months, before Aβ plaques deposition Increased Experimental paradigm: extracellular fEPSP activity elicited by the proconvulsant drug 4-aminopyridine (4-AP) in acute hippocampal slices from 3-month-old WT and Tg2576 slices.
Findings: significantly higher number of electrical discharges, occurring with similar amplitude but shorter intervals, was observed in Tg2576 in comparison to WT hippocampal slices after 4-AP application.Del Vecchio et al. (2004) TgCRND8
(hAPP695 Swedish and Indiana)
Prp promoter6–8 weeks, before Aβ plaques deposition Increased Experimental paradigm: PTZ-induced seizure susceptibility evaluation.
Findings: increased susceptibility to PTZ-induced seizures in TgCRND8 mice.Fontana et al. (2017) PS2APP
(hAPP Swedish and hPSEN2: N141I)
Thy1 (hAPP) and Prp(hPSEN2) promoters3 months, before Aβ plaques deposition and cognitive impairment Increased Experimental paradigm: in vivo spontaneous LFPs in DG.
Findings: network hypersynchronicity was observed in the DG of PS2APP mice.Busche et al. (2012) APP23xPS45
(hAPP751 Swedish and PSEN1-Gly384→Ala384, G384A)
Thy-1 promoter1.5 months, before Aβ plaques deposition and cognitive impairment Increased Experimental paradigm: in vivo two-photon calcium imaging of the hippocampal CA1 neurons.
Findings: selective increase in hyperactive neurons in hippocampus of APP23xPS45 mice before Aβ plaques deposition suggesting that soluble species of Aβ may underlie this impairment. Acute treatment with the γ-secretase inhibitor LY-411575 reduced soluble Aβ levels and rescued the neuronal dysfunction.Davis et al. (2014) 3xTg-AD
(hAPP Swedish. htau P301L, and hPSEN1: M146V)
Thy-1.2 promoter4–6 months, before Aβ plaques deposition Increased Experimental paradigm: in vivo hippocampal electrophysiology recordings.
Findings: synaptic hyperexcitability in DG and CA1.Kazim et al. (2017) 3xTg-AD
(hAPP Swedish. htau P301L, and hPSEN1: M146V)
Thy-1.2 promoter3 weeks, before Aβ plaques deposition and cognitive impairment Increased Experimental paradigm: audiogenic seizure susceptibility; ex vivo hippocampal CA3 intracellular recordings after GABAA blockade with bicuculline.
Findings: increased audiogenic seizure susceptibility and prolonged epileptiform discharges after bicuculline application in hippocampal CA3 intracellular recordings in 3×Tg-AD mice.Fu et al. (2019) hAPP-J20
(APP Swedish and Indiana)
PDGF-β promoter1 and 2 months, before Aβ plaques deposition and cognitive impairment Increased Experimental paradigm: in vivo EEG recordings.
Findings: epileptic spikes at 1 month of age with robust seizure activity at 2 months of age.4-AP, 4-aminopyridine; Aβ, amyloid β; DG, dentate gyrus; EC, entorhinal cortex; EEG, electroencephalogram; fEPSPs, field EPSPs; hAPP, human amyloid β precursor protein; hPSEN, human presenilin; LFPs, local field potentials; LTP. Long-term potentiation; mEPSCs, miniature EPSCs; mGluR5, metabotropic glutamate receptor 5; mIPSCs, miniature IPSCs; MPEP, 2-methyl-6-(phenylethynyl)pyridine hydrochloride; PDGF, platelet-derived growth factor; Prp, prion protein; PTZ, phenylenetetrazole; PV, parvalbumin; REM, rapid eye movement; SWDs, spike-wave discharges; WT, wild type.
Author(s) and publication year Mouse model/transgene(s) Age/stage of pathology Neuronal network excitability status Experimental paradigm/neuronal network excitability observation(s) Rocher et al. (2010) rTg4510
(htau P301L)8.5 months, NFTs and neurodegeneration Increased Experimental paradigm: in vitro whole cell patch clamp recordings of layer 3 frontal cortex pyramidal neurons.
Findings: increased action potential firing rates and a significantly depolarized resting membrane potential in transgenic mice slices, independent of NFTs.Hoover et al. (2010) rTg4510
(htau P301L)
Rat hippocampal neurons transfected with hτP301LrTg4510 cultured hippocampal neurons from E18: DIV 22–30, decreased excitatory glutamate receptor levels.
Rat hippocampal neurons transfected with hτP301L: DIV 22–30, increased phospho-tauDecreased Experimental paradigm: in vitro hippocampal neurons electrophysiology, mEPSCs recording.
Findings: reduced mEPSCs frequency and amplitude both in rTg4510 cultured hippocampal neurons and rat hippocampal cultured neurons transfected with hτP301L.Crimins et al. (2011) rTg4510
(htau P301L)9 months, NFTs and neurodegeneration Increased Experimental paradigm: in vitro whole cell patch clamp recordings of layer 3 frontal cortex pyramidal neurons.
Findings: increased spontaneous synaptic activity (increased frequency of sEPSCs).Crimins et al. (2012) rTg4510
(htau P301L)< 4 (1–3) mo and > 8 (9–13) mo; soluble hyperphosphorylated tau species at <4 months, NFTs and neurodegeneration at >8 months Increased Experimental paradigm: in vitro whole cell patch clamp recordings of layer 3 frontal cortex pyramidal neurons.
Findings: increased excitability both in early and advanced tauopathy. Depolarized resting membrane potential, an increased depolarizing sag potential and increased action potential firing rates—all indicative of hyperexcitability. Hyperexcitability reversed by suppression of human mutant tau transgene.Menkes-Caspi et al. (2015) rTg4510
(htau P301L)3 months, accumulation of hyperphosphorylated and misfolded tau in cortex; 5 months, pathologic tau and NFTs in cortex Decreased Experimental paradigm: in vivo intracellular recordings from frontal cortex in anesthetized mice, In vivo extracellular recordings/LFPs in awake behaving mice.
Findings: reduced activity both of single neocortical pyramidal cells and of the neocortical network including decreased firing rates and altered firing patterns.Witton et al. (2016) rTg4510
(htau P301L)7–8 months, NFTs and neurodegeneration Increased Experimental paradigm: in vivo hippocampal CA1 electrophysiology recordings, both single-unit and LFPs.
Findings: increased propensity of excitatory pyramidal neurons in hippocampus to fire action potentials in a phase locked manner during SWRs; inhibitory interneurons were less likely to fire phase‐locked spikes during SWRs.Hatch et al. (2017) rTg4510
(htau P301L, 13-fold higher human tau expression as compared with endogenous tau)
pR5
(htau P301L, at lower level than rTg4510, 0.7-fold higher human tau as compared with endogenous tau)rTg4510: 1–2 months, early stage tauopathy before overt tau hyperphosphorylation and synaptic impairment
4–6 months, mid-stage with extensive tau hyperphosphorylation and impairment of synaptic activity and spatial memory
12–14 months, late stage with synaptic loss and neurodegeneration
PR5: 15–17 months, tau pathology in hippocampusDecreased Experimental paradigm: in vitro whole cell patch clamp recordings from hippocampal CA1 pyramidal neurons.
Findings: reduced action potential firing rate because of a depolarization shift in action potential generation and reduced action potential amplitude at all ages in the CA1 pyramidal neurons of P301L mice. pR5 mice CA1 pyramidal neurons showed less severe action potential impairment compared with rTg4510, including action potential depolarization shift and reduced action potential amplitude.Busche et al. (2019) rTg4510
(htau P301L)
rTg21221
(htau overexpression)rTg4510: 6–12 months, tau aggregation and NFTs
3–4 months; soluble tau.
rTg21221: 6–12 months, human tau overexpression.Decreased Experimental paradigm: in vivo two-photon Ca2+ imaging of neurons in layer 2/3 of the cortex.
Findings: strong reduction in cortical activity as reflected by increased number of silent neurons in both 6- to 12- and 3- to 4-month-old rTg4510 mice. Reduction in cortical activity in 6- to 12-month-old rTg2210 mice. Reducing tau in 3- to 4-month-old rTg4510 mice decreased the number of silent neurons.Van Erum et al. (2020) tau58/4
(htau P301S)3 months, htau and phospho-tau in the frontal cortex and pons
12–15 months, NFTs throughout the brain.Increased Experimental paradigm: video EEG recordings; PTZ-induced seizure susceptibility.
Findings: increased PTZ-induced seizure susceptibility in young (3-month-old) tau58/4 mice as compared with age-matched WT littermates. Young tau58/4 animals displayed more severe seizures and had a reduced latency to the first seizure compared with WTs. While, age-related differences in susceptibility could be demonstrated for both genotypes, old tau58/4 did not exhibit a significantly higher seizure susceptibility as compared with WT mice.Marinković et al. (2019) P301S
(htau P301S)2 m, injected with tau preformed fibrils for NFTs seeding, evaluated for cortical activity up to 50 d after injections Decreased Experimental paradigm: in vivo two-photon Ca2+ imaging of neurons in layer 2/3 of the cortex in awake, head-fixed mice.
Findings: strong reduction in cortical activity, independent of NFTs presence, suggesting the impairing role of soluble, mutated tau protein species.Mondragón-Rodríguez et al. (2018b) 3xTg-AD
(htau P301L, hAPP Swedish, hPSEN1)1 month, increased phospho-tau, intraneuronal APP/Aβ, prior to cognitive impairment. Decreased Experimental paradigm: in vitro whole cell patch clamp recordings from hippocampal CA1 pyramidal neurons; in vitro LFPs recordings from hippocampal slices.
Findings: no difference in amplitude and frequency of action potentials between 3×Tg-AD and non-Tg CA1 pyramidal neurons. Overall, the young 3×Tg-AD mice showed less excitable hippocampal network activity, likely related to abnormally hyperphosphorylated tau at microtubule domain region (MDr).Ahnaou et al. (2017) Tg tau P301L
(h tau P301L)3 months, injected with preformed tau fibrils to induce tau aggregation. Normal Experimental paradigm: in vivo EEG recordings. Network oscillations, phase amplitude cross frequency coupling, mismatch negativity (MMN) of event related brain potentials, and coherence was analyzed.
Findings: weakening of θ oscillations, drastic impairments in θ–γ oscillations phase-amplitude cross frequency coupling, and disrupted MMN complex amplitude (all vital for memory and learning performance) induced by tau seeding. No epileptiform activity or network hyperexcitability.Angulo et al. (2017) EC-htau
(hτP301L)
EC-hAPP
(hAPP)
EC-hAPP/htau
(hAPP, hτP301L)EC-htau: 2.5–3.5 months, htau and phospho-tau accumulation in EC.
EC-hAPP: 2.5–3.5 months, soluble Aβ.Normal/
resistance to induced-hyperexcitability in EC-htau miceExperimental paradigm: in vitro hippocampal EC/subiculum electrophysiology recordings: single electrode evoked and sEFPs, single neuron patch clamp, and extracellular multielectrode recordings.
Findings: mutated htau induced resistance to EC-hippocampus hyperexcitability in EC-htau mice evidenced by resistance to increased network activity evaluated by sEFP durations after GABAA blockade with picrotoxin. Increased neuronal excitability in EC in EC-hAPP mice evidenced by higher frequency of relatively prolonged sEFPs in lateral EC and epileptiform-ictal like discharges in medial EC. While no differences were observed in sEFPs duration and frequency in EC/CA1/subiculum regions (using multielectrode recordings) between EC-hAPP/htau mice and WT mice, a smaller percentage of slices displayed epileptiform discharges. Co-expression of hAPP and htau produced an intermediate phenotype, mostly driven by tauMaeda et al. (2016) htau-A152T 4–9 months, soluble tau Increased Experimental paradigm: in vivo EEG recordings in awake behaving mice; epileptic spikes quantification both at baseline and after injection of non-convulsive dose of PTZ.
Findings: increased epileptic spike counts both at baseline and after PTZ injection in htau-A152T mice as compared with non-Tg controls.Decker et al. (2016) htau-A152T 12–14 months, accumulation of hyperphosphorylated and missorted tau, neurodegeneration, and synaptic loss in hippocampal CA3 region. Increased phospho-tau in hippocampal slice cultures at DIV 10. Increased Experimental paradigm: in vivo EEG recordings in awake behaving mice; epileptic spikes quantification both at baseline and after injection of non-convulsive dose of PTZ.
Findings: enhanced basal synaptic transmission in CA3 region of the hippocampus in htau-A152T mice (increased fEPSPs in mossy fiber pathway in acute slices from 12-month-old mice and increased somatic field potentials in stratum pyramidale of area CA3 in organotypic hippocampal slices at DIV 30. Increase in picrotoxin-induced epileptiform burst frequency as well as in firings per burst in organotypic slice cultures expression htau-A152T mutation (both at DIV 10 and 30) which was prevented by ceftriaxone (stimulates astrocytic glutamate uptake via the transporter EAAT2/GLT1).Das et al. (2018) htau-A152T 4–6 months; soluble tau Increased Experimental paradigm: in vivo EEG recordings; epileptic spikes quantification.
Findings: increased epileptic spike counts at resting state in htau-A152T mice as compared with non-Tg controls. Antiepileptic drug levetiracetam treatment reduced epileptic spike counts in htau-A152T mice.García-Cabrero et al. (2013) FTDP-17 mice
(htau G272V, P301L, and R406W; Overexpression of human tau isoform with 2 N-terminal inserts, 4-microtubule-binding-repeat elements)1–5, 6–14, and 15–22 months; mutant tau transgene overexpressed at 3 m of age, activated microglia at 4 months, reactive astrocytes at 9 months, and phospho-tau aggregates at 18–20 months Increased Experimental paradigm: in vivo video EEG recordings and PTZ-seizure susceptibility testing.
Findings: spontaneous epileptiform activity and epileptic seizures in 70% of FTDP-17 mice at the age of 5.5 m and thereafter.
Increased PTZ-induced seizures susceptibility at 6–14 months of age and thereafter.Aβ, amyloid β; EC, entorhinal cortex; DIV, days in vitro; EAAT2, excitatory amino acid transporter 2; EEG, electroencephalogram; fEPSPs, field EPSPs; FTDP, frontotemporal dementia with parkinsonism; GLT1, glutamate transporter 1; hAPP, human amyloid β precursor protein; hPSEN, human presenilin; LFPs, local field potentials; mEPSCs, miniature EPSCs; NFTs, neurofibrillary tangles; PTZ, phenylenetetrazole; sEFPs, spontaneous extracellular field potentials; sEPSCs, spontaneous EPSCs; SWDs, spike-wave discharges; SWRs, sharp-wave ripples; Tg, transgenic; WT, wild type.
- Table 3
Summary of studies analyzing the effect of tau reduction on network excitability in Aβ mice and in WT mice
Author(s) and publication year Mouse model/
transgene(s)Age/stage of pathology Baseline neuronal network
excitability statusExperimental paradigm and tau reduction strategy/effect of tau
reduction on neuronal network excitabilityRoberson et al. (2007) hAPPJ20/tau+/+,
hAPPJ20/tau+/−,
hAPPJ20/tau−/−,
tau+/+, tau+/−,
tau−/−.hAPP(J20): hAPPSwe, hAPPInd, PDFG promoterhAPP/tau+/+: 4–7 months, Aβ plaques, neurodegeneration Increased in hAPP/tau+/+ Experimental paradigm: in vivo PTZ-induced and kainate-induced seizure susceptibility evaluation. tau reduction achieved through crossing hAPPJ20 line with tau knock-out (tau−/−).
Findings: tau reduction increased resistance to both PTZ-and kainate-induced seizures. Seizures were less severe in hAPP/tau+/– and hAPP/tau–/– mice than in hAPP/tau+/+ mice. Seizures were also less severe in tau–/– mice than in tau+/+ mice. The onset of seizure was also delayed by tau reduction. tau reduction also ameliorated learning and memory deficits in hAPPJ20 mice.Roberson et al. (2011) hAPPJ20/tau+/+, hAPPJ20/tau+/−, hAPPJ20/tau−/−,
hAPPJ9/tau+/+, hAPPJ9/tau−/−
tau+/+, tau−/−.7–14 months for in vivo EEG detection of frequency of epileptiform spikes: Aβ plaques, neurodegeneration.
5–8 months for in vivo PTZ-induced seizure susceptibility evaluation: Aβ plaques, neurodegenerationIncreased in hAPPJ20/tau+/+ and hAPPJ9/tau+/+ mice Experimental paradigm: in vivo PTZ-induced seizure susceptibility evaluation. In vivo EEG detection of frequency of epileptiform spikes in freely moving mice. In vitro epileptiform discharges in area CA1 of the hippocampus after bicuculline administration in acute slices. tau reduction achieved via crossing hAPPJ20 or hAPPJ9 line with tau knock-out (tau−/−) mice.
Findings: tau reduction decreased PTZ-induced seizure severity and frequency of generalized epileptiform spikes in hAPPJ20 mice. tau reduction also prevented bicuculline-induced epileptiform bursting in acute hippocampal slices from WT (tau−/−) and hAPPJ20 mice.DeVos et al. (2013) WT (C57BL/6J),
tau−/−3–5 months, no pathology Normal Experimental paradigm: in vivo EEG recordings, baseline and after picrotoxin administered via reverse microdialysis. In vivo PTZ-induced seizure susceptibility evaluation. tau reduction was achieved via ASOs.
Findings: reduction in normalized spike frequency after picrotoxin administration in ASO-treated WT mice and tau−/− as compared with controls. Total tau protein levels in the hippocampus of mice highly correlated with normalized spike frequency. PTZ-induced seizure severity was significantly reduced in ASO-treated WT mice. Seizure severity and tau protein levels correlated well in all tested mice.Li et al. (2014) tau+/+, tau+/−,
tau−/−.
(all on C57Bl/6J background)24 months, no pathology, age-appropriate cognitive function Normal Experimental paradigm: in vivo PTZ-induced seizure susceptibility evaluation. tau reduction achieved via genetic homozygous or heterozygous knock-out.
Findings: PTZ-induced seizure severity was significantly reduced in tau knock-out aged mice. Also, aged tau+/− and tau−/− mice had longer seizure latencies than tau+/+ mice.Ittner et al. (2010) APP23 (APPSwe, Thy1 promoter),
Δtau74 (amino acids 256–441 removed from the longest human tau isoform, htau40),
tau−/−, Δtau74.tau−/−,
APP23. Δtau74,
APP23. tau−/−,
APP23. Δtau74.tau−/−APP23: 2–3 months, no plaques.
Δtau74: 2–3 months, tau missorting, normal endogenous tau but negligible phospho-tauIncreased in APP23 mice Experimental paradigm: in vivo PTZ-induced seizure susceptibility evaluation. tau reduction achieved via crossing APP23 line with Δtau74 or tau−/− mice.
Findings: seizure severity was significantly reduced in Δtau74, tau−/−, and Δtau74. tau−/− compared with the WT, while the latency to develop severe convulsion was increased. APP23 mice presented with a reduced convulsion latency and showed the most severe seizure response. However, when APP expression was combined with Δtau expression or tau deficiency, this significantly decreased seizure severity, reduced fatality, and increased convulsion latency. The double mutant Δtau74.tau−/− prevented severe seizures better than Δtau74 or tau−/−alone, on both WT and APP23 backgrounds.Aβ, amyloid β; ASOs, antisense oligonucleotides; EEG, electroencephalogram; hAPP, human amyloid β precursor protein; PDGF, platelet-derived growth factor; PTZ, phenylenetetrazole; WT, wild type.
In this issue

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}




Jump to section
- Article
- Abstract
- Significance Statement
- Introduction
- Aβ Induces Neuronal Network Hyperexcitability in AD, Even before the Development of Plaques
- The Role of tau in Neuronal Network Excitability: The Enhancement versus Suppression Conundrum
- Perspectives on the Similar versus Divergent Roles of Aβ and tau in Neuronal Network Hyperexcitability in AD: Which One Has a Dominant Effect, Aβ or tau?
- Future Directions and Concluding Remarks
- Acknowledgments
- Footnotes
- References
- Synthesis
- Figures & Data
- Info & Metrics
- eLetters