

Article Figures & Data
Figures
- Figure 1.
Nerve pathways between colon and the inferior mesenteric ganglion. A, Simplified schematic diagram showing the nerve connections between the colon, inferior mesenteric ganglion, and pelvic ganglia. While pelvic ganglia are always paired, the inferior mesenteric ganglion was either an unpaired single ganglion, or more commonly, two separate ganglia, each of which associated with one of the hypogastric nerves. B, Schematic diagram showing a colonic tube preparation with connections to the pelvic ganglia and inferior mesenteric ganglion as typically arranged in an organ bath for recording. Note the diagram shows the most common arrangement featuring a paired IMG with a single connecting hypogastric nerve trunk. For simplicity, the other IMG is not shown. The centrifugal processes of cut lumbar colonic nerves, which could be seen emerging from the IMG, were isolated in paraffin oil for sympathetic nerve recordings, as shown. Conversely, rectal nerve recordings (of viscerofugal neuron activity) were made from the peripheral sides of cut nerves. Smooth muscle electrical activity was recorded by suction electrode within ∼5 mm of recorded rectal nerve entry to the gut wall (15–20 mm from the terminal rectum). IMG, inferior mesenteric ganglion; LCN, lumbar colonic nerves; PG, pelvic ganglia; HGN, hypogastric nerve; RN, rectal nerve; IMN, intermesenteric nerve; IMA, inferior mesenteric artery.
- Figure 2.
Coordinated burst firing in rectal nerves during motor complexes. A, Colonic EMG and rectal nerve recording traces showing ∼15 min of ongoing motor complexes and firing activity. Four motor complexes can be seen in the EMG trace. B, A single motor complex corresponding to the event marked by an asterisk below nerve 3 voltage trace in A. At this timescale, oscillations in the EMG trace and bursts of firing in the nerve recording traces are evident. There are also several larger amplitude spikes in nerve recording traces 1 and 2 that do not participate in the burst firing activity. C, A further expanded view of the motor complex shown in B. Here, the bursts of firing are clearly seen. Furthermore, burst firing appears coordinated between each of the three rectal nerves and EMG oscillations.
- Figure 3.
Drug responses in fresh preparations. A, Example of the prompt abolition of motor complexes and large reduction in rectal nerve firing by nicotinic receptor antagonist, hexamethonium. Neither burst firing, nor motor complexes occurred in hexamethonium (n = 7). B, Example of capsaicin-evoked firing in a fresh preparation, consistent with the presence of spinal afferent neurons.
- Figure 4.
Coordinated burst firing in rectal nerves during motor complexes after organ culture. A, Colonic EMG and rectal nerve recording showing ∼15 min of ongoing activity in an organ cultured preparation, including two motor complexes. The firing rates and spike shapes of four single units discriminated from the rectal nerve recording are shown. Most single units showed an increase in firing rate during motor complexes. B, A single motor complex, revealing burst firing behavior in the rectal nerve. Several units contributed to the burst firing behavior; their individual spike events are indicated below the nerve recording trace. C, The same event shown in B with an expanded timescale, showing in more detail the individual contributions to burst firing made by four single units. D, An individual burst corresponding to the event marked by an asterisk in C.
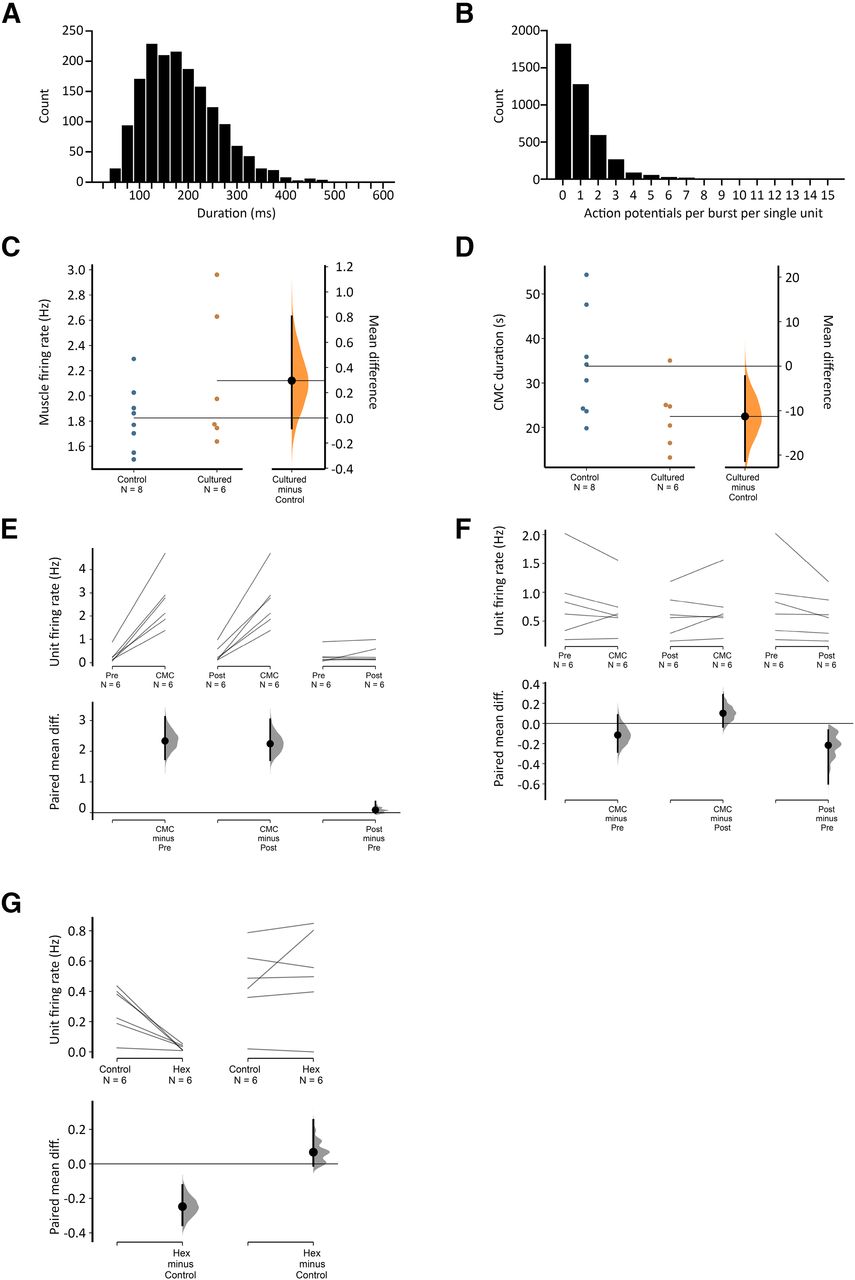
- Figure 5.
Viscerofugal neuron burst firing characteristics. A, The duration of 1676 individual bursts of firing in rectal nerves during 31 motor complexes in six organ cultured preparations shown as a frequency distribution. B, Distribution of the average numbers of action potentials each unit contributed to each burst in rectal nerves. Single units did not contribute to every instance of burst firing in rectal nerves, as may be seen in Figure 4. This highlights the importance of synchronization among multiple neurons, since single units alone were unlikely to encode the entire burst firing pattern. Single units contributed between 0 and 12 action potentials to a burst of firing. C, D, Comparisons of CMC smooth muscle firing rate and duration in control and cultured preparations, showing no significant differences between the two groups (p = 0.200 and 0.069, respectively, independent samples t test). Estimated mean differences between the groups are shown in a separate graph to the right. E, F, Average single unit firing rates before, during and after CMCs among units that contributed to burst firing behavior (E) and those that did not (F). All data are paired, and mean differences are shown graphically below mean firing rates. Firing in single units increased by 1–3 Hz among those that contributed to burst firing during the CMC compared with before or after CMCs, but not among those that did not contribute to burst firing (F). See text for details. G, The effect of hexamethonium on ongoing firing rate was significant among contributing but not non-contributing burst firing units.
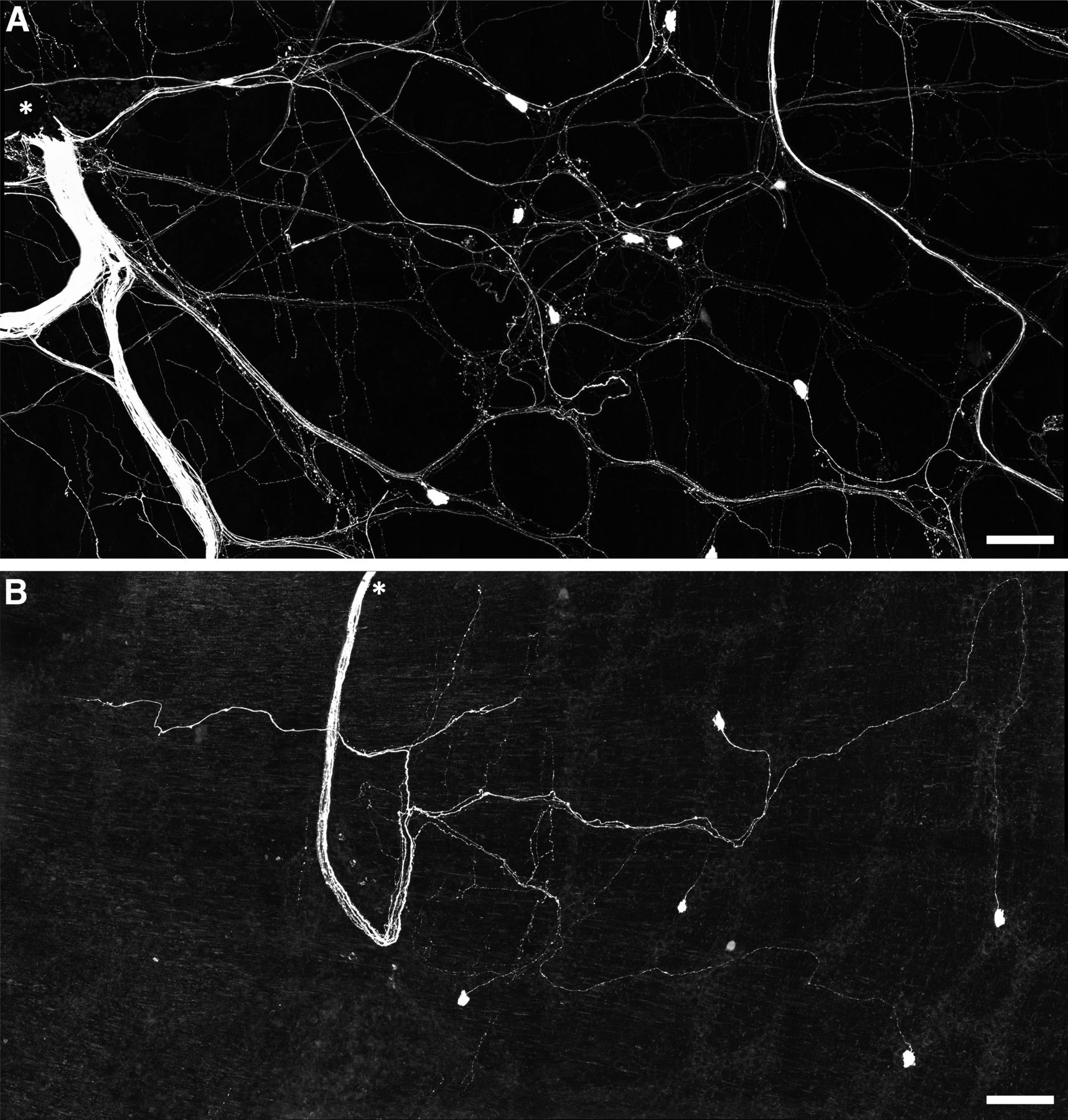
- Figure 6.
Persistence of viscerofugal nerve cell bodies in organ cultured preparations. A, Confocal micrograph showing biotinamide neuronal tracing from a rectal nerve trunk in a fresh preparation. The labeled nerve trunk is indicated by an asterisk. Numerous large axons and smaller varicose fibers can be seen coursing throughout the myenteric plexus in all directions. Viscerofugal nerve cell bodies were also labeled. B, Biotinamide neuronal tracing from rectal nerves after organ culture also revealed viscerofugal nerve cell bodies but a substantially reduced density of fine varicose fibers. Calibration, 100 μm.
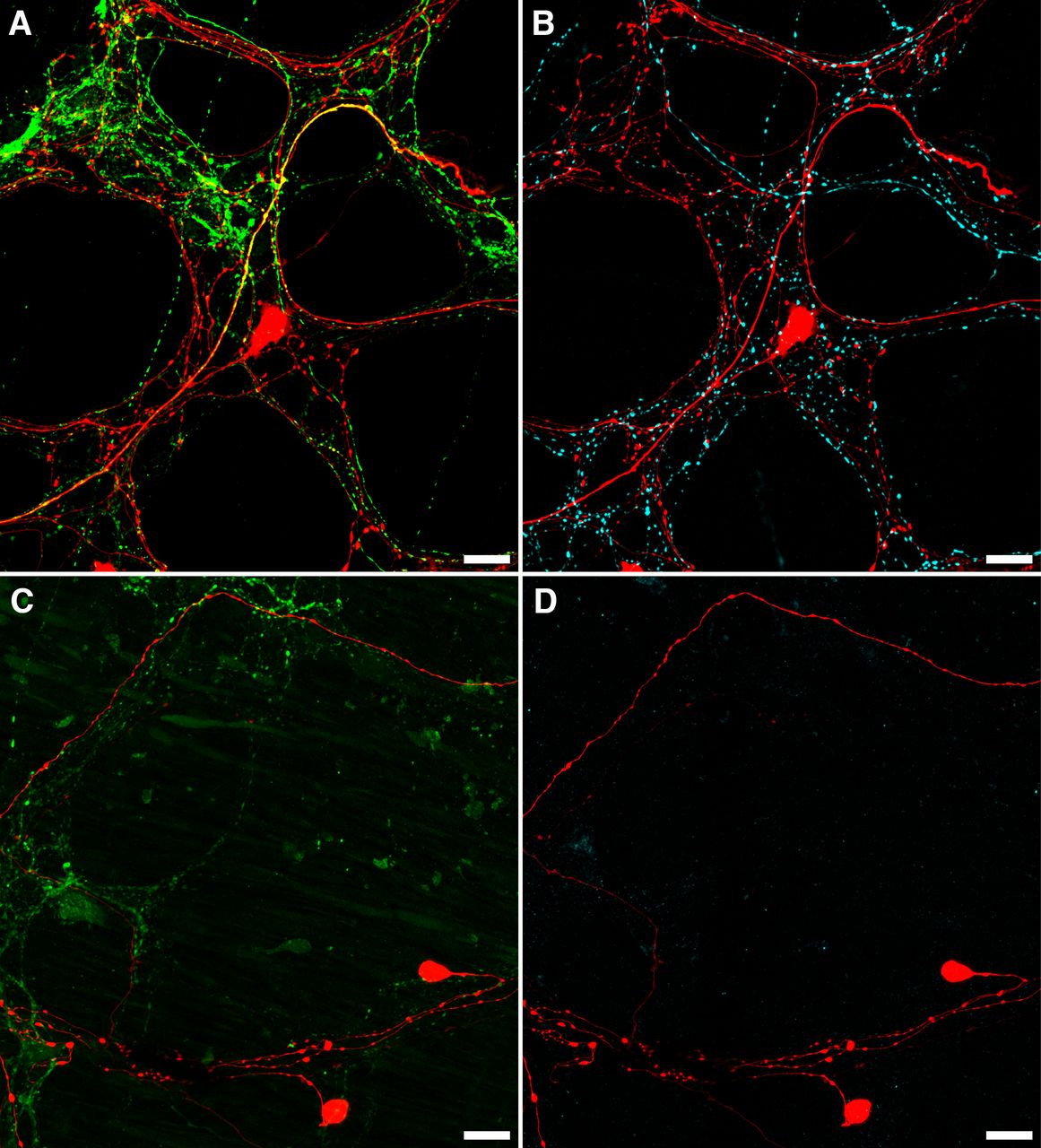
- Figure 7.
Loss of CGRP and TH after organ culture. A, B, An example of CGRP (A; green) and TH (B; cyan) immunofluorescence in the same biotinamide labeled (red) control preparation. Numerous varicosities containing TH or CGRP can be seen within the myenteric plexus, with some co-labeling of biotinamide-labeled axons and varicosities apparent with CGRP (yellow; A), but not TH (n = 3). This is consistent with the presence of spinal afferent axons in control preparations. In organ cultured preparations, immunohistochemically detectable CGRP (C; green) and TH (D; cyan) were dramatically reduced, showing degeneration of extrinsic nerve fibers, while viscerofugal nerve cell bodies persisted (biotinamide; red). Additionally, no co-labeling of these markers occurred with biotinamide-labeled axons. Expectedly, faintly CGRP-immunoreactive varicosities and nerve fibers persisted in organ cultured preparations. This is consistent with a population of intrinsic enteric neurons. Together, these data support a viscerofugal origin of the activity recorded from rectal nerve trunks in organ cultured preparations.
- Figure 8.
Sympathetic neuron firing and motor complexes. A, Colonic EMG and lumbar colonic nerve recordings (central efferent side) showing ongoing motor complexes over ∼10 min. Sympathetic firing increased during each motor complex. B, A single motor complex showing sympathetic burst firing at close to 2 Hz and single unit spikes.
- Figure 9.
Simultaneous afferent/efferent nerve recording and the abolition of lumbar colonic nerve efferent firing by hypogastric nerve crush. A, Example of three motor complexes and associated firing in two efferent recordings (lumbar colonic nerves) and a single afferent recording (rectal nerve). B, The second motor complex shown in A on an expanded timescale. Note the coordination of efferent firing between the two lumbar colonic nerves. Some burst firing activity in the rectal nerve (afferent) can be seen among dense ongoing firing. C, Part of the same event shown in B with an expanded timescale. At this scale both afferent (rectal nerve) and efferent (lumbar colonic nerves) burst firing can be visualized. D, Abolition of efferent nerve firing by hypogastric nerve crush. Motor complexes persisted following hypogastric nerve crush, without associated bursts of sympathetic neuron firing.







In this issue

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}



