

Article Figures & Data
Figures
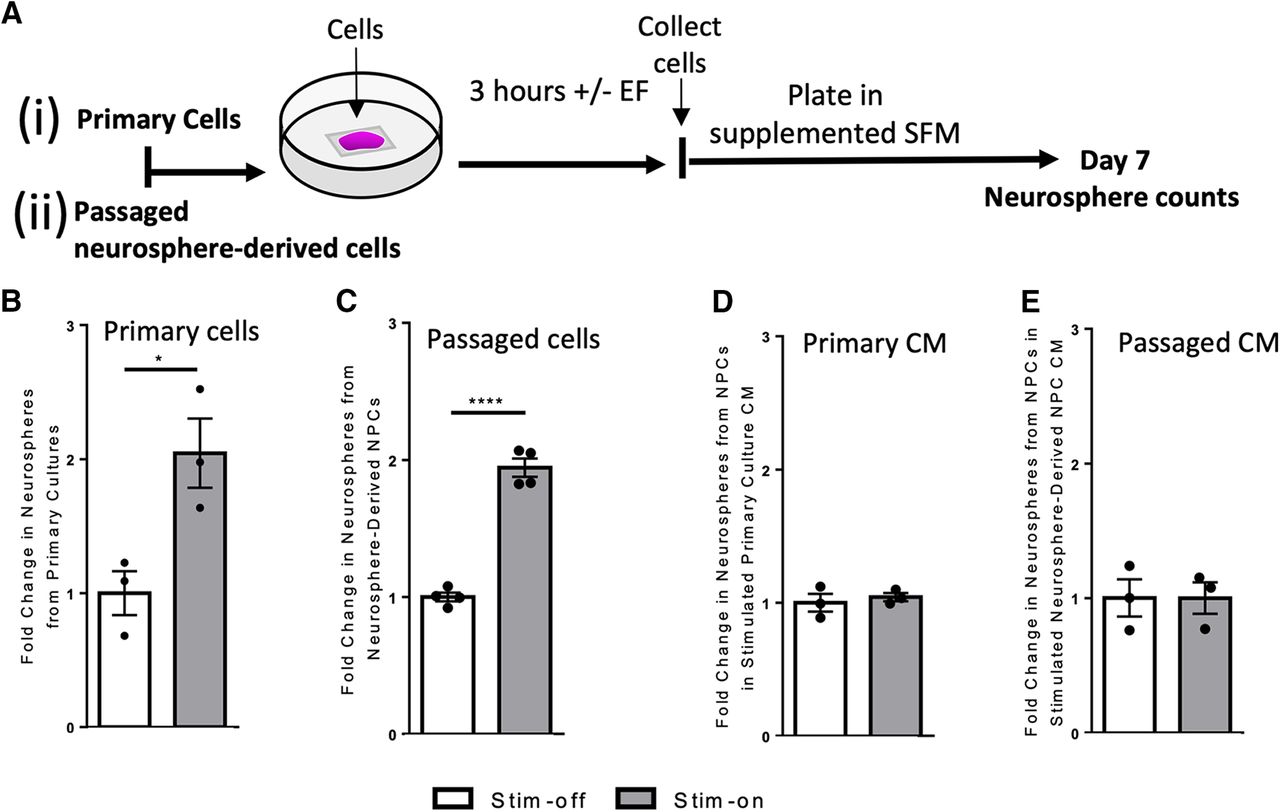
- Figure 1.
In vitro electrical stimulation increases the number of neurospheres. Ai, Aii, Experimental paradigms for in vitro electrical stimulation. B, C, Fold change in the number of neurospheres from stim-off cells and stim-on cells from (B) primary cultures (n = 3 independent experiments, *p = 0.03) and (C) neurosphere-derived NPCs (n = 4 independent experiments, ****p < 0.0001). Fold change in the number of neurospheres cultured using CM from stim-off and stim-on (D) primary cells (n = 3 independent experiments) and (E) neurosphere-derived NPCs (p = 0.5 and p = 0.9, respectively; n = 3 independent experiments) Each point in the graph represents an independent experiment, plotted with mean ± SEM. Analysis performed via a two-tailed unpaired t test between stim-off and stim-on groups. Neurosphere counts from in vitro stimulation found in Extended Data Figure 1-1. Comparison of fold changes in neurospheres following stimulation with and without Wnt inhibitor to inhibit symmetric divisions found in Extended Data Figure 1-2.
- Figure 2
In vitro electrical stimulation increases cell number and regulates NPC differentiation. A, A 1.7-fold increase in the number of living cells is seen following stim-on when compared with the stim-off cells processed the same way (62,400 ± 5705 vs 36,400 ± 3205 cells, stim-on vs stim-off respectively; n = 10 individual experiments; ***p = 0.0009). B, C, Fold change in the number of colonies from stim-off and stim-on in (B) primary cells (p = 0.06; n = 3 independent experiments) and (C) neurosphere-derived NPCs (p = 0.1; n = 4 independent experiments). Neurosphere counts from in vitro stimulation found in Extended Data Figure 1-1. D, Differentiated neurospheres stained for βIII tubulin (neurons), GFAP (astrocytes), and O4 (oligodendrocytes). White arrows indicate examples of cells co-localized with DAPI (blue). Scale bar = 100 µm. E–G, Quantification of cells as a percent of DAPI (n = 10 spheres/treatment, 3 mice per group) expressing (E) βIII tubulin (*p = 0.02), (F) GFAP (*p = 0.01), and (G) O4 (*p = 0.03). Each point in the graph represents an independent experiment, plotted with mean ± SEM. All analysis performed via a two-tailed unpaired t test between stim-off and stim-on.
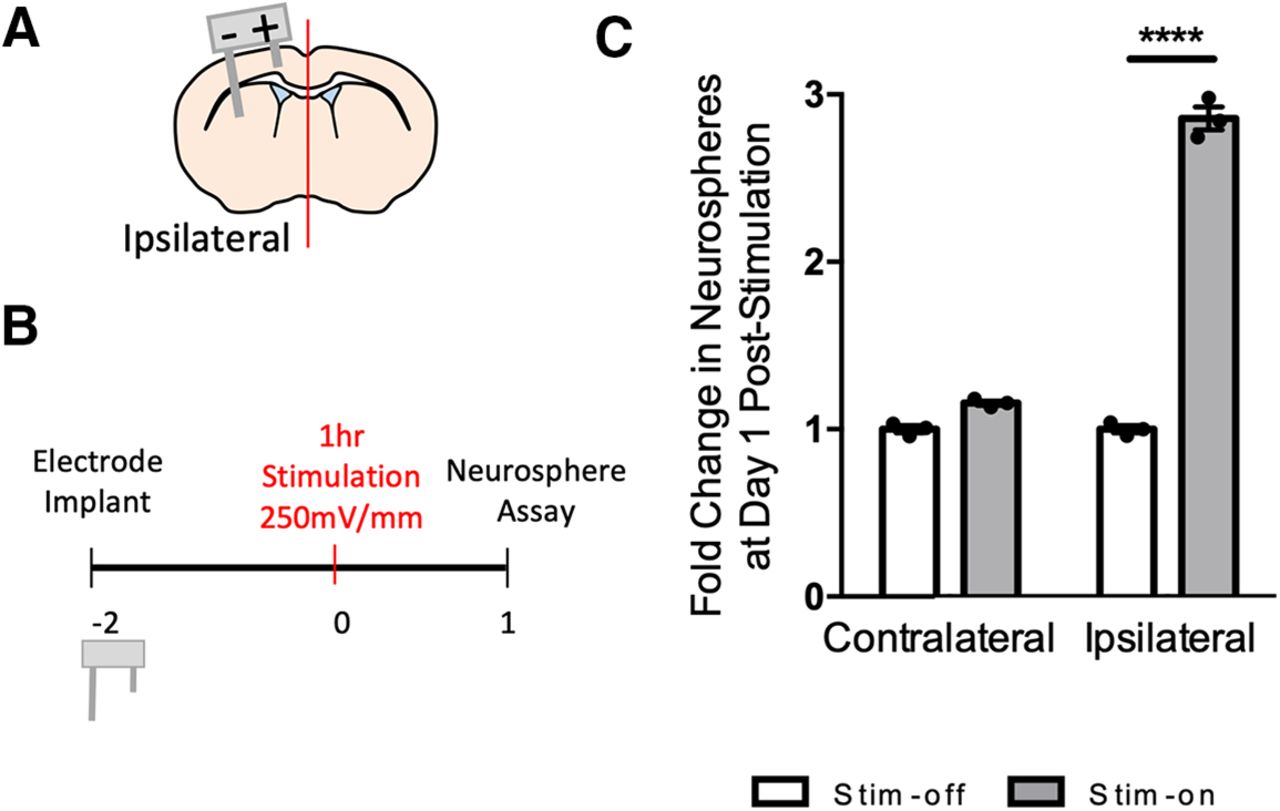
- Figure 3.
In vivo electrical stimulation increases the number of neurospheres and proliferation by a means other than activation. A, Image of 3D-printed electrode and schematic of a coronal section through the forebrain indicating the electrode implantation. Anode (+) and cathode (–) are shown. SVZ, subventricular zone. B, Experimental paradigm. C, D, Fold change in the number of neurospheres from stim-off and stim-on brains in the ipsilateral and contralateral hemisphere at (C) day 1 poststimulation (**p < 0.04) and (D) day 3 poststimulation [****p < 0.0001; n = 3 mice/group, two-way ANOVA (α = 0.05) with Tukey’s post hoc analysis]. There was no difference in fold change between days 1 and 3 as seen in Extended Data Figure 3-3. Neurosphere counts from in vivo stimulation found in Extended Data Figure 3-2. E–H, Images of the dorsolateral corner of the lateral ventricle ipsilateral to the electrode in 600 µm2 (EdU&DAPI) or 350 µm2 (EdU&Sox2, EdU&DCX, EdU&Iba1) and quantification. White arrows indicate cells positive for both labels in each image. Ctx, cortex; LV, lateral ventricle; CC, corpus callosum. White box indicates high power inset of dorsolateral corner (or corpus callosum for Iba1&Edu) in bottom left of image. Scale bar =100 µm (EdU&DAPI) or 50 µm. Quantification of cells as a percent of DAPI in the region of interest for stim-off and stim-on brains for (E) EdU+ cells (****p < 0.0001; n = 7 mice/group), (F) EdU+Sox2+ cells (*p = 0.03; n = 3 mice/group), (G) EdU+DCX+ cells (**p = 0.001; n = 3 mice/group), and (H) EdU+Iba1+ cells (p = 0.6; n = 3 mice/group). Analysis performed via two-tailed t test unpaired t test between stim-off and stim-on groups. I, Contour plots showing cell sorts from the stim-off and stim-on brains with the gating strategy as seen in Extended Data Figure 3-1. J, The relative percentage of activated and quiescent cells (n = 4 independent experiments, 5 mice per experiment). The green boxes in the contour plots represent GFAP::GFP+CD133+EGFRhigh cells, blue represents GFAP::GFP+CD133+EGFRlow cells, and red represents GFAP::GFP+CD133– cells (two-way ANOVA, α = 0.05 with Tukey’s post hoc multiple comparison test). Each point in the graphs represents an independent experiment, plotted with mean ± SEM. ns indicates there was no significance in quiescent to activated shift in either stim off or stim on group.
- Figure 4.
Striatal electrodes increase the number of neurospheres. A, Schematic of striatal electrodes in brain. Anode (+) and cathode (−). B, Experimental paradigm. C, Fold change in the number of neurospheres from stim-off and stim-on brains in the ipsilateral hemisphere following striatal stimulation (****p < 0.0001; n = 3 mice/group, two-tailed t test between each stim-off and stim-on groups). Each point in the graphs represents an independent experiment, plotted with mean ± SEM. Neurosphere counts from in vivo striatal stimulation are found in Extended Data Figure 4-1.



Extended Data
Extended Data Figure 1-1
Neurosphere counts from in vitro stimulation. Data reported in mean ± SEM; n = 3–4 mice per group. Download Figure 1-1, DOC file.
Extended Data Figure 3-1
Gating strategy for separating GFAP::GFP+CD133+EGFRhigh cells from GFAP::GFP+CD133+EGFRlow cells adapted from Codega et al. (2014). A, Cell debris was removed and NPCs were sorted using forward scatter (FSC-H) for cell size and side scatter (SSC-H) for cell granularity. B, Single, live cells were gated by removing doublets (C) and (D) DAPI– cells. E, GFP+ cells from the GFAP::GFP+ were selected and three populations were defined: GFAP::GFP+CD133– (red), GFAP::GFP+CD133+EGFRlow (blue) and GFAP::GFP+CD133+EGFRhigh (green). Age matched wild-type C57BL/6J mice were used for single color-stained cells and fluorescent minus one (FMO) gating optimization in which GFP was not required. Download Figure 3-1, TIF file.
Extended Data Figure 3-2
Neurosphere counts from in vivo cortical stimulation. Data reported in mean ± SEM; n = 3 mice per group. Download Figure 3-2, DOC file.
Extended Data Figure 3-3
Changes in the ipsilesional hemispheres from stim off and stim on. There was no difference in fold change between days 1 and 3; n = 3 mice per group, two-tailed unpaired t test, p = 0.1. Download Figure 3-3, TIF file.
Extended Data Figure 4-1
Neurosphere counts from in vivo striatal stimulation. Data reported in mean ± SEM; n = 3 mice per group. Download Figure 4-1, DOC file.
Extended Data Figure 1-2
Comparison of fold changes in neurospheres following stimulation with and without Wnt inhibitor (sFRP2). A, sFRP2 blocked the vast majority of neurosphere growth in the stim-off cultures as predicted with a 16-fold decrease of stem cells and (B) a 17-fold decrease in stim-on conditions (****p < 0.0001). C, Blocking symmetric division does not inhibit the increase in neurospheres following electrical stimulation; n = 3 independent experiments, two-tailed unpaired t test, p = 0.3. Download Figure 1-2, TIF file.
In this issue

{kind=link}
{kind=link}
{kind=link}
{kind=link}



