

Article Figures & Data
Figures
- Figure 1.
Neuronal chloride accumulation during spontaneous ILDs and epileptogenesis in organotypic hippocampal slices in vitro. A, Microphotograph of incubated organotypic hippocampal slice from CLM1 mice captured at DIV14 and corresponding two-photon fluorescence image of the CA1 pyramidal cell layer. Overlays of 50 imaging planes from 0 to 100 μm below the slice surface (2-μm steps) are shown. Neuronal cell bodies are pseudo colored according to [Cl–]i (Dzhala et al., 2010, 2012). B, Simultaneous extracellular field potential recording and two-photon fluorescence chloride imaging were used to monitor neuronal network activity and [Cl–]i as a function of time. Example of spontaneous recurrent ILDs and corresponding [Cl–]i transients in the CA1 pyramidal cells at DIV14. C, Recurrent ILDs can last from 10 to 20 s to >5 min, the ILD duration threshold for SE. Example of recurrent ILDs and SE at DIV14. D, Percentage of slices with spontaneous recurrent ILDs and SE, and corresponding mean duration and frequency of ILDs as a function of age. E, [Cl–]i transients in the CA1 pyramidal cells during recurrent ILDs at DIV14. The sodium channel blocker TTX (1 μm) rapidly abolished recurrent ILDs and corresponding [Cl–]i transients, and significantly reduced the base line [Cl–]i. Exponential fits (y = A1exp(-x/t1) + y0) were used to measure base line [Cl–]i (y0) in individual cells (solid navy curves) during recovery from ILD (adj. R2 = 0.9–0.96). F, Box (left) + data (right) plots correspond to median (25−75%) [Cl–]i in paired cells (filled circles) and their distribution curves; open squares and whisker range indicate mean ± SD; ***p < 0.001 (paired sample Wilcoxon signed-ranks test). G, Median baseline [Cl–]i in individual slices as a function of age (DIV0–DIV28).
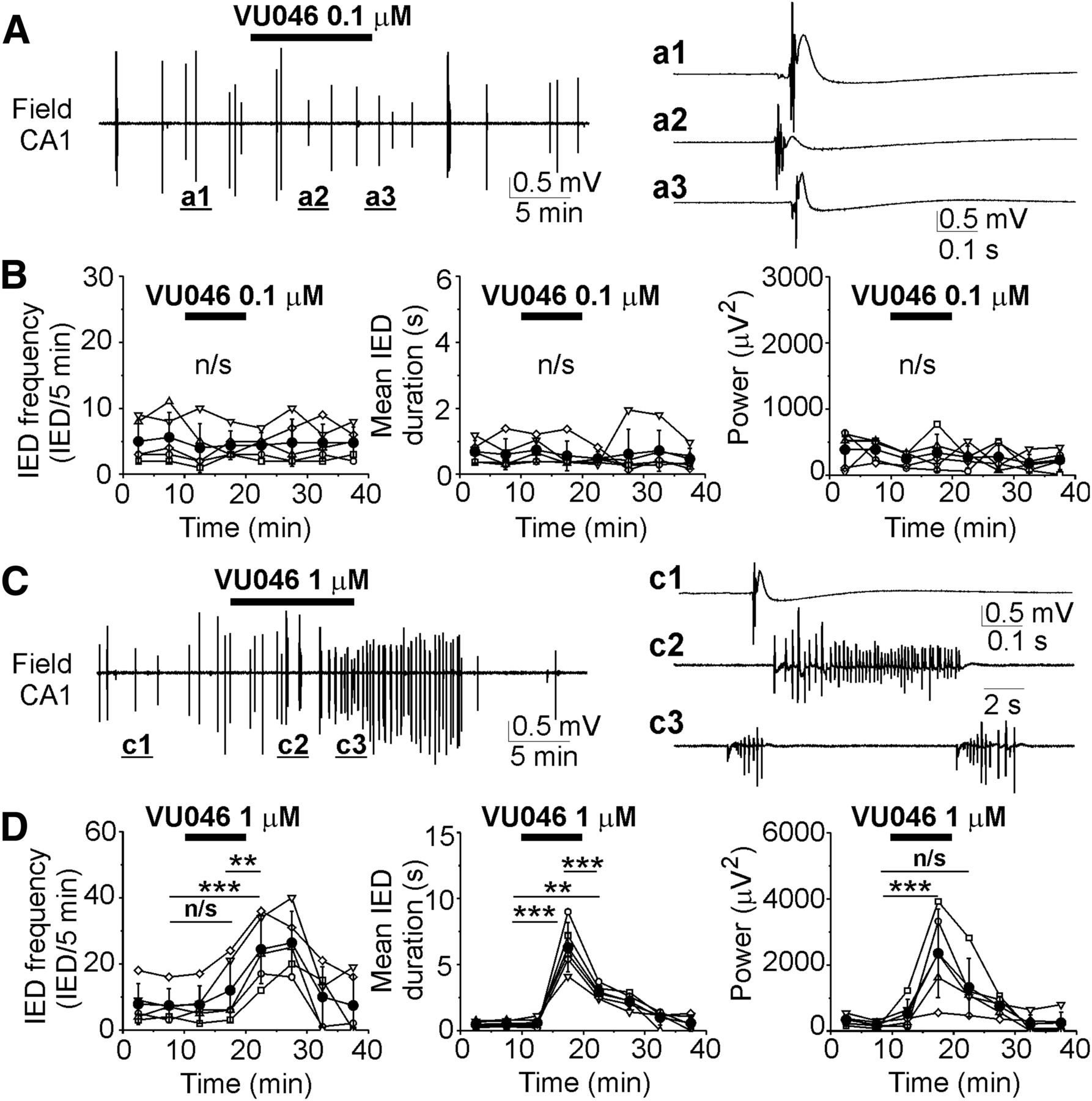
- Figure 2.
Effects of the KCC2 blocker VU0463271 on recurrent IEDs at DIV5–DIV7. A, C, Extracellular field potential recordings in the CA1 pyramidal cell layer in the organotypic hippocampal slices at DIV6 before (control), during and after application of 0.1 μm (A) and 1 μm (C) VU0463271 for 10 min. Expansion of IEDs in control and in the presence of VU0463271. B, D, Corresponding summary plots of the frequency and mean duration of recurrent IEDs in individual cultures (open symbols) and corresponding power of electrical activity in 5-min windows. Filled symbols indicate group mean ± SD. VU0463271 (1 μm) significantly increased the mean frequency, duration, and power of epileptiform discharges (n/s corresponds to p > 0.05; *p < 0.05; **p < 0.01; ***p < 0.001, one-way RM ANOVA, Tukey’s test).
- Figure 3.
Effects of the KCC2 blocker VU0463271 on recurrent ILDs and intracellular chloride at DIV14–DIV21. A, B, Extracellular field potential recordings in the CA1 pyramidal cell layer in the organotypic hippocampal slices at DIV18 before (control), during and after application of 0.1 μm (A) and 1 μm (B) VU0463271 for 30 min. Expansion of ILDs in control and in the presence of VU0463271. C–E, Corresponding summary plots of the frequency and mean duration of recurrent ILDs and corresponding power of electrical activity in 10-min windows in individual slice cultures (open symbols). Filled symbols indicate group mean ± SD. VU0463271 (1 μm) significantly reduced the mean frequency of ILDs and increased their duration and power (*corresponds to p < 0.05; ***p < 0.01, one-way RM ANOVA, Tukey’s test). F, Exponential fit (y = A1exp(–(x – x0)/t1) + y0) was used to measure base line [Cl–]i (y0) and decay time (t1) of base line [Cl–]i recovery in individual cells (solid navy curves) during ILD in control, during application of VU0463271 and TTX (adj. R2 = 0.9–0.96). F–H, VU0463271 (1 μm) increased base line [Cl–]i (y0) and delayed decay time of baseline chloride recovery (t1) during ILDs. G, TTX (1 μm) abolished ILDs and slowly reduced the median baseline [Cl–]i (*p < 0.05, Friedman RM ANOVA on ranks, Tukey’s test). H, Decay time of baseline [Cl–]i recovery during application of TTX in the presence of VU0463271 was significantly higher as in control (***p < 0.001, one-way RM ANOVA, Tukey’s test). I, Effects of VU0463271 in the presence of TTX on [Cl–]i. VU0463271 application in the presence of TTX did not significantly change the median baseline chloride concentration (p > 0.05, Wilcoxon signed-ranks test).
- Figure 4.
Effects of DMSO on recurrent ILDs at DIV12–DIhttV17. A, Extracellular field potential recording in the CA1 pyramidal cell layer in the organotypic hippocampal slices at DIV14 before (control), during and after application of DMSO (100 μl/100 ml). Expansion of ILDs in control (a1, a2), in the presence of DMSO (a3, a4), and during washout of DMSO (a5, a6). B, Corresponding summary plots of the frequency and mean duration of recurrent ILDs and corresponding power of electrical activity in 10-min windows in individual slice cultures (open symbols). Filled symbols indicate group mean ± SD (n/s corresponds to p > 0.05, one-way RM ANOVA).
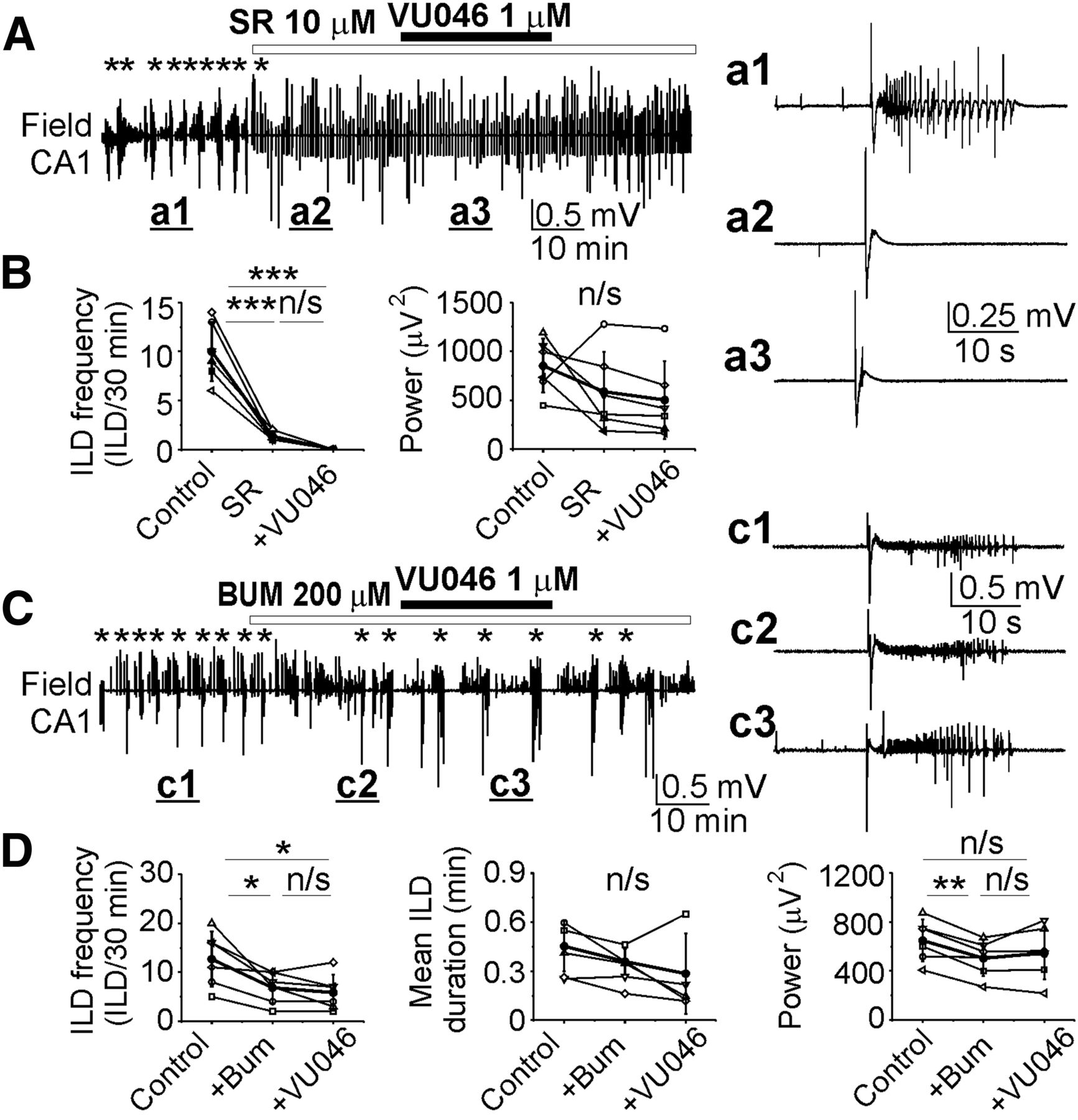
- Figure 5.
Selectivity of KCC2 blocker VU0463271. A, C, Extracellular field potential recordings in the CA1 pyramidal cell layer in the organotypic hippocampal slices in vitro. A, B, Application of the GABAA-R antagonist SR95531 (10 μm) abolished spontaneous ILDs and induced large amplitude IEDs. Subsequent application of VU0463271 (1 μm) in the presence of SR95531 did not change the mean frequency of ILDs discharges (n/s, p > 0.05; ***p < 0.001; one-way RM ANOVA, Tukey’s test) and power of electrical activity (p > 0.05; one-way RM ANOVA). A, a1–a3, Expansion of ILD in control (a1) and IEDs in the presence of SR95531 (a2) and VU0463271 (a3). C, D, Application of VU0463271 (1 μm) in the presence of NKCC1 and KCC2 blocker bumetanide (200 μm) did not change the mean frequency and duration of ILDs, and the mean power of electrical activity (p > 0.05, one-way RM ANOVA, Tukey’s test). c1–c3, Expansion of ILDs in control (c1), during application of bumetanide (c2), and in the presence of VU0463271 (c3).
- Figure 6.
Effects of NKCC1/KCC2 blocker furosemide on recurrent epileptiform discharges and [Cl–]i. A, B, Extracellular field potential recording in the CA1 pyramidal cell layer in the organotypic hippocampal slice at DIV22 before (control), during and after application of furosemide (0.1 and 1 mm for 30 min). C–E, Summary plots of the frequency of ILDs, IEDs, and power of corresponding activity in 10-min windows in individual slice cultures (open symbols). Filled symbols indicate mean ± SD. Furosemide (0.1 mm) reduced the frequency of ILDs and increased the frequency of IEDs. Furosemide (1 mm) abolished recurrent ILDs, increased the frequency of IEDs, and significantly decreased the power of electrical activity (*corresponds to p < 0.05; **p < 0.01, paired sample t test). F–H, Effects of furosemide (0.1 and 1 mm) on [Cl–]i. Furosemide (1 mm) abolished recurrent ILDs and corresponding [Cl–]i transients, and progressively reduced the baseline [Cl–]i. H, Baseline [Cl–]i changes in the presence of 0.1 mm furosemide and 1 mm furosemide as a function of an initial baseline [Cl–]i. I–K, Effects of TTX (1 μm) and furosemide (1 mm) in the presence of TTX on [Cl–]i. TTX rapidly abolished chloride transients, reduced the median baseline [Cl–]i (*p < 0.05 compared with control, Friedman RM ANOVA on ranks, Tukey’s test) and prevented effects of furosemide on [Cl–]i (p > 0.05 compared with TTX). L, Averaged mean [Cl–]i changes before and after applications of TTX (black) or furosemide (red). Data were fitted with exponential fit to calculate decay time of [Cl–]i extrusion during suppression of ILDs. Box (left) + data (right) plots correspond to median (25−75%) [Cl–]i in paired cells (filled circles) and their distribution curves; open squares and whisker range indicate mean ± SD. Furosemide significantly delayed chloride extrusion during suppression of ILDs (***p < 0.001; Mann–Whitney rank-sum test).
- Figure 7.
GABAA receptor-dependent anti-ictal effects of furosemide was enhanced in the presence of NKCC1 and KCC2 antagonists. A, C, E, Extracellular field potential recordings in the CA1 pyramidal cell layer in the organotypic hippocampal slices in vitro. Furosemide (1 mm) was applied in the presence of (A) GABAA receptor antagonist SR95531 (10 μm), (C) NKCC1 blocker bumetanide (10 μm), and (E) KCC2 blocker VU0240551 (10 μm). A, B, SR95531 abolished recurrent ILDs, induced hypersynchronous interictal bursts, and prevented the anti-ictal effect of furosemide. C, D, Furosemide potentiated anticonvulsant effect of bumetanide (10 μm). E, F, Furosemide in the presence of VU0240551 abolished ictal and IEDs. B, D, F, Corresponding summary data of the frequency of recurrent ILDs and power of electrical activity before and during drugs applications. Anti-ictal effects of furosemide were significant in the presence of bumetanide (D) and VU0240551 (F). N = 5–6 slices in each group of experiments; *p < 0.05, **p < 0.01, ***p < 0.001; one-way RM ANOVA, Tukey’s test.
- Figure 8.
The KCC2 enhancer CLP257 reduced duration of ILDs. A, B, Extracellular field potential recordings in the CA1 pyramidal cell layer before (control), during and after application of CLP257 (1 and 30 μm for 1 h). Examples of ictal-like events in control and during application of 1 μm CLP257 (A, a1, a2) and 30 μm CLP257 (B, b1, b2). C–E, Summary plots of the frequency and mean duration of ILDs, and power of electrical activity in individual slices (open symbols) and corresponding group mean ± SD (filled symbols) before, during and after 1 μm (top plots) and 30 μm (bottom plots) CLP257 applications. CLP257 (1 μm) significantly reduced the mean duration of ILDs (*p < 0.05, **p < 0.01, ***p < 0.001; paired sample t test). F, Effects of CLP257 on [Cl–]i as a function of time in n = 5 individual cell. G, H, Statistical histograms showing the effects of CLP257 (1 μm) on baseline [Cl–]i and decay time of chloride extrusion (*p < 0.05, Wilcoxon signed-rank test). CLP257 (1 μm) significantly accelerated extrusion rate (t1) of neuronal chloride during recurrent ILDs.







In this issue

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}



