

Article Figures & Data
Figures
- Figure 1.
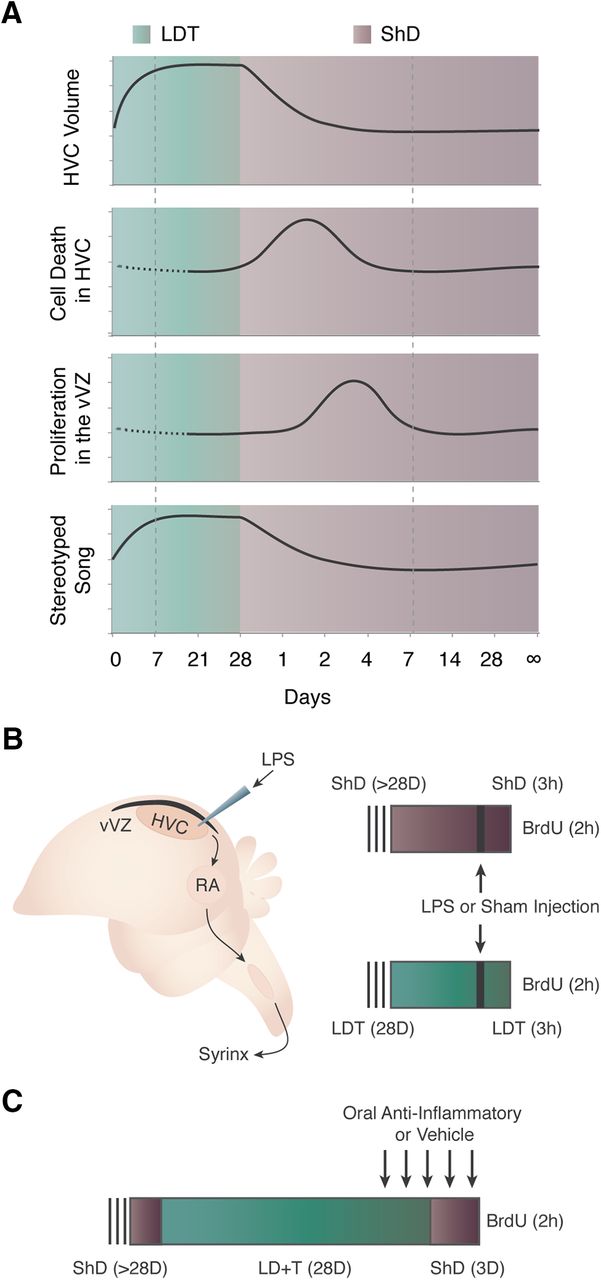
Experimental background and design. A, Seasonal growth and regression of HVC and effects on song stereotypy are illustrated. As white-crowned sparrows transition from breeding (green background) to nonbreeding conditions (purple background), HVC regresses in volume due to cell death. Increased proliferation of NPCs in the vVZ (black strip dorsal to HVC) that gives rise to new HVC neurons follows temporally and is induced by HVC cell death (Larson et al., 2014). Morphology and physiology reaches homeostasis in both breeding and nonbreeding conditions (indicated by dotted box). Dotted line indicates a lack of published data for those time points. B, Schematics illustrating the song production circuit and experimental design for testing the role of acute, local inflammation on NPC proliferation. The song production circuit consists of HVC (proper name), the RA, and nXIIts, nucleus para-ambiguus. Birds in nonbreeding (ShD; purple) or breeding (LDT; green) condition for 28 d received LPS through needle microinjection into HVC, while controls received sham injections. To label proliferating NPC, BrdU was administered systemically 1 h following LPS or sham injection, and 2 h before tissue harvesting. C, Experimental design to test the necessity of inflammation for reactive neurogenesis following natural cell death in HVC. Birds received oral doses of the anti-inflammatory agent minocycline starting 2 d before and continuing throughout the 3 d of HVC regression after rapid transition of birds from LDT to ShD conditions. Control birds received oral doses of water. All birds were injected with BrdU to label proliferating NPC 2 h before tissue harvesting.
- Figure 2.
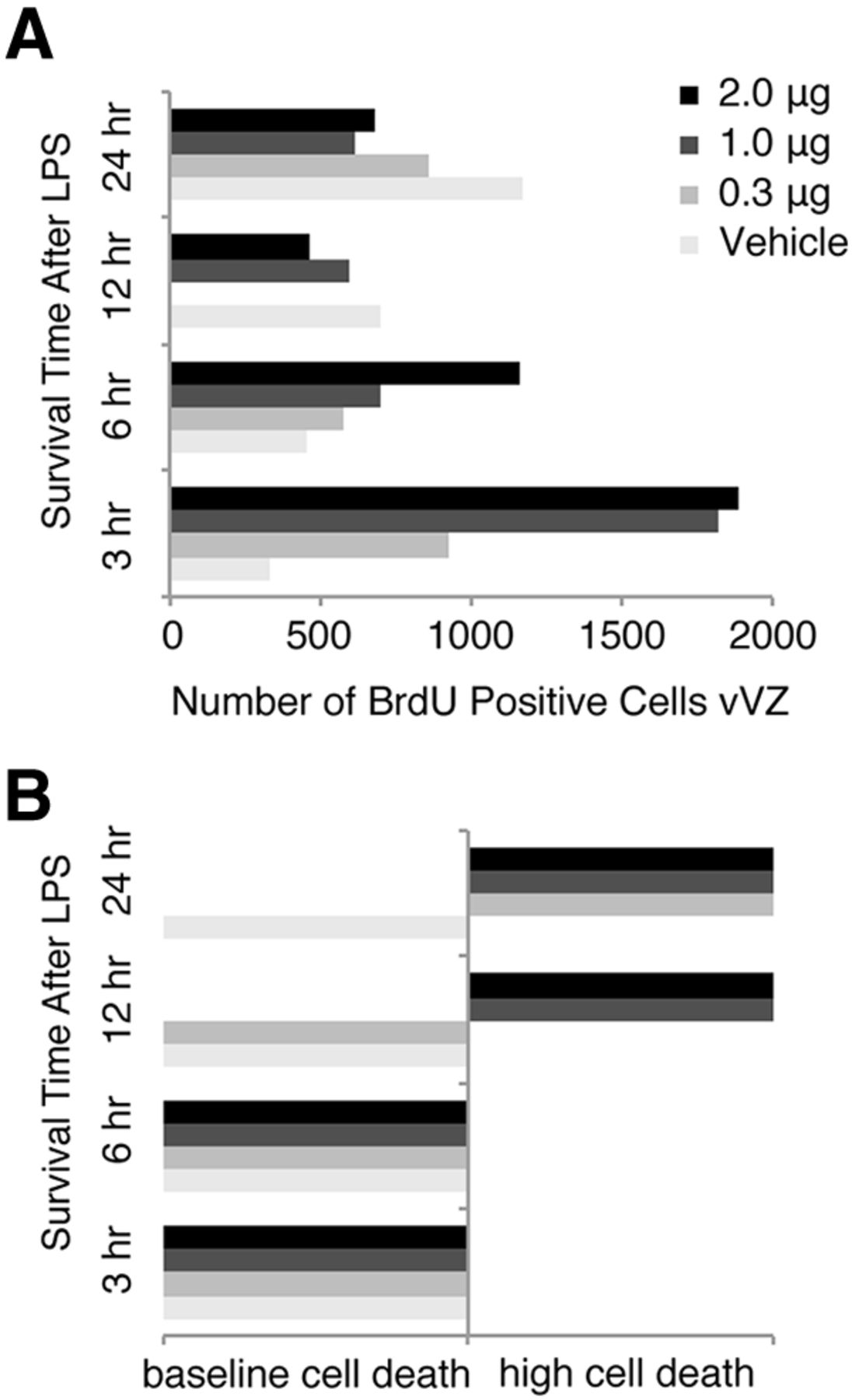
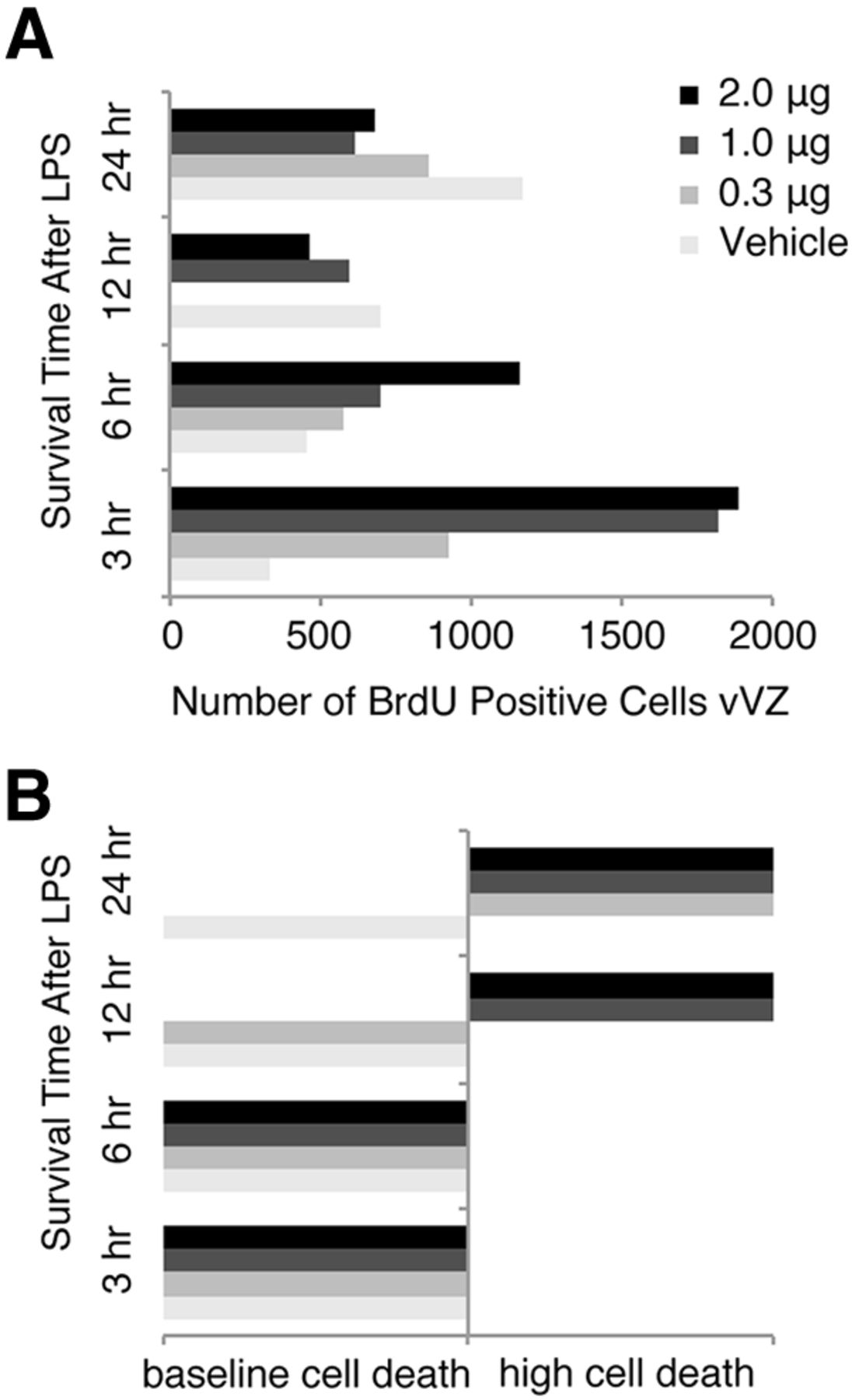
Identification of an LPS injection dose and pulse time that increases vVZ NPC proliferation but does not increase HVC cell death. A, BrdU-positive cells in unilateral vVZ following injection of LPS ranging from 0 to 2 μg and a survival time of 3–24 h before tissue harvesting. NPC proliferation appeared to increase as soon as 3 h following LPS injection for all groups except saline vehicle. Saline appeared to increase the number of BrdU-positive vVZ cells by 24 h following HVC injection. Each experimental and control group, n = 2. B, Grossly observed cell death in HVC following injection of LPS or vehicle over the time course. LPS appeared to induce cell death as soon as 12 h following LPS injection in a dose-dependent manner. By 24 h, all doses of LPS induced HVC cell death obviously visible by large numbers of Nissl-stained pyknotic cells. LPS appeared to not induce large quantities of HVC cell death with 3 and 6 h survival times. Because a dose of 1 μg of LPS induced a large increase in vVZ proliferation but no cell death after 3 h, we chose this dose and time course for our experiment.
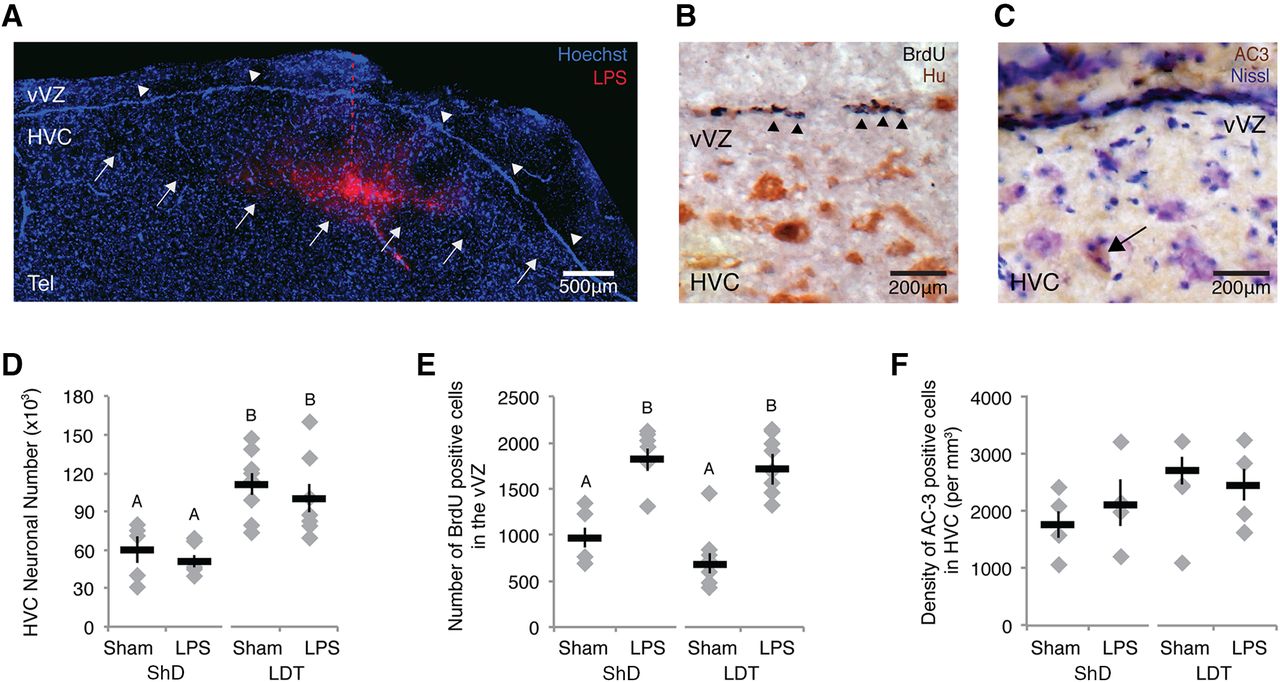
- Figure 3.
Local, acute inflammation induced a rapid increase in NPC proliferation independently of physiological condition (i.e., breeding or nonbreeding) and cell death. A, Image of Alexa Fluor-conjugated LPS localization within HVC 3 h after microinjection. LPS is shown in red; Hoechst 33342, a nuclear stain, is shown in blue. White arrows indicate the ventral boarder of HVC, white arrowheads indicate the vVZ, and the red dotted line indicates the injection needle track. Tel, Telencephalon. B, BrdU labeling (dark purple, indicated by black arrowheads) in the vVZ, just dorsal to HVC. Tissue was colabeled with an antibody against the neuronal antigen HuC/D to provide histologic landmarks during analyses. C, Cell death within HVC as indicated by AC-3-positive immunolabeling in brown, as indicated by the black arrow. Tissue is Nissl counterstained to identify the borders of HVC. D, Number of all HuC/D-positive neurons (i.e., new plus mature) in one hemisphere of HVC across experimental and control groups. The HVC neuron number was consistently lower in nonbreeding than breeding conditions. LPS did not affect the total neuronal number in breeding or nonbreeding condition birds. E, Number of BrdU-positive cells in the vVZ of experimental and control groups in breeding and nonbreeding conditions. LPS increased the numbers of new cells in vVZ equally in both breeding and nonbreeding condition birds. F, The density of AC-3-positive cells did not differ between LPS-injected and control birds, or between long-term breeding and long-term nonbreeding conditions. E, F, Different letters above error bars indicate significant differences among groups, as determined with post hoc Tukey’s test analyses (Table 1, ANOVA effects). All data are plotted as the mean ± SEM (black), with data for each individual bird displayed as gray diamonds.
- Figure 4.
Representative images of microglia with different morphologies observed within HVC. A–D, Microglia are stained with RCA-I (black), and neurons are immunolabeled for neuronal antigen HuC/D (brown). Nuclei are counterstained with methyl green (green). A, Three morphologies of microglia quantified in HVC (within dotted box). Ramified microglia (arrowhead) in close proximity to the vVZ (arrows; farthest left image). Active microglia including intermediate microglia (arrowhead; middle image) and reactive microglia (arrowhead; farthest right image) with commingling with neurons (arrows). B, Ramified microglia (arrowhead) in HVC contacting the vasculature (arrow). C, Ramified microglia (arrowhead) with processes intermingling with neuronal soma (arrows). D, Microglia with processes in contact with neuronal soma (arrow) and a non-neuronal cell (arrowhead).
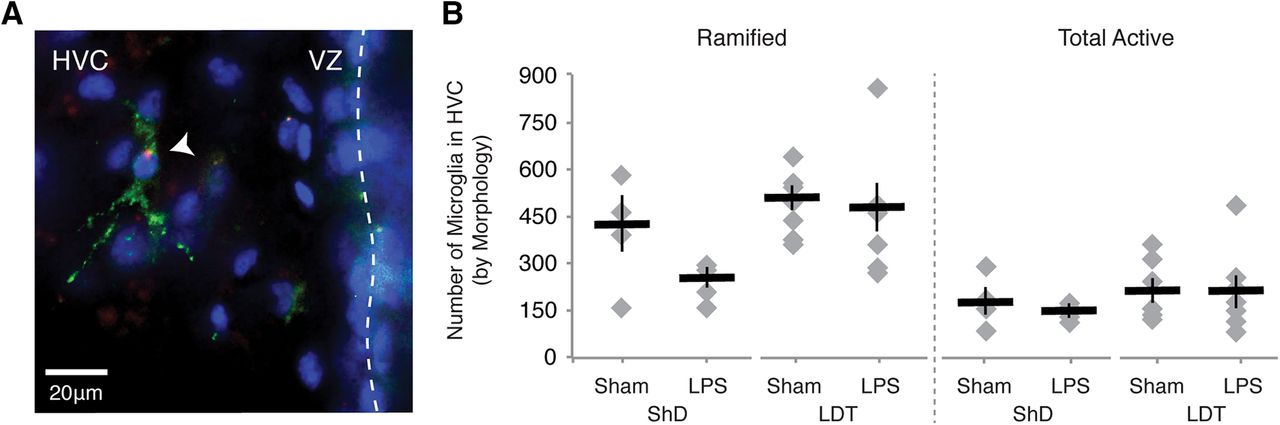
- Figure 5.
Microglial response to microinjection LPS into HVC. A, A microglial cell labeled with RCA-I (green) in HVC near the vVZ (dotted line). The presence of fluorophore-conjugated LPS (red) puncta within the cell body of the microglia is suggestive of endocytosis of the LPS by microglia. Cell nuclei stained with Hoechst 33342 are shown in blue. B, Number of RCA-I-labeled microglia as distinguished by morphology in HVC of one hemisphere (see also Table 1).
- Figure 6.
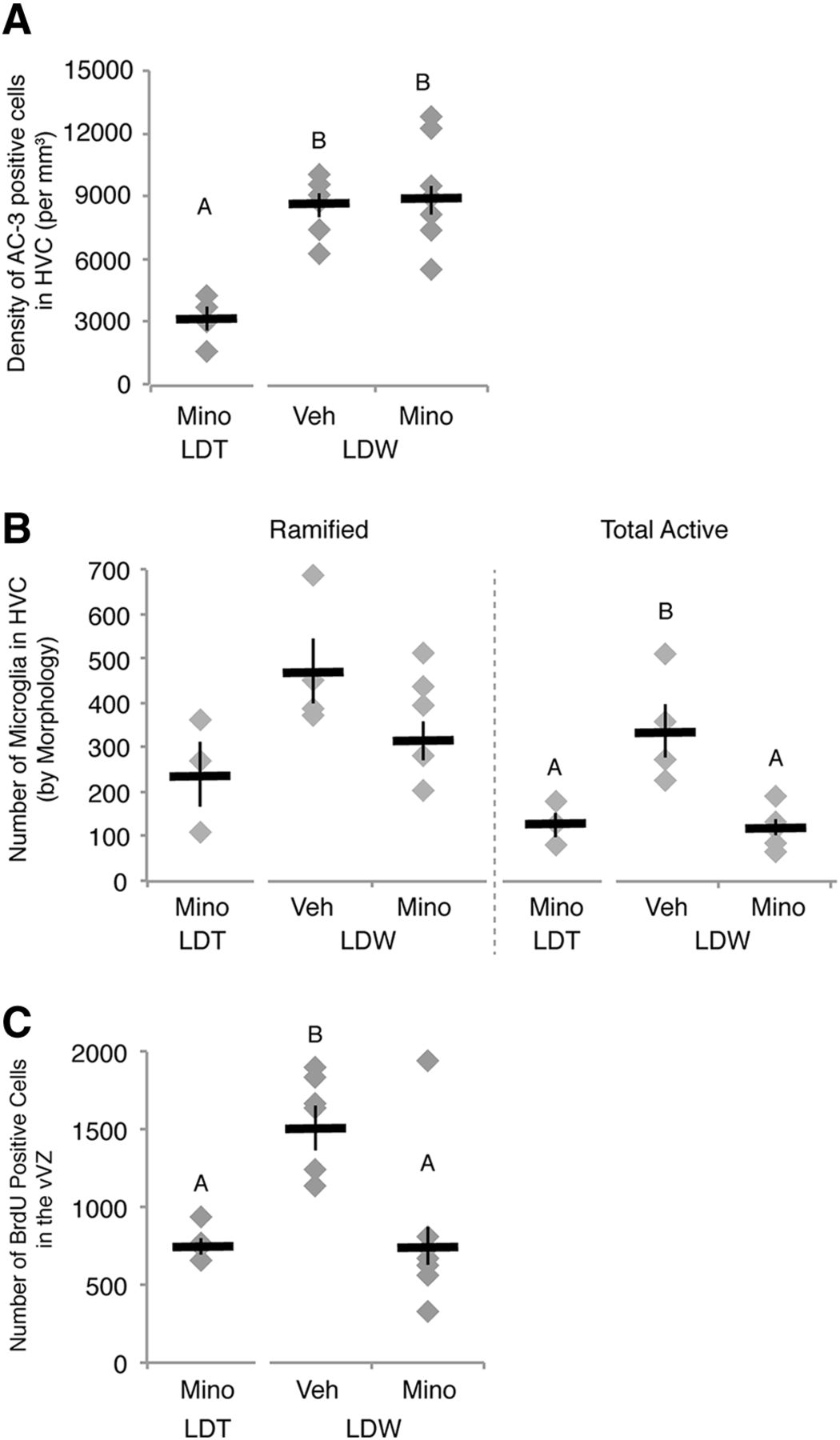
Systemic administration of minocycline (Mino) prevented natural, reactive NPC proliferation following HVC neuronal loss. A, Density of apoptotic cells in HVC. The number of dying cells in HVC as shown by AC-3-positive labeling increased in birds transitioned from breeding to nonbreeding conditions and was not altered by minocycline administration. B, Number of RCA-I-labeled microglia as distinguished by morphology in HVC of one hemisphere (see also Fig. 4, Table 2). The combined number of microglia with activated morphology (intermediate and reactive) was upregulated during regression [i.e., LDW with vehicle (Veh) administration (LDW-V)] when compared with breeding condition birds [i.e., LDT with vehicle (LDT-V)]. Minocycline reduced the activation of microglia during periods of neuronal loss in HVC in LDW birds (i.e., LDW-M). C, The number of BrdU-positive cells in the vVZ was greater in vehicle control birds transitioned from LDT to ShD, as previously reported (Larson et al., 2014), than birds maintained in LDT. Administration of minocycline prevented reactive NPC proliferation in the vVZ during active HVC regression, suggesting that inflammation is necessary for reactive neurogenesis. A–C, Letters above bars indicate significant differences among groups (ANOVA, post hoc Tukey’s test). Different letters above error bars indicate significant differences among groups as determined with post hoc Tukey’s test analyses (but see Table 2 for ANOVA effects). All data are plotted as the mean ± SEM (black), with individual data values displayed as gray diamonds.



Tables
- Table 1
Sex steroid levels, morphometrics, and cellular changes with induction of inflammation
ShD LDT ANOVA main effect(two-way) Injection type(Sham vs LPS) Condition (ShD vs LDT) Injection * Condition Sham
(n = 5)LPS
(n = 6)Sham
(n = 9)LPS
(n = 9)Testosterone Plasma levels (ng/ml) 0.07 ± 0.03a 0.08 ± 0.02a 15.80 ± 1.69b 17.70 ± 1.65b F(3,20) = 31.8914; p < 0.0001 F = 0.2986; p = 0.5908 F = 990.7318; p < 0.0001 F = 0.2920; p = 0.5949 HVC Unilateralvolume (mm3) 0.42 ± 0.05a 0.41 ± 0.04a 0.80 ± 0.05b 0.75 ± 0.06b F(3,24) = 10.6481; p < 0.0001 F = 0.3873; p = 0.5396 F = 30.5611; p < 0.0001 F = 0.2033; p < 0.0001 Neuron density(×103/mm3) 130 ± 5a,b 117 ± 4a 129 ± 3b 128 ± 4b F(3,25) = 6.8570; p = 0.0016 F = 1.8998; p = 0.1803 F = 16.3837; p = 0.0004 F = 1.9168; p = 0.1784 Unilateral neuronnumber (×103) 60 ± 10a 51 ± 5a 111 ± 9b 100 ± 11b F(3,24) = 5.3377; p = 0.0058 F = 0.0773; p = 0.7834 F = 15.1049; p = 0.0007 F = 0.5399; p = 0.4696 AC-3 cell density(per mm3) 1747 ± 230 2132 ± 411 2696 ± 241 2445 ± 271 F(3,21) = 2.0696; p = 0.1349 F = 0.0432; p = 0.8373 F = 4.4773; p = 0.0465 F = 1.1176; p = 0.3025 Unilateral AC-3 cell number 829 ± 200a 946 ± 245a 2328 ± 292b 2060 ± 360a,b F(3,21) = 5.9919; p = 0.0041 F = 0.1334; p = 0.7186 F = 16.1711; p = 0.0006 F = 0.4803; p = 0.4803 Microglia in HVC Ramified 295 ± 86 231 ± 29 479 ± 38 451 ± 73 F(3,18) = 2.6917; p = 0.0770 F = 2.1850; p = 0.1566 F = 5.4407; p = 0.0315 F = 1.0983; p = 0.3085 Intermediate 85 ± 30 128 ± 11 142 ± 23 139 ± 40 F(3,18) = 0.5415; p = 0.660 F = 0.3760; p = 0.5474 F = 1.1190; p = 0.3041 F = 0.2635; p = 0.6139 Reactive 32 ± 12 43 ± 7 48 ± 10 51 ± 11 F(3,18) = 0.4788; p = 0.7010 F = 0.4027; p = 0.5337 F = 1.0125; p = 0.3276 F = 0.0982; p = 0.7576 Total active 129 ± 41 156 ± 12 190 ± 33 190 ± 49 F(3,18) = 0.4510; p = 0.7197 F = 0.0976; p = 0.7583 F = 1.1961; p = 0.2885 F = 0.1018; p = 0.7533 Total all 552 ± 108 361 ± 41 668 ± 57 641 ± 93 F(3,18) = 2.5088; p = 0.0915 F = 1.6603; p = 0.2139 F = 5.4621; p = 0.0312 F = 0.9271; p = 0.3484 vVZ Unilateral BrdU-positive cel number 971 ± 164a 1819 ± 123b 690 ± 113a 1712 ± 106b F(3,22) = 22.0745; p
< 0.0001F = 54.4770; p < 0.0001 F = 2.394; p = 0.1412 F = 0.4709; p = 0.4997 Age Minimum age(month) 21 ± 2 19 ± 3 20 ± 2 17 ± 2 F(3,25) = 0.3227; p = 0.8089 F = 0.5843, p = 0.4518 F = 0.2273, p = 0.6377 F = 0.0493, p = 0.8262 All values are the mean ± SEM. Superscript letters denote significant differences across treatment groups with post hoc Tukey’s test, only if the main effect of ANOVA was significant.
- Table 2
Sex steroid levels, morphometrics, and cellular changes with inhibition of inflammation
LDT LDW ANOVA main effect
(one-way)Minocycline
(n = 4)Vehicle
(n = 7)Minocycline
(n = 10)Testosterone Plasma levels (ng/ml) 10.64 ± 3.15a 0.45 ± 0.07a 0.85 ± 0.18a F(2,16) = 20.7571; p < 0.0001 HVC Unilateral volume (mm3) 0.71 ± 0.03a 0.58 ± 0.01a,b 0.47 ± 0.04b F(2,17) = 12.8758; p = 0.0004 Neuron density (×103/mm3) 133 ± 3 139 ± 3 139 ± 3 F(2,16) = 0.9627; p = 0.4029 Unilateral neuron number (×103) 94 ± 3a 80 ± 2b 61 ± 3c F(2,16) = 29.6698; p < 0.0001 AC-3 cell density (per mm3) 3188 ± 573a 8600 ± 561b 8941 ± 696b F(2,17) = 15.1670; p = 0.0002 Unilateral AC-3 cell number 2222 ± 538a 5033 ± 380b 4108 ± 153b F(2,16) = 20.3012; p < 0.0001 Microglia in HVC Ramified 237 ± 72 463 ± 72 355 ± 55 F(2,9) = 2.5743; p = 0.1306 Intermediate 63 ± 27a 218 ± 26b 71 ± 12a F(2,9) = 17.4903; p = 0.0008 Reactive 61 ± 4 108 ± 55 40 ± 20 F(2,9) = 1.0589; p = 0.3864 Total active 124 ± 27a 326 ± 60b 111 ± 21a F(2,9) = 9.6118; p = 0.0058 Total all 361 ± 100a 789 ± 67b 465 ± 43a F(2,9) = 11.2252; p = 0.0036 vVZ Unilateral BrdU-positive cell number 747 ± 54a 1513 ± 125b 751 ± 147a F(2,16) = 9.3280; p = 0.0021 Age Minimum age (months) 22 ± 3 22 ± 2 21 ± 2 F(2,18) = 0.1321; p = 0.8771 All values are the mean ± SEM. Superscript letters denote significant differences across treatment groups with post hoc Tukey’s test, only if the main effect of ANOVA was significant.
In this issue

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}



