

Article Figures & Data
Figures
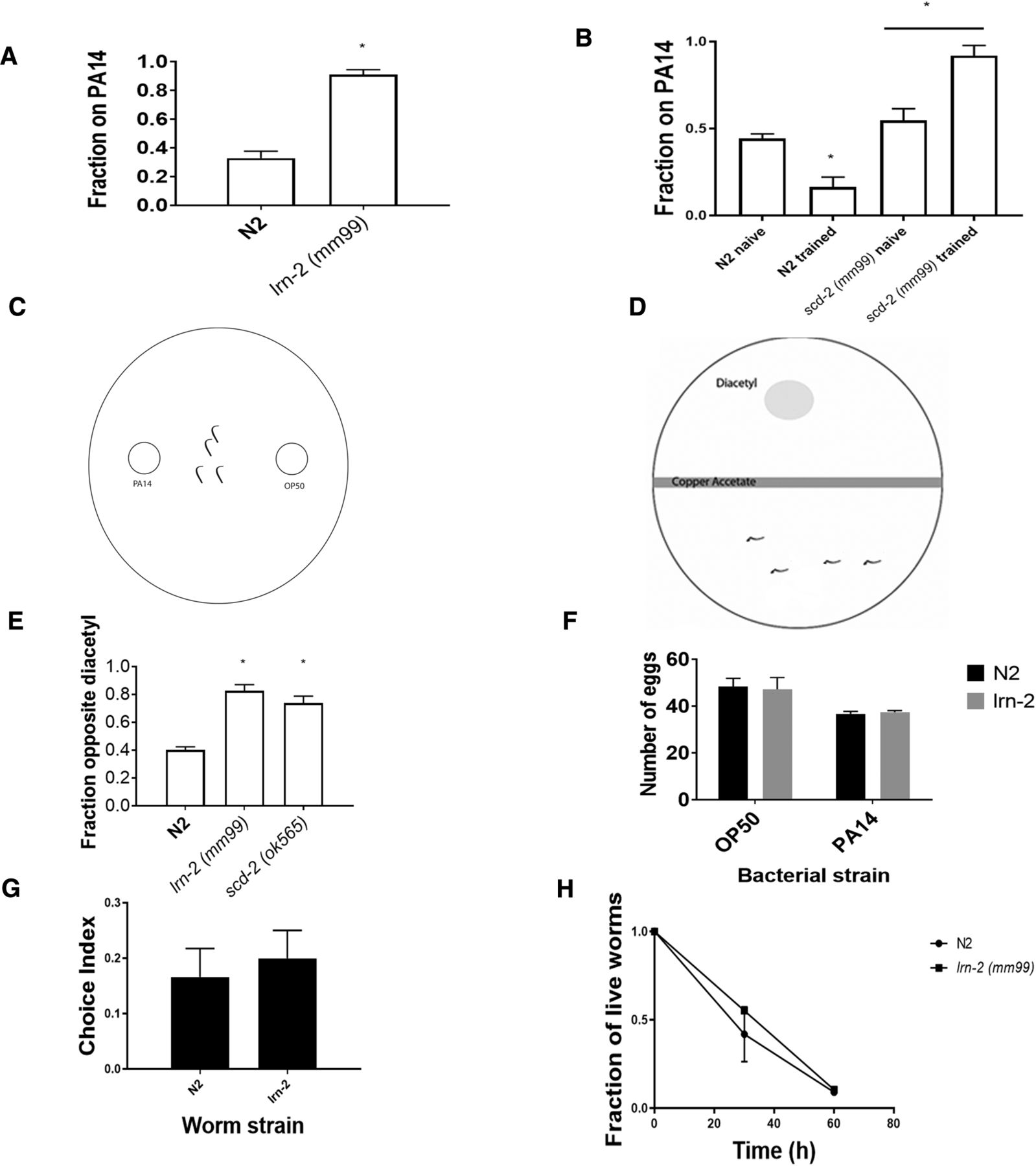
- Figure 1.
The lrn-2 mutation displays both associative learning and sensory integration deficits. A, This assay is modified from Zhang et al. (2005, their Fig. 1A). It simplifies the procedure by combining the testing and training phases into a single plate. When assayed for associative learning by pairing odor and pathogenicity of PA14, lrn-2 mutants did not learn to leave the pathogenic lawn, whereas N2 worms made the association and move to the safer OP50 bacteria. Learning was compared with wild-type N2 using a Student’s t test. *p < 0.05; n ≥ 3 plates; mean ± SEM. B, When assayed for associative learning, using the original method by Zhang et al. (2005, their Fig. 1A) that includes separate training and testing trials, lrn-2 mutants still did not learn a negative association with the pathogenic lawn (one-way ANOVA, F(3,14) = 27.54; p < 0.0001) with Bonferroni correction (*p < 0.05; n ≥ 5 plates; mean ± SEM), whereas N2 worms made the association and chose the safer OP50 bacteria when presented with point sources of OP50 and PA14 and compared with naïve worms that were not exposed to PA14 during training. This is comparable to the modified assay in A. lrn-2 mutants have an increased attraction to PA14 after training, which may be the result of appetitive learning that is masked by aversive learning in wild-type. Furthermore, this deficit may be caused by an inability to properly detect the pathogenicity of PA14 in the mutant. C, The experimental setup for the testing phase of the PA14 assay. After exposure to PA14 for 4 h, worms are transferred to a new plate, in which they are placed between two point sources of bacteria. One side of the plate has a point of PA14 and the other side has a point of OP50. Worms in the middle can crawl to either side depending one learning. D, The experimental setup for the sensory integration assay, based on Ishihara et al. (2002, their Fig. 1A). A Petri dish of NGM with a barrier of copper (II) acetate down the middle and a droplet of diacetyl on one side is used. Worms are placed opposite the diacetyl spot and after 1 h, worms on each side of the copper barrier are counted. Worms that are able to integrate two opposing sensory cues and cross the aversive barrier to reach diacetyl are considered to have normal sensory integration. E, Both lrn-2 (mm99) and scd-2 (sa249) failed to integrate copper and diacetyl cues, as they crossed the aversive copper barrier to reach an attractive diacetyl odor less than wild-type N2 (one-way ANOVA, F(2,6) = 30.83; p < 0.001) with Bonferroni correction (*p < 0.05; n ≥ 3 plates; mean ± SEM). F, N2 and lrn-2 both produced similarly fewer eggs on PA14 after 40 h compared with N2. Numbers of eggs produced (both laid and retained in the gonad over the 40 h assay) were analyzed using a two-way ANOVA. There was a significant main effect of bacterial strain (F(1,24) = 87.02; p < 0.0001; n ≥ 6 worms; mean ± SEM), but no significant effect of worm strain nor any significant interaction, showing that N2 and lrn-2 worms showed similar suppression of egg laying in response to food deprivation. G, Given a choice between PA14 and OP50 E. coli, both N2 and lrn-2 mutants showed a similar naïve preference for PA14 after 1 h (Student’s t test; n.s., not significant; n ≥ 6 plates; mean ± SEM), despite its pathogenicity. Positive chemotaxis index indicates PA14 approach. H, Both N2 and lrn-2 worms die at a similar rate when exposed to pathogenic bacteria (PA14), therefore it is unlikely that the difference in learning is caused by a resistance to PA14 pathogenicity. Using two-way ANOVA, there was no significant effect of time or strain.
- Figure 2.
scd-2 is the locus of the lrn-2 mutation. A, The F1 progeny of a cross between scd-2 (RB783) and a GFP tagged lrn-2 strain showed that these mutations do not complement, and that the cross progeny still have the mutant PA14 learning deficit compared with wild-type N2 using a one-way ANOVA (F(4,35) = 59.50; p < 0.0001) with Bonferroni correction (*p < 0.0001; n ≥ 7 plates; mean ± SEM). This indicates that the lrn-2 mutation is likely located in the scd-2 gene. B, A fosmid containing wild-type scd-2 was expressed in a lrn-2 (mm99) background. When tested for PA14 learning the worms expressing the fosmid and the pmyo2::mCherry coinjection marker showed a rescue of N2 like learning. The rescue phenotype was not significantly different when compared with wild-type N2 using a one-way ANOVA (F(4,27) = 18.47; p < 0.0001) with Bonferroni correction (*p < 0.01; n ≥ 4 plates; mean ± SEM). Non-glowing worms were counted as not containing the fosmid and show a learning deficit. These data suggest that expression of scd-2 can alleviate the mutant deficit, further indicating that it is the locus of the lrn-2 mutation. C, Four scd-2 mutant alleles containing both point mutations and deletions replicated the deficit in learning to avoid pathogenic bacteria displayed by lrn-2. Learning in lrn-2 was reduced compared with wild-type N2 worms as shown by a one-way ANOVA (F(5,9) = 76.74; p < 0.0001) with Bonferroni correction for multiple individual comparisons (*p < 0.01, n.s., not significant; n ≥ 3 plates; mean ± SEM). D, Other suppressor of constitutive dauer mutants, scd-1 and scd-3 do not show deficits in associative learning about PA14 (one-way ANOVA, F(3,13) = 13.18; p < 0.001; with Bonferroni correction, *p < 0.01, n.s., not significant; n ≥ 3 plates; mean ± SEM). This suggests that the learning deficit is not caused by the dauer-related phenotype alone.
- Figure 3.
There is a dissociation between associative learning and sensory integration at the genetic level. A, When tested in the diacetyl and copper assay for sensory integration, fsn-1;scd-2 double mutants showed a deficit similar to scd-2 single mutants compared with wild-type N2 (one-way ANOVA (F(3,12) = 21.29; p < 0.0001) with Bonferroni correction (*p < 0.05, n.s., not significant; n ≥ 4 plates; mean ± SEM). B, The fsn-1;scd-2 double mutants did not show a deficit in learning about pathogenic bacteria; instead the double mutants had a similar learned response to N2 or fsn-1 (hp1) single mutants (one-way ANOVA, F(5,22) = 15.32; p < 0.0001) with Bonferroni correction (*p < 0.05, n.s., not significant; n ≥ 4 plates; mean ± SEM). This suggests that SCD-2 and FSN-1 have a different interaction in associative learning compared with sensory integration.
- Figure 4.
AIA neurons are necessary for sensory integration and NSM neurons are necessary for associative learning. A, pgcy-28.d::scd-2;pmyo-3::GFP worms expressed wild-type scd-2 in AIA interneurons within an scd-2 mutant background. Expression in AIA rescued the worm’s ability to integrate sensory cues by crossing an aversive copper barrier to reach an attractive odorant. These data replicate the results originally found by Shinkai et al. (2011). Integration was compared with wild-type N2 using a one-way ANOVA (F(2,6) = 16.86; p < 0.005) with Bonferroni correction (*p < 0.0001; n ≥ 8 plates, mean ± SEM. B, Expression of wild-type scd-2 in AIA did not rescue associative PA14 learning. This indicates that the learning deficit seen in scd-2 mutants does not require AIA expression, unlike sensory integration (one-way ANOVA, F(2,23) = 52.6; p < 0.0001) with Bonferroni correction (*p < 0.05, n.s., not significant; n ≥ 3 plates; mean± SEM. C, Pceh-2::scd-2;Pmyo-2::mcherry worms expressed wild-type scd-2 in NSM neurons within an scd-2 mutant background. Expression in NSM did not rescue the ability to integrate sensory cues and cross an aversive copper barrier to reach an attractive diacetyl spot. These results are consistent with the findings that AIA expression is sufficient for sensory integration. Integration was compared with wild-type N2 using a one-way ANOVA (F(3,10) = 7.203; p < 0.01) with Tukey’s test for multiple comparisons (*p < 0.05; n ≥ 4 plates, mean ± SEM). D, Expression of wild-type scd-2 in NSM neurons within an scd-2 mutant background rescued associative learning about PA14. This indicates that the role of scd-2 in PA14 learning requires expression in NSM. Learning was compared with wild-type N2 using one-way ANOVA (F(3,24) = 17.17; p < 0.0001) with Tukey’s test for multiple comparisons (*p < 0.05; n ≥ 7 plates; mean ± SEM).
- Figure 5.
scd-2 mutants can form an associative memory independent of scd-2-mediated sensory integration. A, When N2 and scd-2 were tested for an associative memory in response to copper acetate post diacetyl and copper training, both strains showed an attenuated aversion to copper acetate. “Fraction on copper acetate side” refers to the fraction of worms on the side of the testing plate that had a point source of copper acetate. This represents the fraction of worms that are attracted to copper acetate. Learning was analyzed using a two-way ANOVA; there was a main effect of worm strain (F(1,12) = 6.954; p < 0.05), a main effect of training condition (F(2,12) = 12.08; p < 0.05), but no significant interaction. Tukey’s test for multiple comparisons was performed as a post hoc analysis (*p < 0.05 compared with naïve conditions, n.s., not significant; n ≥ 3 plates; mean ± SEM). These data indicate that while testing for sensory integration using diacetyl and copper, the worms learn that copper is associated with diacetyl, and thus decrease their aversive response to copper. Because scd-2 was still able to learn, despite deficits in PA14 associative learning, this memory is formed by a separate mechanism from associative PA14 learning. B, Testing to diacetyl instead of copper does not show learning perhaps because of a ceiling effect. N2 and scd-2 (mm99) worms were trained to associate diacetyl and copper, then approach to a point source of diacetyl was measured. However, diacetyl remained highly attractive across control and trained conditions. There is no significant difference between groups. C, N2 and scd-2 (mm99) worms were exposed to diacetyl for 1 h, followed by copper for 1 h (and vice versa), and then tested to a copper (II) acetate point source. Copper (II) acetate and diacetyl were also presented together for 1 h in a paired condition following 1 h on a plate with no odorants. These data indicate that diacetyl and copper (II) acetate should be presented simultaneously in order for a learned association to form. Learning was analyzed using a two-way ANOVA; there was a main effect of condition (F(2,41) = 19.73; p < 0.0001). Tukey’s test for multiple comparisons was performed as a post hoc analysis (*p < 0.05 compared with naïve conditions; n ≥ 9 plates; mean ± SEM). The slight difference between the two conditions in which copper and diacetyl were presented separately was not found to be statistically significant.





Tables
Data structure Type of test Power (α = 0.05) a (Fig. 1A) Normally distributed Student’s t test 0.0005 b (Fig. 1B) Normally distributed One-way ANOVA <0.0001 c (Fig. 1E) Normally distributed One-way ANOVA 0.0007 d (Fig. 1F) Normally distributed Two-way ANOVA Bacterial strain <0.0001 Worm strain 0.8379 e (Fig. 1G) Normally distributed Student’s t test 0.2824 f (Fig. 1H) Normally distributed One-way ANOVA 0.0003 g (Fig. 1I) Normally distributed Two-way ANOVA Strain 0.8493 Time h (Fig. 2A) Normally distributed One-way ANOVA <0.0001 i (Fig. 2B) Normally distributed One-way ANOVA <0.0001 j (Fig. 2C) Normally distributed One-way ANOVA <0.0001 k (Fig. 3A) Normally distributed One-way ANOVA <0.0001 l (Fig. 3B) Normally distributed One-way ANOVA <0.0001 m (Fig. 4A) Normally distributed One-way ANOVA 0.0034 n (Fig. 4B)o (Fig. 4C)p (Fig. 4D) Normally distributedNormally distributedNormally distributed One-way ANOVAOne-way ANOVAOne-way ANOVA <0.00010.0074<0.0001 q (Fig. 5A) Normally distributed Two-way ANOVA Worm strain 0.0217 r (Fig. 5B)s (Fig. 5C) Normally distributedNormally distributed Two-way ANOVATwo-way ANOVA Training cond. 0.0013Worm strain 0.0986Training cond. 0.6410Worm strain 0.1643Training cond. <0.0001
In this issue

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}



