

Article Figures & Data
Figures
- Figure 1.
The experimental chamber. a, The box was separated into a start box and an arena. Animals were put in the start box at the start of the trials. The animals were allowed to enter the arena through the door opened and closed by an experimenter. There was a bay window on the opposite side of the door in the arena. b, The photograph shows a rat approach the bay window after the opening of the door at the start of a trial. c–e, A diagram and photograph of the bay window. The rats put their heads into the bay window as shown in d to lick a spout in front of the bay window. Infrared LEDs below and sensors above the bay window detected the entry of the heads. The metal spout was connected with a touch sensor via a metal clip and wire.
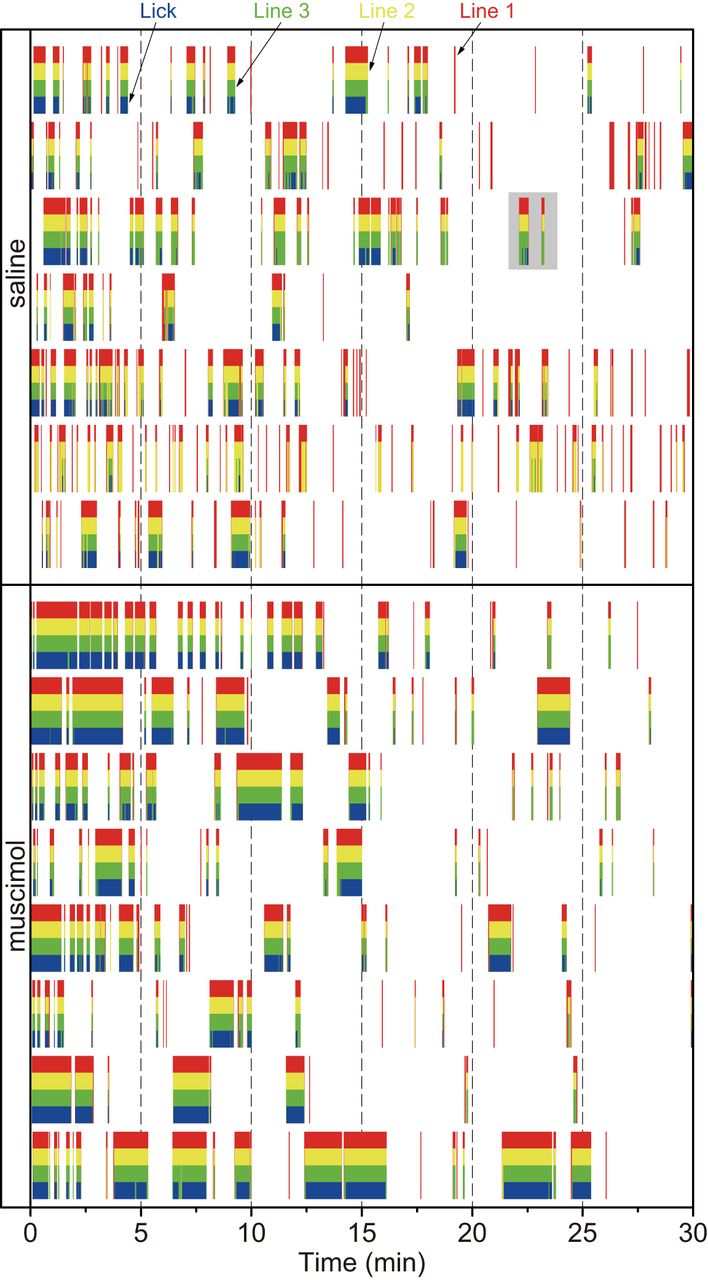
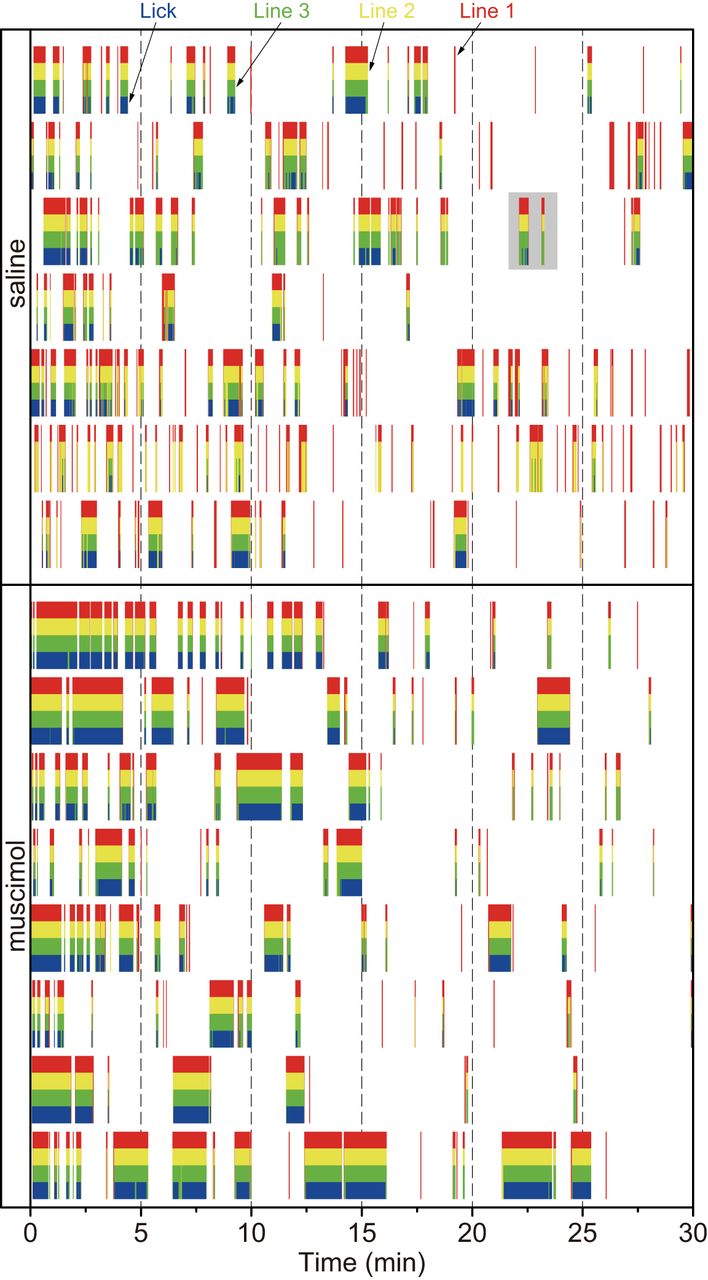
- Figure 2.
Raster plots of the responses of the passing and touch sensors in each animal on Test 2. The red, yellow, and green vertical lines indicate Line 1 (far from the spout), Line 2 (middle of the bay window), and Line 3 (close to the spout), respectively. The red color bars indicate that the rat’s head (tip of nose) stopped at the entrance of the bay window. Both the red and yellow color bars indicate the stop at the middle of the bay window, and the sets of the red, yellow, and green bars indicate the stop just before the spout. The plots with a gray background in the saline group are enlarged in Figure 3 for explaining behavioral analyses.
- Figure 3.
Scheme for behavioral analyses. a, The analysis of the entrance of the rat’s heads into the bay window (Entry). Top, The blocks containing all the color bars indicate Entry-Lick, while the ones containing all the colors without blue indicate Entry-Stop. Bottom, Illustrations showing the animal’s behavior on Entry-Lick and Entry-Stop. b, Schematic diagram of the licking pattern. The vertical lines indicate licks.
- Figure 4.
Histologic findings. The locations of the injection cannulae tips were reconstructed based on the Paxinos and Watson brain atlas (Paxinos and Watson, 2007). Orange circles, muscimol-injected rats; black squares, saline-injected rats.
- Figure 5.
The intra-BLA muscimol altered Entry. a, The mean duration of all Entry in the saline-injected (SAL) and muscimol-injected (MUS) groups on Conditioning, Test 1, and Test 2. After the CS-US pairings, the mean duration of Entry decreased in both groups. The microinjections of muscimol into the BLA on Test 2 impaired the decline in the mean duration of Entry. b, The proportion of the frequency of Entry-Lick and Entry-Stop. The conditioning decreased Entry-Lick and increased Entry-Stop. The MUS group showed smaller proportion of Entry-Stop and larger Entry-Lick than the SAL group on Test 2; ***p < 0.001 (vs SAL), *p < 0.05, †p < 0.05 (vs Conditioning), $$p < 0.01 (vs Test 1).
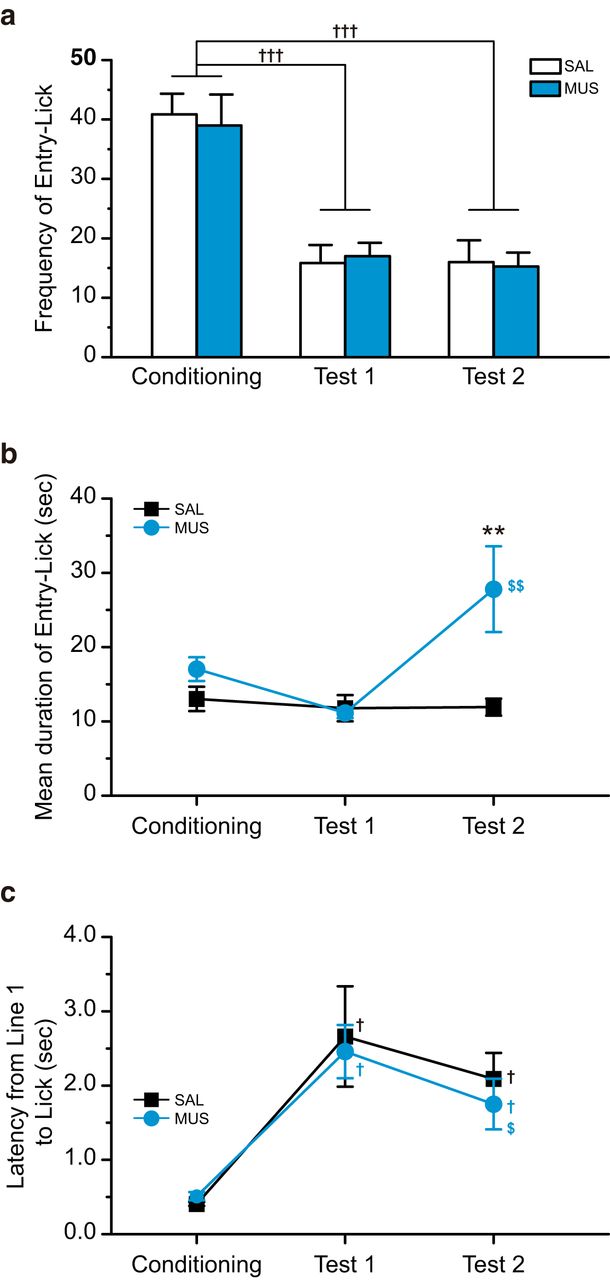
- Figure 6.
The intra-BLA muscimol decreased the mean duration of Entry-Lick. a, The frequency of Entry-Lick in the saline-injected (SAL) and muscimol-injected (MUS) groups on Conditioning, Test 1, and Test 2. The CS-US pairings considerably decreased the frequency of Entry-Lick. Both the SAL and MUS groups showed suppressed frequency of Entry-Lick on Test 2. There was no group difference. b, The mean duration of Entry-Lick. The rats showed a slightly shorter mean duration of Entry-Lick on Test 1 than Conditioning. On Test 2, the SAL group showed the same level of mean duration with Test 1, while the MUS group demonstrated significantly longer mean duration than Test 1 and the SAL group on Test 2. c, The latency from the cut of Line 1 to Lick. The CS-US pairings significantly increased latency. The microinjections of muscimol into the BLA did not alter the latency in Test 2; **p < 0.01 (vs SAL), †††p < 0.001, †p < 0.05 (vs Conditioning), $$p < 0.01, $p < 0.05 (vs Test 1).
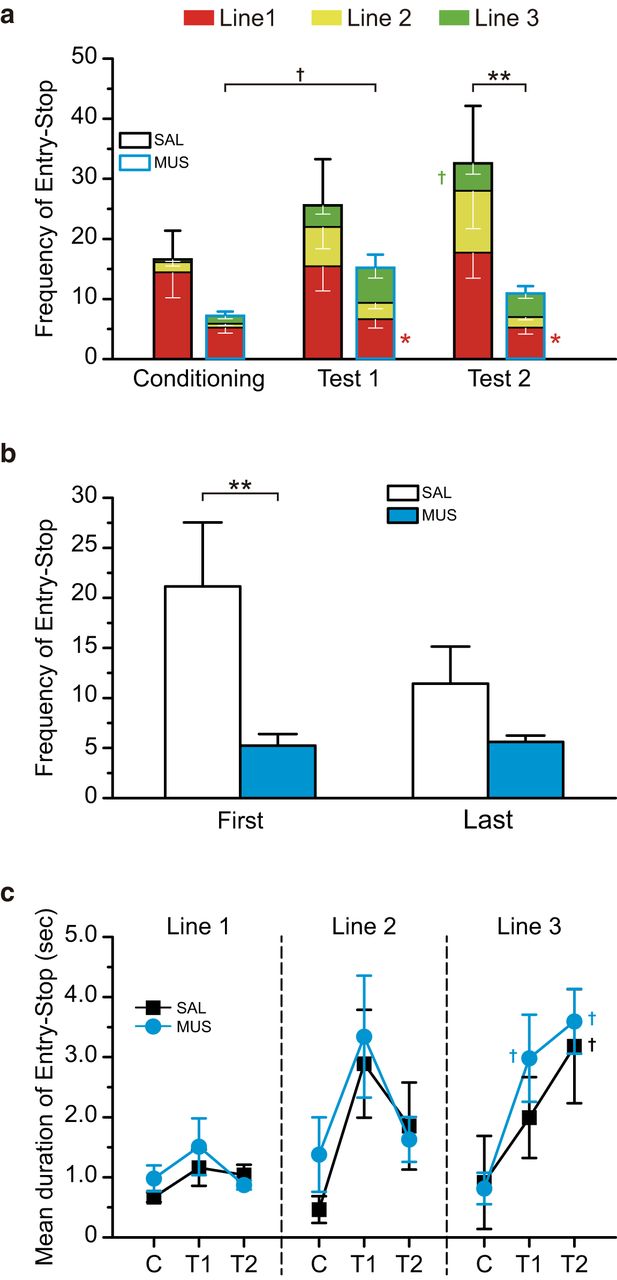
- Figure 7.
The intra-BLA muscimol decreased the frequency of Entry-Stop. a, The frequency of Entry-Lick in the saline-injected (SAL) or muscimol-injected (MUS) group in Conditioning, Test 1, and Test 2. The CS-US pairings increased entry-stop. On test 2, the MUS group showed significantly smaller stops than the SAL group. b, The frequency of Entry-Stop during the first and last half of Test 2. The muscimol-injected group showed smaller frequency of the stop on both the time windows. There was a significant group difference for the first half. c, Mean duration of Entry-Stop at each line. The CS-US pairings significantly increased mean duration at lines 2 and 3. The mean duration at Line 2 decreased in both groups for Test 2 and increased at Line 3 for Test 2. However, there were no differences between the groups. C, Conditioning; T1, Test 1; T2, Test 2; **p < 0.01, *p < 0.05 (vs SAL), †p < 0.01 (vs Conditioning).
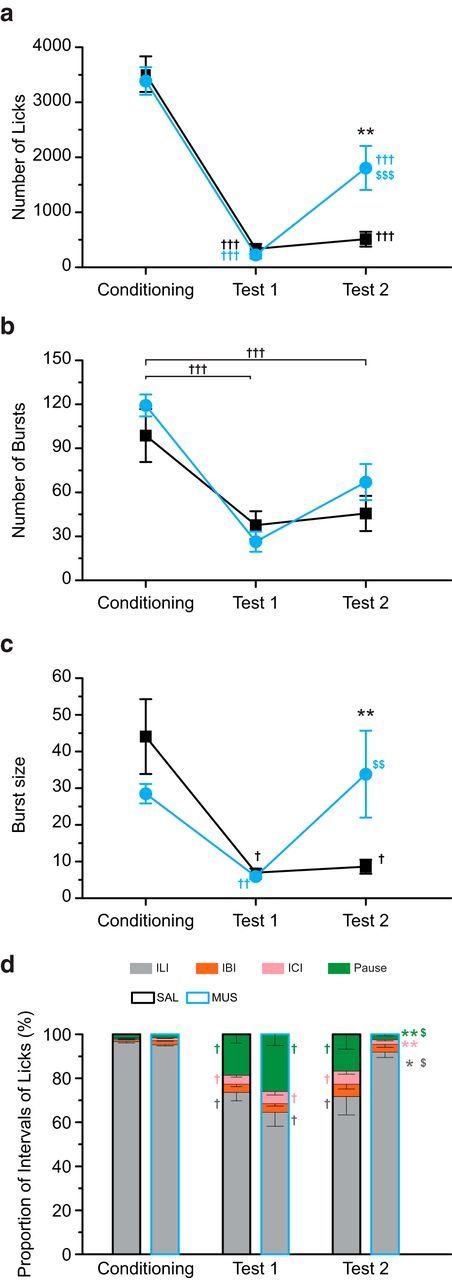
- Figure 8.
The intra-BLA muscimol attenuated aversion to the CS. a, The total number of licks in the saline-injected (SAL) and muscimol-injected (MUS) groups on Conditioning, Test 1, and Test 2. The CS-US pairings significantly decreased the total number of licks. The MUS group showed larger CS licking than the SAL group on Test 2. b, The number of bursts. The CS-US pairings decreased the burst number. There was no group difference for Test 2. c, The size of the burst. The CS-US pairings decreased the burst size. The MUS group demonstrated a larger burst size than the SAL group for Test 2. d, The proportion of ILI. The CS-US pairings increased the proportions of IBI, ICI, and pause, which were smaller in the MUS group than in the SAL group; **p < 0.01, *p < 0.05 (vs SAL), †††p < 0.001, ††p < 0.01, †p < 0.05 (vs Conditioning), $$$p < 0.001, $$p < 0.01, $p < 0.05 (vs Test 1).
- Figure 9.
The intra-BLA muscimol increased larger burst size. Plots for all burst sizes in the saline-injected (SAL) and muscimol-injected (MUS) groups for Test 2. a, The size of almost all the bursts was <100 licks in the SAL group. b, The MUS group showed a larger number of lick bursts containing over 100 licks. c, There was a significant group difference in the average number of >100 lick burst size; *p < 0.05 (vs SAL).







Tables
Conditioning Test 1 Test 2 SAL MUS SAL MUS SAL MUS Latency of 1st entry 4.66 ± 0.84 5.91 ± 0.63 8.73 ± 0.93 4.18 ± 0.94 13.34 ± 4.27 4.46 ± 0.87* Frequency of entry 57.43 ± 5.89 46.13 ± 4.86 41.43 ± 7.30 32.13 ± 4.02 67.86 ± 21.4 26.13 ± 2.91††** Total duration of entry 534.6 ± 64.14 625.93 ± 35.99 243.19 ± 41.56 239.63 ± 41.57 255.67 ± 32.15 393.18 ± 62.49 ** p < 0.01.
↵* p < 0.05 (vs SAL).
†† p < 0.01 (vs Conditioning).
In this issue

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}



