

Article Figures & Data
Figures
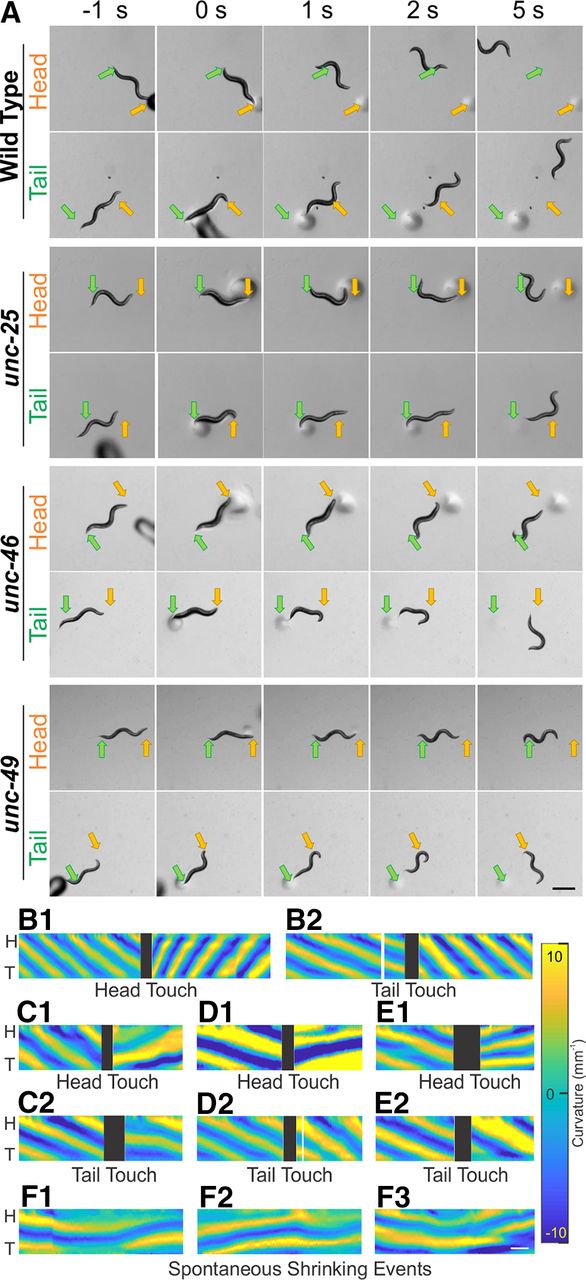
- Figure 1.
Wild-type and GABA transmission knock-outs shrank after harsh touch to the head or tail, while wild-type animals escaped rapidly. A, While moving forward, animals were touched by a blunt glass probe as a harsh stimulation to the head (yellow arrow) or tail (green arrow). Wild-type animals briefly shrank immediately after stimulation (compare 0 s to before stimulation (-1 s) and rapidly moved away from the stimulus, while GABA transmission knock-outs shrank for a longer time and moved away slowly. Note that by 5 s, all animals had moved away from the stimuli. Common scale bar is 0.5 mm. Dark and circular shadows are the glass probe and the marks it left on agar. After harsh stimulation, wild-type animals (B) increased undulation frequency to escape backward (B.1) or forward (B.2). In contrast, all three GABA transmission mutants, unc-25 (C), unc-46 (D), and unc-49 (E), decreased undulation frequency when they moved away from the stimuli. Moreover, there was a period in which the body posture of these mutants did not change after the stimulation (C–E). F, The mutants, unc-25 (F.1), unc-46 (F.2), and unc-49 (F.3), occasionally shrank without stimulation. Yellow and blue shaded areas in B–F represent dorsoventral curvature (vertical axis is the length of animal from the head (H) on the top to the tail (T) color bar is curvature -10–10 mm-1, indifferent to dorsoventrality), along time (horizontal axis, common scale bar is 1 s); black blocks indicate gap in tracking during the touch stimulation.
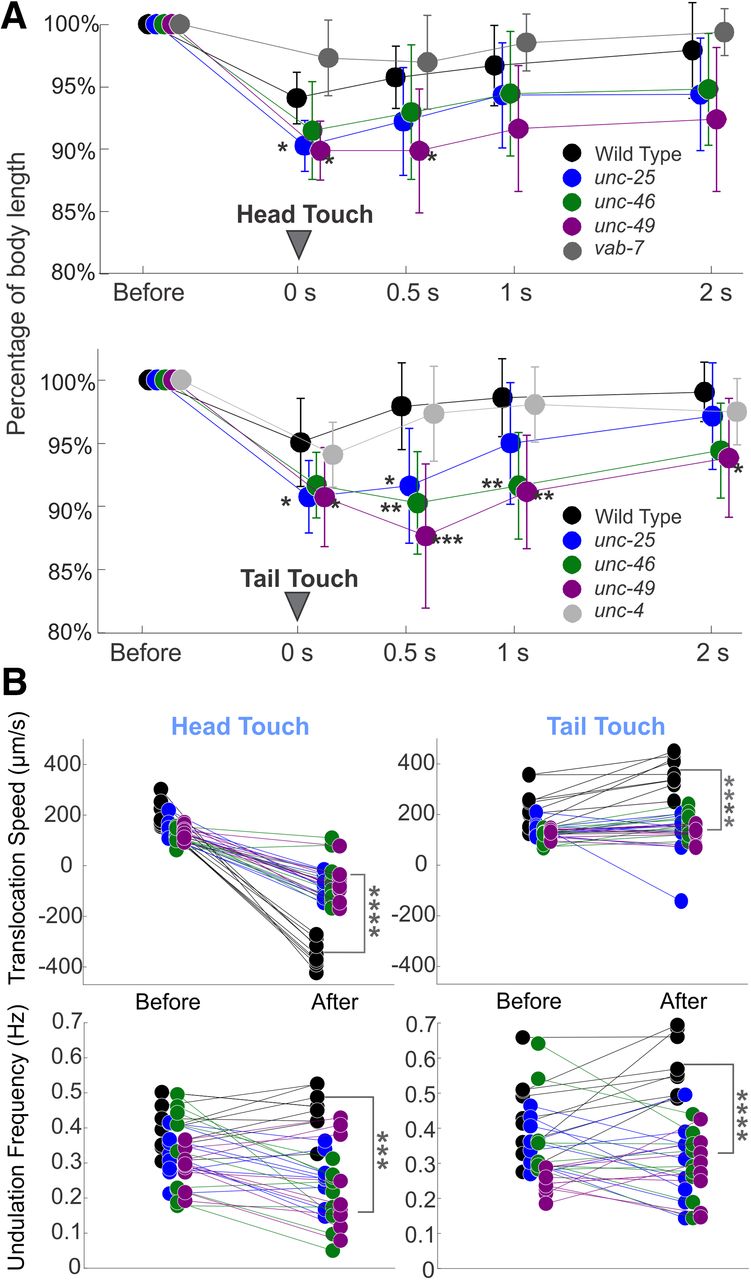
- Figure 2.
Both wild-type and GABA transmission knock-out animals shrank after harsh touch stimuli; wild-type animals shrank less, recovered sooner, and crawled away more rapidly than knock-out animals. A, Wild-type and GABA transmission knock-out animals decreased their body length after harsh stimuli to the head or tail (0 s). Compared with wild-type, GABA transmission mutants decreased their body length more and recovered more slowly (0.5, 1, and 2 s after stimuli). The body length change of vab-7 and unc-4 were not statistically different from that of wild type. Body length is normalized to prestimulus value for each animal. See Extended Data Figure 2-1 for details. B, Wild-type animals moved with higher mean undulation frequency and mean translocation speed during 5 s poststimulus compared with the 5 s prestimulus, while the mutants did not. Two unc-46 mutant animals did not change locomotion direction after harsh head touch, neither did one unc-25 mutant animal after harsh tail touch. In all plots, one-way ANOVA with Tukey’s pairwise comparison to wild-type; *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001. Black circles: wild-type, blue: unc-25, green: unc-46, and purple: unc-49. See Extended Data Figures 2-2, 2-3 for details.
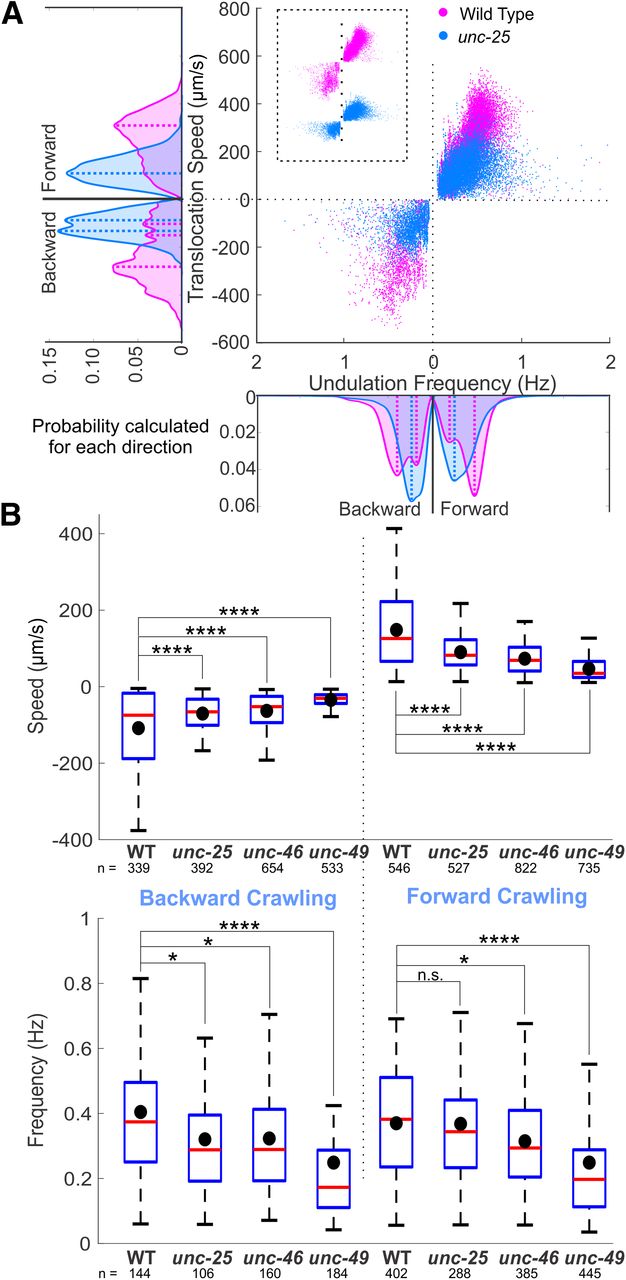
- Figure 3.
GABA transmission mutants crawled more slowly than wild type. A, unc-25 knock-out animals crawled on agar surface with lower translocation speed and undulation frequency than wild type. Negative translocation speed and undulation frequency left (and right) of 0 represent backward (and forward) locomotion. Area plots show the probability of occurrence for each direction with dashed lines indicating the main peaks in the area plots. Probabilities were generated from histograms with bin size of 20 μm/s and 0.02 Hz translocation speed and undulation frequency, respectively. B, Translocation speeds of all the GABA transmission knock-outs were significantly lower than that of wild-type (WT) during forward and backward crawling. The undulation frequencies of GABA transmission knock-outs were significantly lower than those of WT animals, except for that of unc-25 during forward crawling. Blue boxes: first to third quartiles of each dataset; red lines: medians; black dots: mean values; whiskers: minimum and maximum, excluding outliers (beyond 1.5-fold interquartile range from middle 50% data). ANOVA with Tukey’s pairwise comparison to WT, n.s. p > 0.05, *p < 0.05, ***p < 0.001, ****p < 0.0001. See Extended Data Figure 3-1 for details and Extended Data Figure 3-2 for shared-control Gardner–Altman plots.
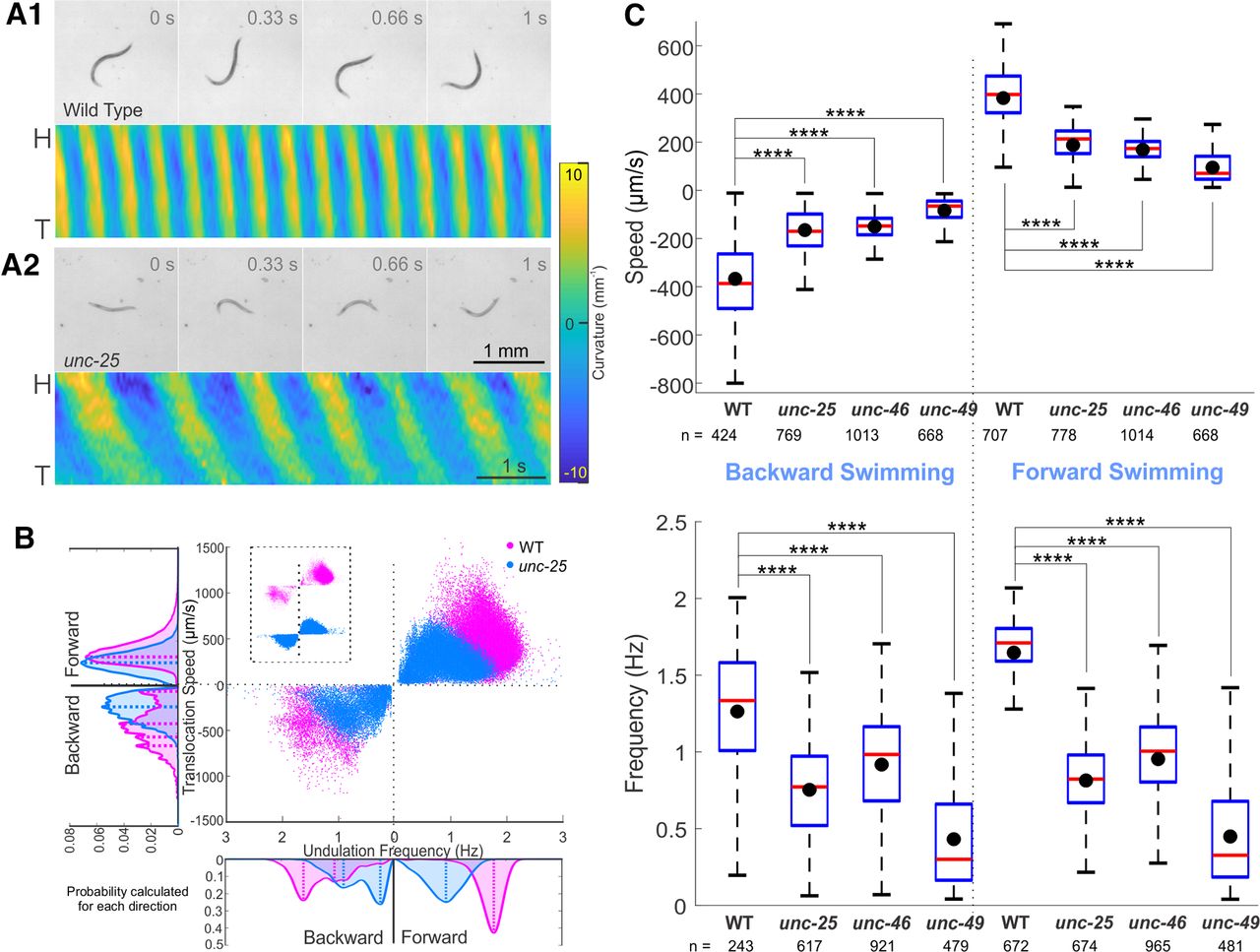
- Figure 4.
GABA transmission knock-outs swam more slowly than wild type. Compared with wild-type (A.1), unc-25 knock-out (A.2) in liquid swam with lower undulation frequency and translocation speed, as well as smaller undulation amplitude and wavelength. Video frames are 0.33 s apart, common scale bar is 1 mm; yellow and blue shaded areas of kymograms represent dorsoventral curvature [vertical axis is the length of animal from the head (H) on the top to the tail (T) color bar is curvature -10–10 mm-1, indifferent to dorsoventrality], along time (horizontal axis, scale bar is 1 s). B, unc-25 knock-out animals swam slower with lower translocation speed and undulation frequency than wild type. Area plots show the probability of occurrence for each direction with dashed lines indicating the main peaks in the area plots. Probabilities were generated from histograms with bin size of 24 μm/s and 0.05 Hz translation speed and undulation frequency, respectively. C, Translocation speeds and undulation frequencies of all the GABA transmission knock-outs were significantly lower than those of wild-type (WT) during forward and backward swimming. Box plots as in Figure 3. ANOVA with Tukey’s pairwise comparison to WT, ****p < 0.0001. See Extended Data Figure 4-1 for details and Extended Data Figure 4-2 for shared-control Gardner–Altman plots.
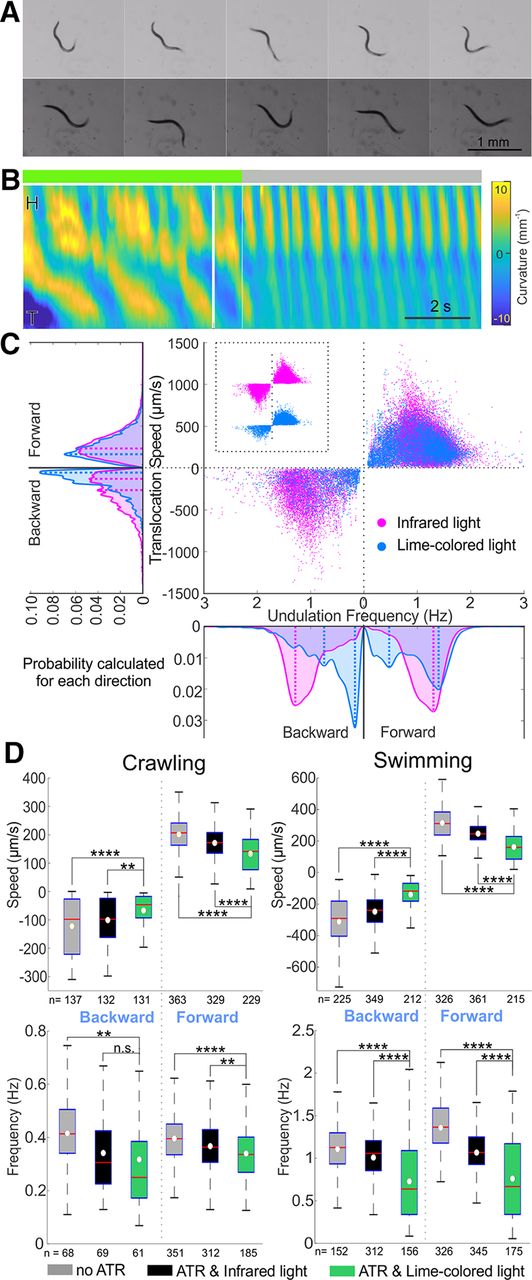
- Figure 5.
Acute optogenetic inactivation in GABAergic motoneurons induced slow locomotion during free swimming and crawling. A, When GABAergic motoneurons were inactivated optogenetically (top framed and green line on top of kymogram) while swimming in liquid, undulation was slower and more uncoordinated. When the optogenetic inhibition was withdrawn (bottom frames and gray line on top of kymogram), the animal regained coordinated undulatory swimming and its undulation frequency doubled from 0.5 to 1 Hz. Postures shown at 0.33 s intervals; scale bar is 1 mm; yellow and blue shaded areas of kymograms represent dorsoventral curvature (vertical axis is the length of animal from the head on the top to the tail, color bar is curvature -10–10 mm-1, indifferent to dorsoventrality), along time (horizontal axis, scale bar is 1 s). B, The optogenetic strain fed with ATR swam slower when optogenetically inactivated (by lime-colored light) compared with the same animals under infrared light. Area plots show the probability of occurrence for each direction with dashed lines indicating the main peaks in the area plots. Probabilities were generated from histograms with bin size of 24 μm/s and 0.05 Hz translation speed and undulation frequency, respectively. C, When GABAergic motoneurons were acutely inhibited (green boxes, ATR and lime-colored light), animals crawled and swam at a lower translocation speed and with lower undulation frequency compared with two negative controls, the same animal under infrared light (black boxes) and the same strain not fed ATR (gray boxes); except for backward-crawling undulation frequency that was not statistically different under lime-colored and infrared light. Boxplots similar to Figure 3. ANOVA with Tukey’s pairwise comparison to the animal fed with ATR and under lime-colored light, n.s. p > 0.05, ** p < 0.01, **** p < 0.0001. See Extended Data Figure 5-1 for details and Extended Data Figures 5-2, 5-3 for shared-control Gardner–Altman plots.
- Figure 6.
Undulation frequency correlated with translocation speed during free locomotion and undulation frequencies vary with the ambient viscosity in microfluidic waveform channels. A, B, Undulation frequency was positively correlated with translocation speed during crawling and swimming. Wild-type (magenta dots) and unc-25 (cyan dots) data are during episodes in which animals performed sinusoidal locomotion. Fitted linear regression lines of wild-type (black), unc-25 (blue), unc-46 (green), and unc-49 (purple) strains; coefficients of determination (R2) and equations are labeled with the same colors. C, In waveform microfluidic devices, the animals can undulate in predetermined channels designed to restrict their path with corresponding sequences of postures. When the ambient viscosity was high (3% w/w methylcellulose solution) wild-type animals undulated with lower undulation frequencies than in less viscous environment (0.5% w/w methylcellulose solution). Frequency was measured by tracking the motion of neurons and muscle cells during calcium imaging. Sample size (n) is the number of cells analyzed. D, Same data as C. The mean undulation frequency in 3% methylcellulose solution was 0.28 ± 0.14, and that in 0.5% solution was 0.74 ± 0.26 Hz. Red lines: mean values; pink boxes: 95% confidence intervals for the mean; gray and blue boxes: standard deviation; and gray dots: individual data points.
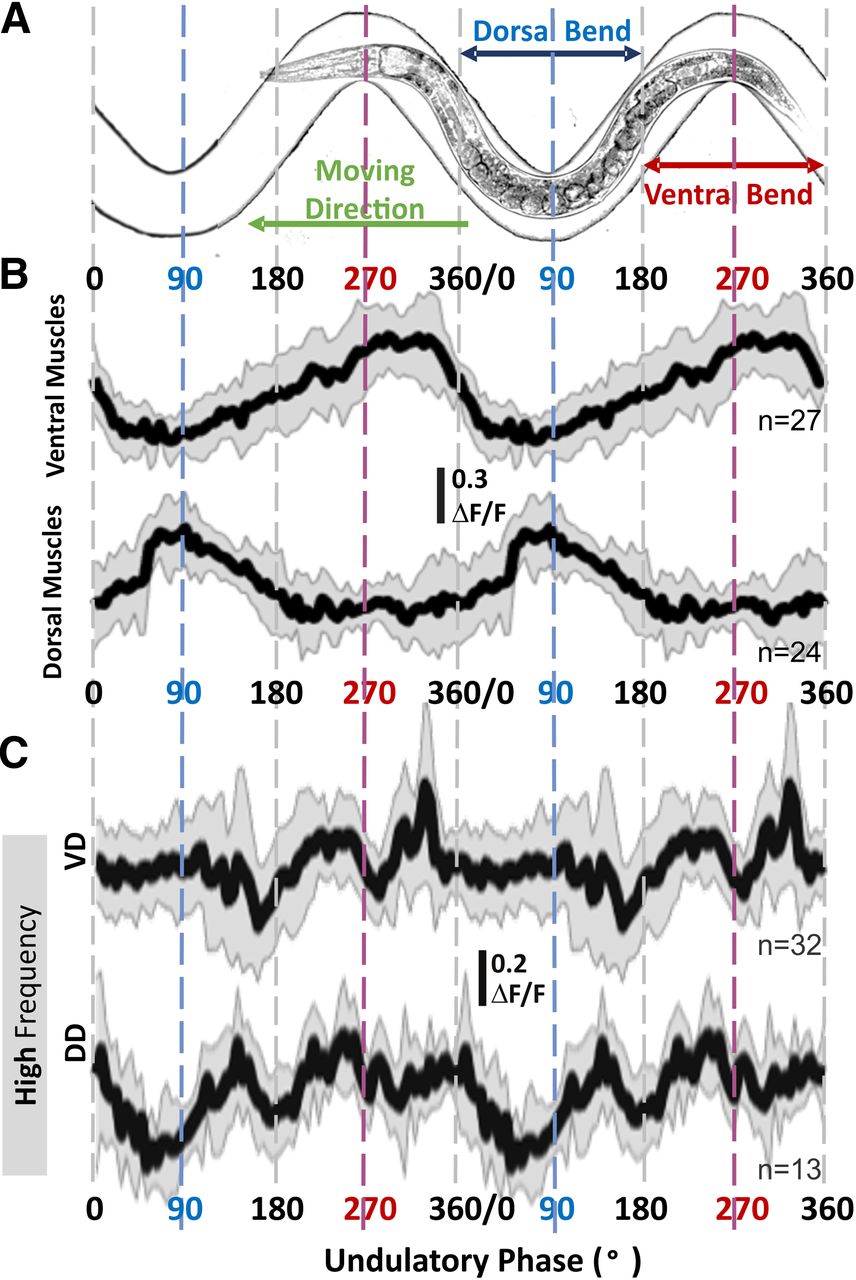
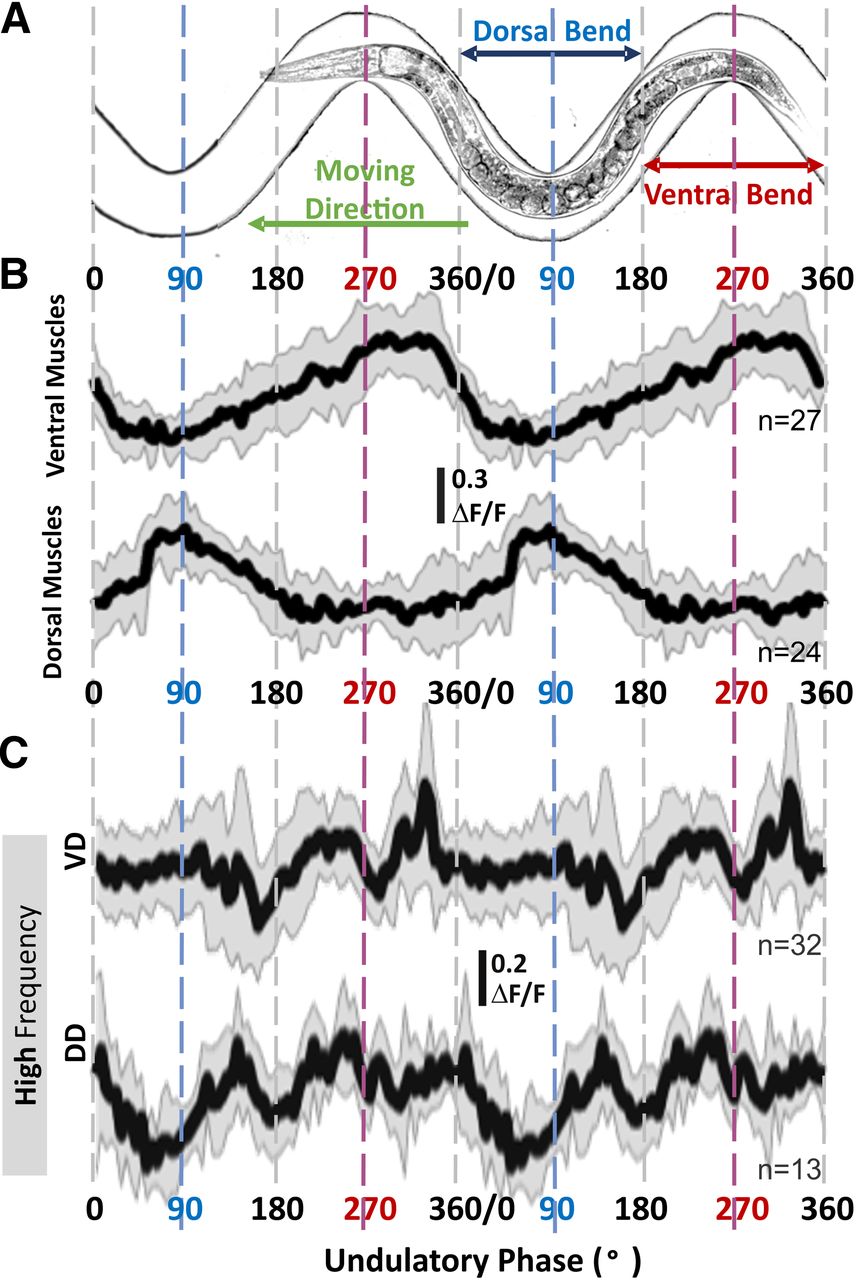
- Figure 7.
Calcium signal from GABAergic motoneurons related to locomotive phase and muscle activity only during high-frequency undulation. A, Animals were free to move in sinusoidal microfluidic channels so that the phase of their undulatory cycle, including dorsal and ventral bends, is fixed and imposed by their spatial location. B, Dorsal and ventral body-wall muscle cells were active during bending of the same side of the body (e.g., dorsal muscles were active during dorsal bend around 90°) regardless of undulation frequency. C, During crawling at high-frequency undulation, VD showed two troughs around 180° and 270°, and DD around 90° and 180°; only VD showed a sharp increase around 300°. The sample size (n) indicates the number of analyzed cells; solid lines and shaded areas in calcium traces are the mean and standard deviation. ΔF/F = (Ftop50% – FBG)/(Σ(Ftop50% – FBG)/n). Changes in fluorescence levels and the perimotor-corrected locomotor phase were analyzed and calculated with a customized MATLAB program (Extended Data 1).
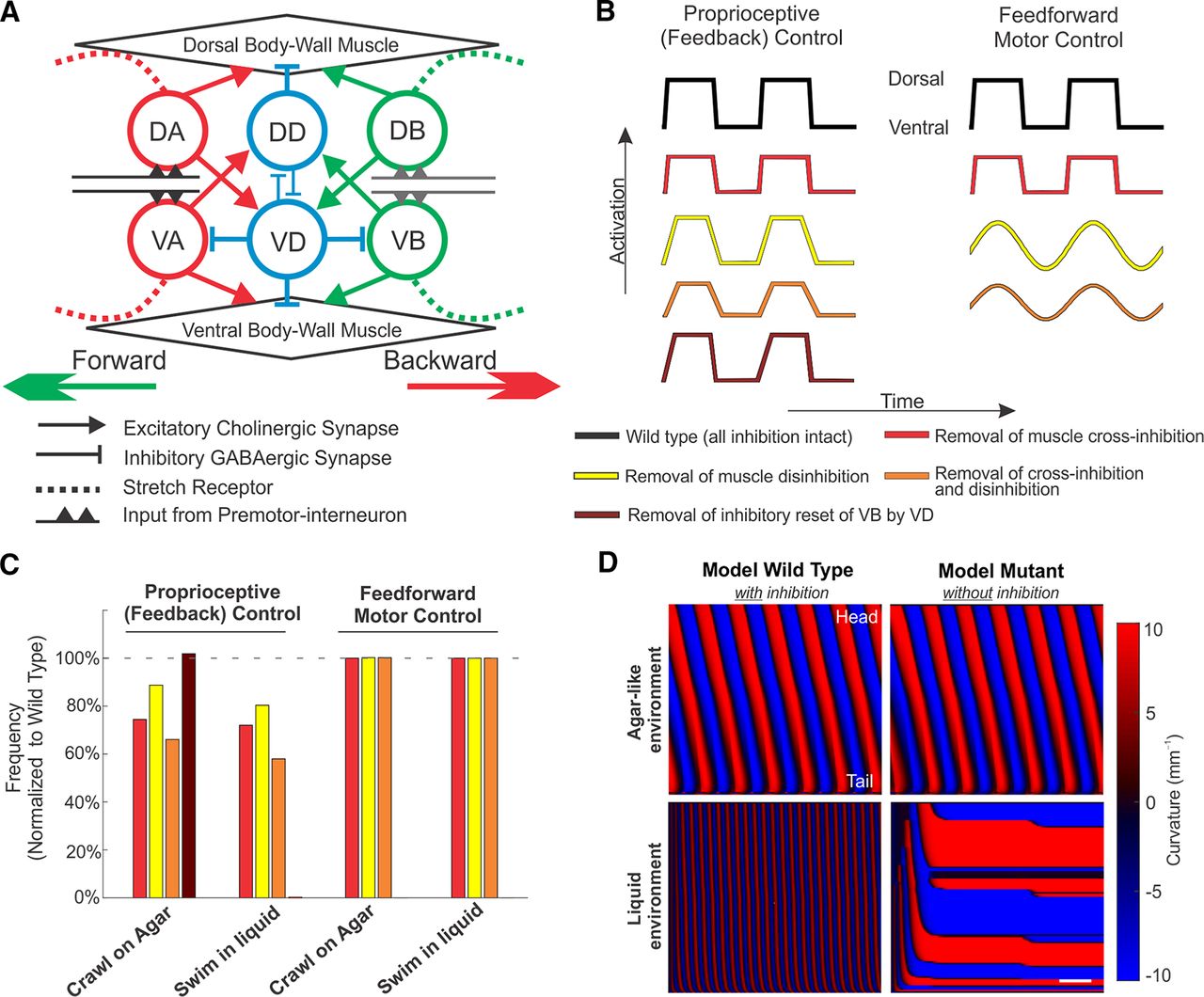
- Figure 8.
Integrated neuromechanical computational models test postulated roles of inhibition in proprioceptive (feedback) driven pattern generation, as compared with CPG (purely feedforward) models. A, Repeating units of the neuromuscular circuit were used to model C. elegans locomotion control. Single-unit schematic motor circuit of the ventral nerve cord used in our proprioceptive model of locomotion. During forward locomotion, VB and VD (DB and DD) motoneurons innervate ventral (dorsal) muscles and VD motoneurons innervate VB motoneurons. During backward locomotion, A-type motoneurons take the place of B-type motoneurons. Postulated mechanosensation (dash lines) encode a proprioceptive signal by integrating the extent of bending over its receptive field. Suprathreshold proprioceptive currents trigger the activation of VB and DB motoneurons during forward locomotion (or VA and DA motoneurons during backward locomotion). The reciprocal synaptic connections between VD and DD motoneurons (blue thin lines) are not considered in the model. Adapted from Cohen and Denham (2019; their Fig. 1B). B, Schematic traces of muscle activation in the proprioceptive control model (left) and feedforward control model (right), under different model perturbations. Positive activation denotes dorsal muscle activation. By construction, dorsal and ventral activation are in exact antiphase. Three hypotheses for the role of inhibition (cross-inhibition of the opposing muscles; disinhibition of the innervated muscles; and inhibitory reset of VB by VD) were tested in proprioceptive models of motor control; postulated roles of muscle cross-inhibition and disinhibition were tested in feedforward models (parameters and parameter sweeps in Extended Data Figs. 8-1, 8-2, respectively). When muscle inhibition is disrupted, the amplitude or waveform of muscle input is modified from the model wild type (black). Shown are schematics of perturbations corresponding to hypothesis 1 (reduced amplitude, red), hypothesis 2 (slower/smoother waveform, yellow), hypotheses 1 and 2 combined (orange), and hypothesis 3 (removal of neural inhibition, brown, proprioceptive model only). C, Relative frequency changes because of model perturbations. Under models of proprioceptive control, the undulation frequency is reduced by ∼10–35% relative to the wild type under a single perturbation to muscle inhibition, or up to ∼40% under the combined perturbation, suggesting that sufficient muscle activation amplitude and rapid activation onset are both required to sustain rapid undulation, during both crawling and swimming. Under proprioceptively driven locomotion, VB inhibition by VD serves to reset the neural rhythm during rapid undulation but has negligible effect in slow crawling-like motion. Bar plots with color scheme as in B depict the undulation frequency of model mutants normalized by the respective frequency of the model wild type. D, Under a model of proprioceptive control, inhibitory reset is necessary for rapid swimming in a liquid environment (bottom), but not for slow crawling on agar-like environments (top). In an agar-like environment, the elimination of VD-to-VB inhibition (right) has no effect. In a water-like environment, elimination of this neural inhibition eliminates the rhythmic pattern altogether. Red and blue shaded areas represent dorsoventral curvature; vertical axis is the length of animal from the head (H) on the top to the tail (T); color bar is ventral (blue) to dorsal (red) curvature -10–10 mm-1; along time (horizontal axis, common scale bar is 1 s).


Extended Data
Extended Data Figure 2-1
Harsh touch stimulation to the head or tail induces shortening of body length in wild-type and a larger shortening in GABAergic mutants. Body length of wild-type and GABA transmission knock-out animals (unc-25, unc-46, unc-47, and unc-49) reduced after harsh stimuli to the head or the tail, but length reduced less and recovered sooner in wild-type animals. Percentage of the body length at four time points: right away (0 s), 0.5, 1, and 2 s, was compared to the prestimulus value. Statistical tests compared mutants to wild-type (ANOVA and Tukey) as well as poststimulus to prestimulus values (paired t test). Calculated probabilities of the null hypotheses (p value) below 0.05 were considered significant and are in red. Download Figure 2-1, DOCX file.
Extended Data 1
Code used in this study in three folders: (1) MATLAB program to plot curvature kymograms from hdf5 file generated by Tierpsy. (2) MATLAB program to analyze the change in fluorescence intensity of identifiable body-wall muscle cells or somata of motoneurons. (3) MATLAB code of computational models. Download Extended Data 1, ZIP file.
Extended Data Figure 2-2
Wild-type animals move faster in response to head or tail touch while GABAergic mutant strains move slower or unchanged. After either head or tail harsh touch, wild-type animals (n = 9) moved away at significantly higher translocation speed, undulation frequency, and amplitude, while the mutant animals (n = 9 for each strain) moved with unchanged or lower translocation speed and undulation frequency, compared to their locomotion before the stimulation. All the animals performed deeper bending with higher amplitude after stimulation while their wavelength did not change. Calculated probabilities of the null hypotheses (p value) below 0.05 were considered significant and are in red. Download Figure 2-2, DOCX file.
Extended Data Figure 2-3
GABAergic mutant strains move at lower speed and frequency than wild-type 5 s poststimulus. Five seconds after a harsh touch to the head or tail, all GABAergic knock-out animals moved with lower translocation speed and undulation frequency than wild type. Calculated probabilities of the null hypotheses (p value) below 0.05 were considered significant and are in red. Download Figure 2-3, DOCX file.
Extended Data Figure 3-1
During free crawling, GABAergic mutants move slower than wild type and with smaller amplitude and wavelength. Animals of three GABA transmission knock-out strains crawled on agar surface with significantly lower translocation speed, undulation frequency, maximal amplitude, and primary wavelength. The only exception is that unc-25/GAD knock-out animals move at the same mean frequency as wild type when moving forward. Calculated probabilities of the null hypotheses (p value) below 0.05 were considered significant and are in red. Download Figure 3-1, DOCX file.
Extended Data Figure 3-2
During free crawling, GABAergic mutants move at lower frequency and slower than wild type in both forward and backward directions. Same data as in Figure 3B presented in shared-control Gardner–Altman plots with raw data (upper axes) and mean differences (lower axes: bootstrap distributions, mean difference are dots; 95%CI are vertical error bars). Download Figure 3-2, TIF file.
Extended Data Figure 4-1
During free swimming, GABAergic mutants move slower than wild type and with smaller amplitude and wavelength. Animals of three GABA transmission knock-out strains swam in saline with significantly lower translocation speed, undulation frequency, maximal amplitude, and primary wavelength than wild type. Calculated probabilities of the null hypotheses (p value) below 0.05 were considered significant and are in red. Download Figure 4-1, DOCX file.
Extended Data Figure 4-2
During free swimming, GABAergic mutants move at lower frequency and slower than wild type in both forward and backward directions. Same data as in Figure 4C presented in shared-control Gardner–Altman plots with raw data (upper axes) and mean differences (lower axes: bootstrap distributions, mean difference are dots; 95%CI are vertical error bars). Download Figure 4-2, TIF file.
Extended Data Figure 5-1
Optogenetic inactivation of all GABAergic neurons during free crawling and swimming reduces locomotion speed. When GABAergic motoneurons were acutely inactivated (animals fed with ATR and illuminated with lime-colored light), they crawled on agar surface or swam in saline with significantly lower translocation speed and undulation frequency. They were compared to two negative controls, the same animal under infrared light (ATR/infrared light) and the same strain not fed ATR (no ATR/infrared light). The only exception is that backward-crawling undulation frequency (but not translocation speed) of animals that were fed ATR were not statistically different under lime-colored or infrared light. Calculated probabilities of the null hypotheses (p value) below 0.05 were considered significant and are in red. Download Figure 5-1, DOCX file.
Extended Data Figure 5-2
Optogenetic inactivation of all GABAergic neurons during free crawling reduces locomotion speed and frequency. Transgenic animals fed with ATR crawl slower and less rapidly under the inactivation-inducing lime-colored light, than animals that were not fed ATR or are under infrared light. Same data as in the left plots of Figure 5D presented in shared-control Gardner–Altman plots with raw data (upper axes) and mean differences (lower axes: bootstrap distributions, mean difference are dots; 95%CI are vertical error bars). Download Figure 5-2, TIF file.
Extended Data Figure 5-3
Optogenetic inactivation of all GABAergic neurons during free swimming reduces locomotion speed and frequency. Transgenic animals fed with ATR crawl slower and less rapidly under the inactivation-inducing lime-colored light, than animals that were not fed ATR or are under infrared light. Same data as in the right plots of Figure 5D presented in shared-control Gardner–Altman plots with raw data (upper axes) and mean differences (lower axes: bootstrap distributions, mean difference are dots; 95%CI are vertical error bars). Download Figure 5-3, TIF file.
Extended Data Figure 8-1
Wild-type model parameters. Download Figure 8-1, DOCX file.
Extended Data Figure 8-2
Suppression of muscle cross-inhibition in computational models revealed differences between putative roles of inhibition in CPG and proprioceptive driven control of locomotion. Normalized undulation frequency (top) and locomotion speed (bottom) plotted as a function of muscle parameters representing (A) muscle activation (testing hypothesis 1) and (B) muscle waveform (testing hypothesis 2). See Figure 8 and text for details regarding hypotheses. Undulation frequency and locomotion speed are normalized by the model output at wild-type parameter values (black vertical dashed lines) in agar-like (red) and water-like (blue) environments. Horizontal dashed lines show frequencies and locomotion speeds in GABA defective nematodes (unc-25 experimental results) in agar (magenta) and water (cyan), normalized by the respective wild-type values. Experimentally, both frequency and speed are reduced in GABA-defective animals as compared to the wild type with a relatively larger reduction in water than on agar. A, Hypothesis 1. Muscle cross-inhibition enhances undulation frequency and locomotion speed. In the proprioceptive model, suppression of muscle cross-inhibition by approximately 8% and 30% quantitatively captures experimental drop in undulation frequency and speed on agar. Suppression of muscle cross-inhibition by the same amount in water results in a similar modulation of frequency. About 33% suppression of inhibition would be required to quantitatively account for the change in locomotion frequency in water, suggesting an additional role of inhibition. Model speed changes with reduced muscle cross-inhibition (
) are similar in CPG and proprioceptively controlled models. In the CPG-controlled model, the reduction in muscle amplitude leads to reduced locomotion speed, even at a fixed frequency, with a stronger speed reduction in water than in agar. B, Hypothesis 2. Muscle disinhibition increases the muscle response rate. In a proprioceptive control model (left), removal of disinhibition is modeled by an increased muscle time scale. Muscle time scales ( τ) of 0.12 and 0.25 s would account for the observed drop in frequency in water and agar, respectively; muscle time scales of 0.2 s would account for relative changes in model speeds. Under models of fixed-frequency CPG control (right), removal of disinhibition is modeled by a change of the waveform of muscle activation (from a near-square wave of the model wild type at
In this issue

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}



