

Article Figures & Data
Figures
- Figure 1.
Response adaptation is altered by a change in the stimulation pattern. A, Serial brain slices of a representative case in which the electrode track is observed. An example of spike responses is shown on the right (bottom recording represent stimuli). B, A schematic diagram of the experimental protocol and stimulation patterns. C, Representative PSTHs of a single unit located in layer 5 show response adaptation when a regular stimulation pattern is applied (left PSTH). Response adaptation is reduced if the interval between the first and the second stimuli increases from 250 to 375 ms (accelerando stimulation pattern; middle PSTH) or decreases to 125 ms (decelerando stimulation pattern; right PSTH). Lines on the PSTHs show the stimulation pattern. Vertical dashes lines indicate 50 ms poststimulus time window in which spike responses are calculated. Figure Contributions: Natali Barros-Zulaica performed the experiments. All authors analyzed the data.
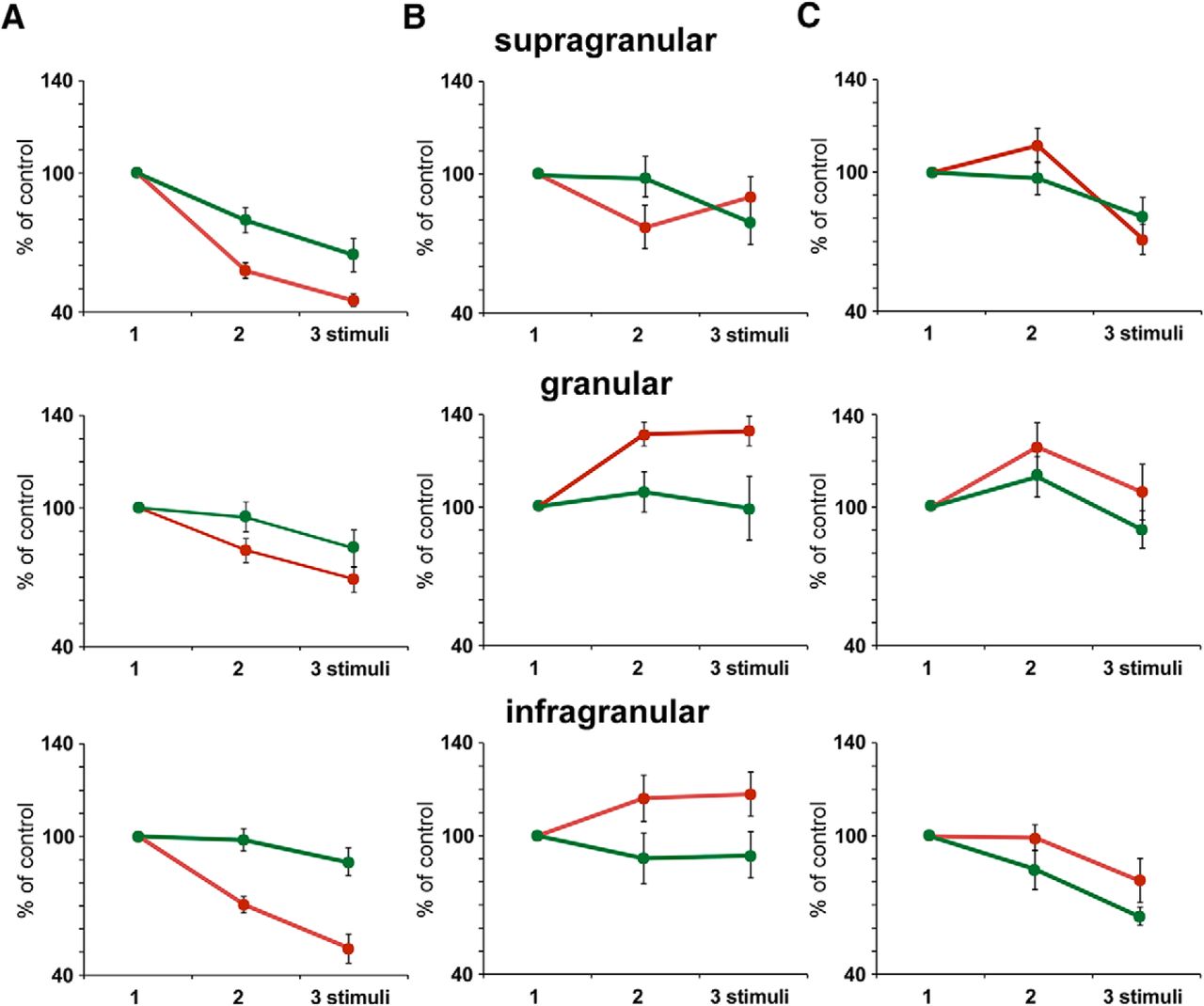
- Figure 2.
Response adaptation is altered by a change in the stimulation pattern. A, Supragranular neurons (n = 14) show a reduction in spike response during the application of the regular stimulation pattern (three stimuli at 4 Hz; blue circles). The mean percentage of the response respect to the control stimulus (first stimulus of the stimulation train) is shown on the left. The response adaptation is reduced if the interval between the first and the second stimuli increases from 250 to 375 ms (accelerando stimulation pattern; orange circle) or decreases to 125 ms (decelerando stimulation pattern; gray circle). The plots shown on the right display differences in the percentage of responses to the 1–3 stimuli during the application of the accelerando or decelerando stimulation patterns respect to the corresponding percentage to the 1–3 stimuli during the application of a regular stimulation pattern. In all cases the response to the second and third stimuli is higher than is expected during the regular stimulation pattern. B, C, Same plots as in A from granular (n = 12) and infragranular (n = 23) neurons, respectively. Equally, responses during the accelerando or decelerando stimulation pattern are greater respect to the corresponding response during the application of a regular stimulation pattern. *p < 0.05; **p < 0.01. Figure Contributions: Natali Barros-Zulaica performed the experiments. All authors analyzed the data.
- Figure 3.
Response adaptation in thalamic neurons is not altered by the stimulation pattern. A, The mean percentage of the response respect to the control stimulus (first stimulus of the stimulation train) is shown for VPM (n = 12). B, Same plot that in A for POm (n = 8) neurons. Response adaptation is not altered by the stimulation pattern (regular, accelerando or decelerando stimulation patterns. Right, Brain slices of representative cases in which the electrode track is observed. Scale bars, 1 mm. Figure Contributions: Natali Barros-Zulaica performed the experiments. All authors analyzed the data.
- Figure 4.
Response adaptation is altered during the stimulation period. Plots of the mean percentage of the response respect to the control stimulus (first stimulus of the stimulation train; 100%) are shown. Data are calculated from PSTHs calculated with the first 25 stimulation trains (red) or with the last 25 stimulation trains (green) of the 100 stimulation train block applied to BC neurons. A, Response adaptation during the application of the regular stimulation pattern is larger in the first 25 stimulation trains for supragranular (n = 14; top plots), granular (n = 12; middle plots), and infragranular (n = 23; bottom plots) neurons. B, C, Same plots that in A for neurons recorded during the application of accelerando or decelerando stimulation patterns, respectively. In all cases the response to the second and third stimuli is higher in the first stimulation trains than in the last stimulation train of the stimulation block. Figure Contributions: Natali Barros-Zulaica performed the experiments. All authors analyzed the data.
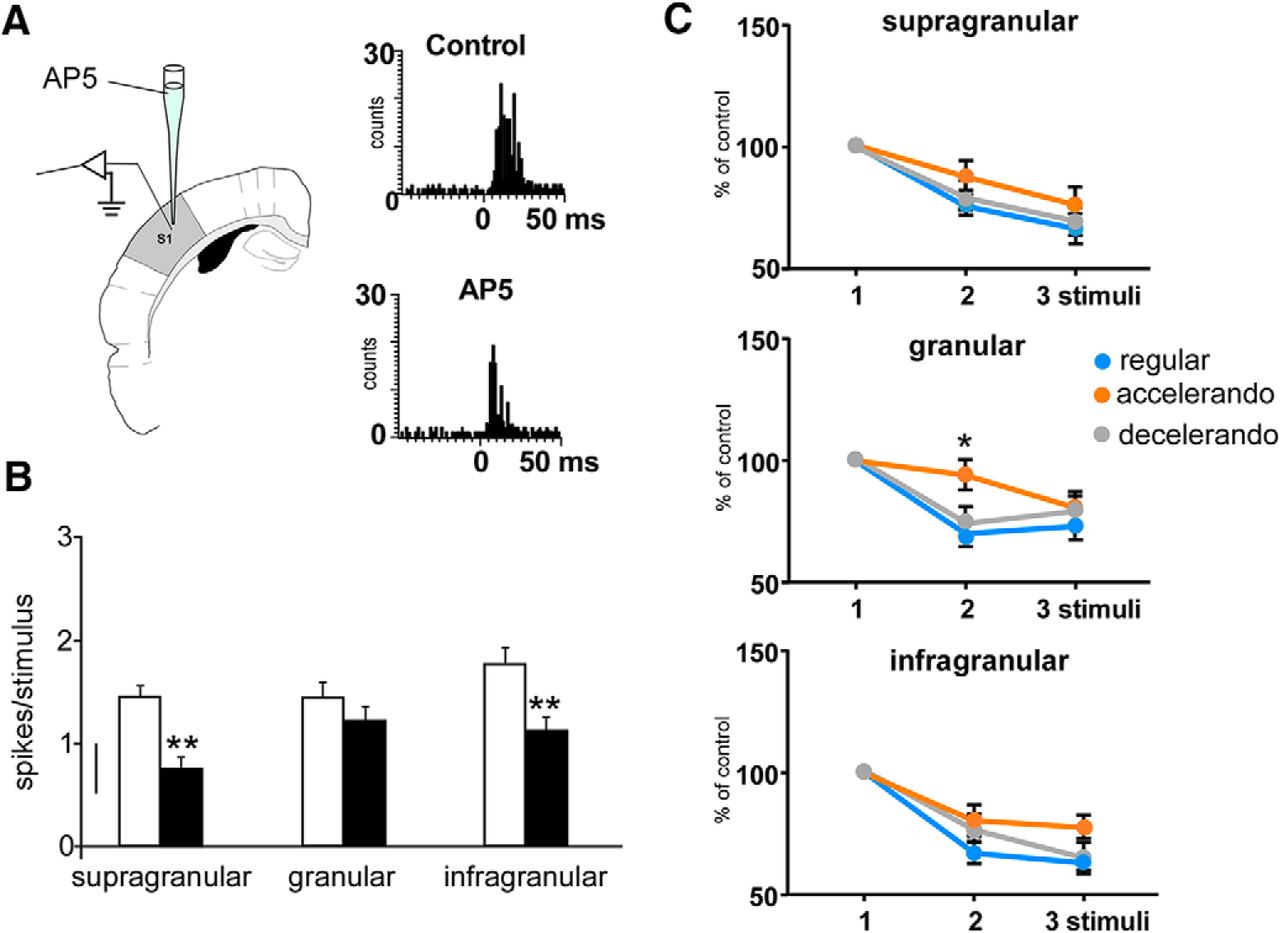
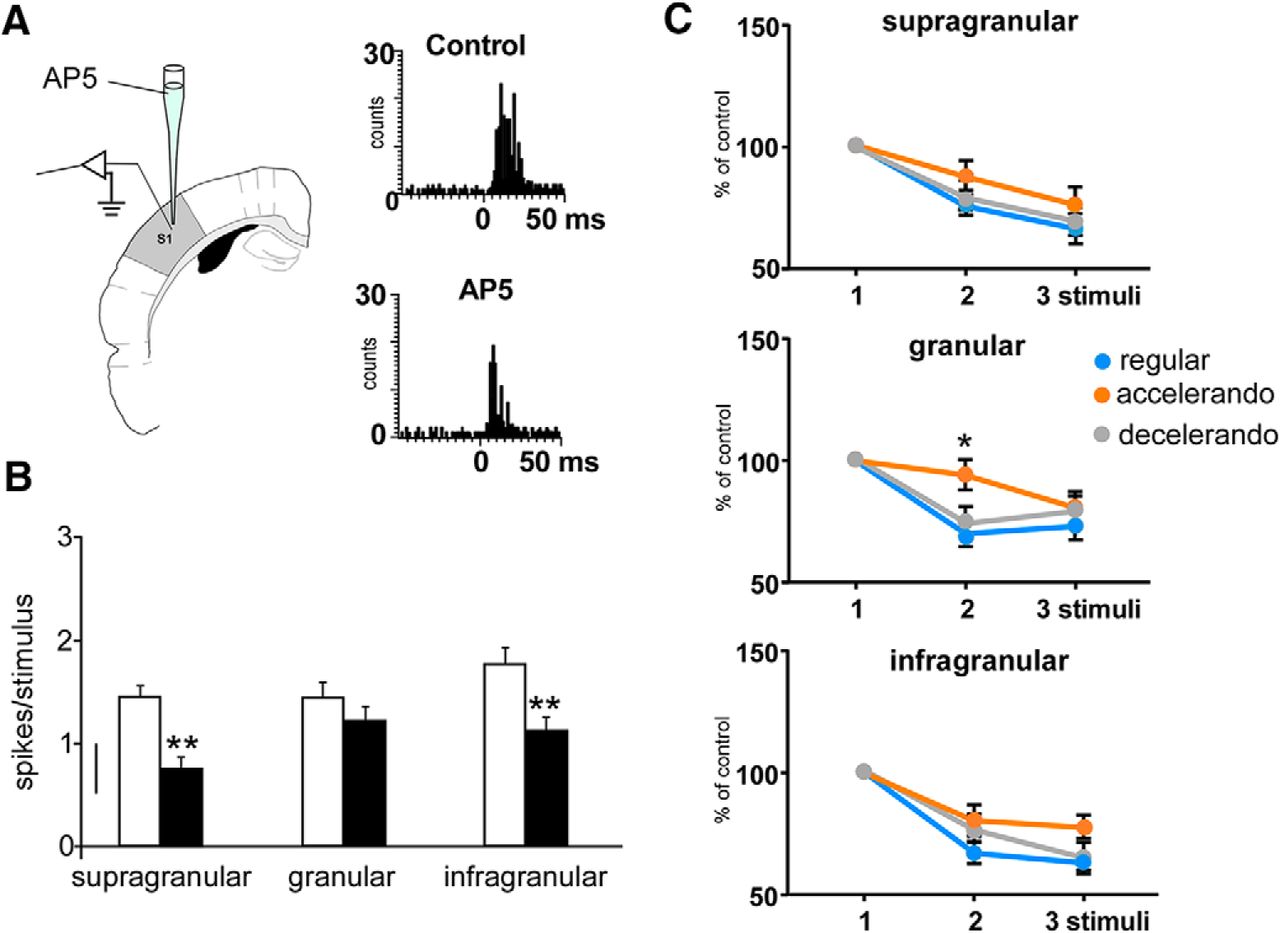
- Figure 5.
Changes in the response adaptation induced by the stimulation pattern are because of activation of NMDA receptors. A, Schematic diagram of the experimental protocol. Insets, PSTHs of the response of layer 5 neurons in the control condition and 10 min after AP5 application (50 μm, 0.1 μl; black bars); the response is clearly reduced. B, Responses to whisker stimuli are reduced after cortical injection of AP5 (50 μm, 0.1 μl; black bars) respect to control values (white bars) in supragranular (n = 8), granular (n = 7), and infragranular (n = 8) neurons. C, Mean percentage of the response respect to the control stimulus (first stimulus of the stimulation train) is shown. Supragranular and infragranular neurons adapt equally in presence of AP5 when the regular, accelerando, or decelerando stimulation patterns are applied. Granular neurons also show response adaptation during the application of the regular and decelerando stimulation patterns after AP5 application. However, a significant reduction of adaptation is observed during the application of the accelerando stimulation pattern. *p < 0.05; **p < 0.01. Figure Contributions: Angel Nuñez performed the experiments. All authors analyzed the data.
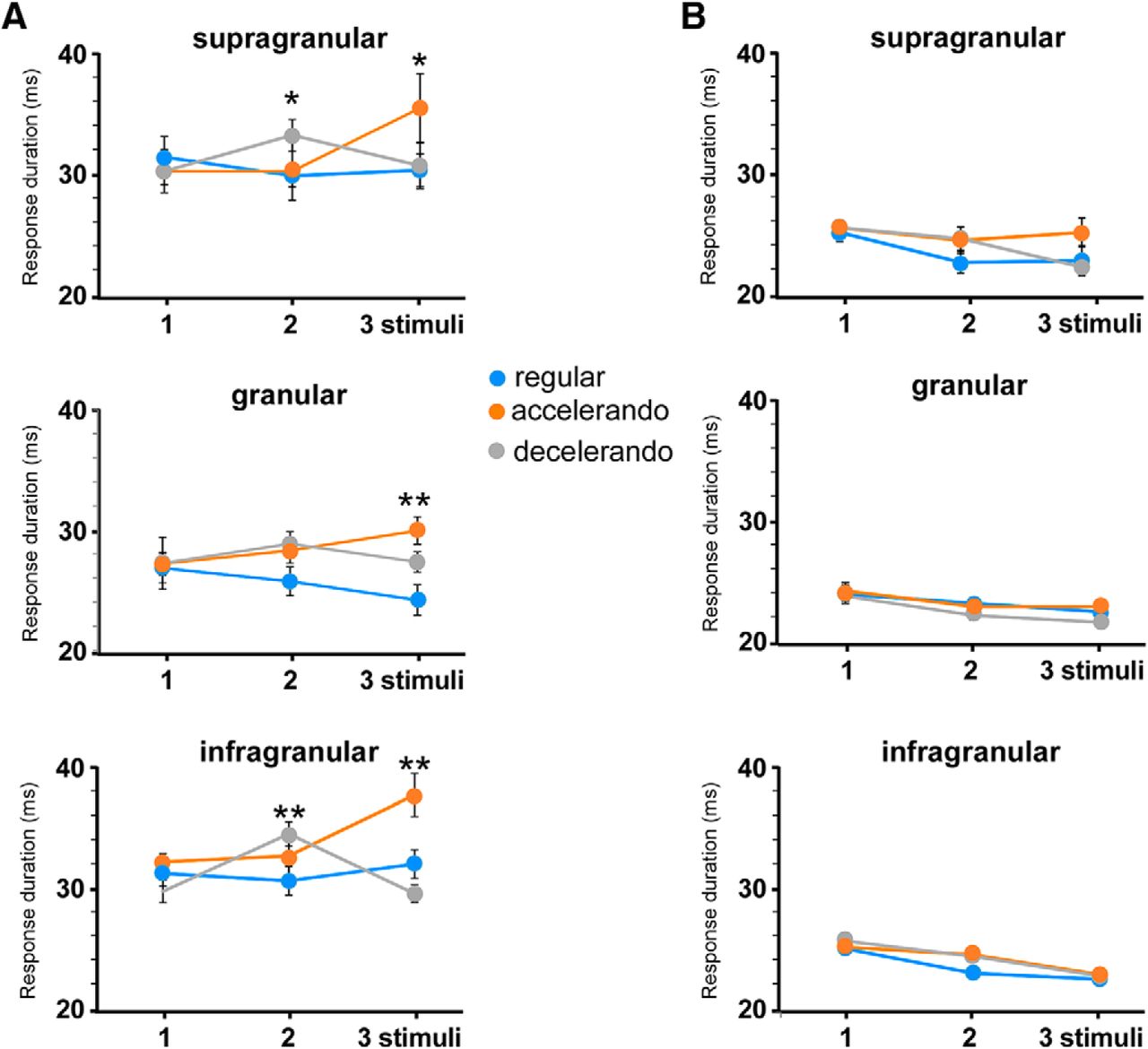
- Figure 6.
The stimulation pattern also induces changes in the whisker response duration. A, Response duration according to the stimulation pattern in each cortical layer. B, Application of AP5 into the BC (50 μm; 0.1 μl) reduces the duration of whisker responses and blocks the effect of the stimulation pattern. *p < 0.05; **p < 0.01. Figure Contributions: Natali Barros-Zulaica and Angel Nuñez performed the experiments. All authors analyzed the data.
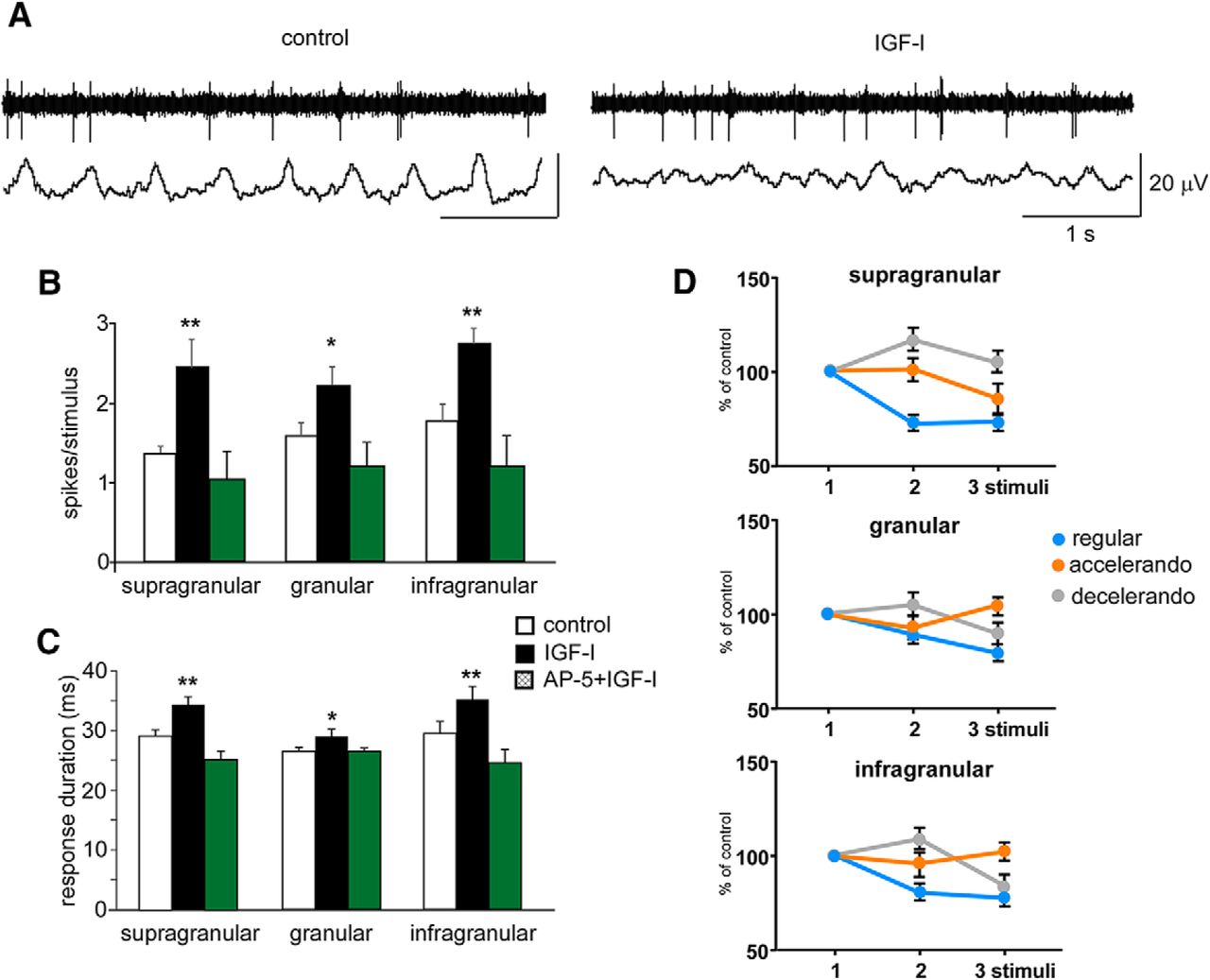
- Figure 7.
Changes in the response adaptation induced by the stimulation pattern are also observed during periods of EEG activation. A, Unit and field potential activities (top and bottom traces, respectively) are shown in a representative case recorded in the supragranular layer. Intraperitoneal injection of IGF-I (1 μg/g body weight) induced a reduction of slow waves and an increase in the spontaneous firing rate. B, Responses to whisker stimuli are increased after IGF-I intraperitoneal injection (closed bars) respect to control values (white bars) in supragranular (n = 6), granular (n = 7), and infragranular (n = 9) neurons. Facilitation by IGF-I of whisker responses is blocked after AP5 (50 μm; 0.1 μl) injection in the BC (green bars). C, IGF-I injection also induces an increase of the response duration (closed bars). Similarly, the increase in the response duration evoked by IGF-I is blocked if AP5 (50 μm; 0.1 μl) is injected previously (green bars). D, Mean percentage of the response respect to the control stimulus (first stimulus of the stimulation train) is shown. Response adaptation to the regular stimulation pattern is reduced and the facilitation of responses during the application of the accelerando or decelerando stimulation patterns are enhanced in supragranular, granular and infragranular layers after IGF-I administration. *p < 0.05; **p < 0.01. Figure Contributions: Angel Nuñez performed the experiments. All authors analyzed the data.




Tables
- Table 1.
Cortical responses to the whisker stimulation train (St. 1–3) according to the stimulation pattern (spikes/stimulus)
Layer Regular Accelerando Decelerando St. 1 St. 2 St. 3 St. 1 St. 2 St. 3 St. 1 St. 2 St. 3 Supragranular 1.37 ± 0.26
n = 140.89 ± 0.18
n = 140.69 ± 0.1
n = 141.26 ± 0.20
n = 141.17 ± 0.23
n = 141.07 ± 0.29
n = 141.56 ± 0.20
n = 141.55 ± 0.26
n = 141.12 ± 0.15
n = 14Granular 1.26 ± 0.16
n = 121.17 ± 0.16
n = 120.99 ± 0.1
n = 121.56 ± 0.25
n = 121.84 ± 0.27
n = 121.87 ± 0.37
n = 121.49 ± 0.25
n = 121.83 ± 0.3
n = 121.44 ± 0.25
n = 12Infragranular 1.88 ± 0.29
n = 231.33 ± 0.13
n = 231.18 ± 0.1
n = 231.77 ± 0.19
n = 231.66 ± 0.19
n = 231.73 ± 0.25
n = 231.94 ± 0.18
n = 231.81 ± 0.2
n = 231.42 ± 0.17
n = 23 - Table 2.
Duration of cortical responses to whisker stimulation trains of three stimuli (St. 1–3), according to the stimulation pattern (in ms)
Layer Regular Accelerando Decelerando St. 1 St. 2 St.3 St. 1 St. 2 St. 3 St. 1 St. 2 St. 3 Supragranular 31.8 ± 1.5
n = 1129.8 ± 0.9
n = 1130.3 ± 1.3
n = 1130.2 ± 1.8
n = 1130.2 ± 2.4
n = 1135.5 ± 2.9
n = 1130.2 ± 1.1
n = 1133.2 ± 1.3
n = 1130.6 ± 1.9
n = 11Granular 27.0 ± 1.2
n = 1125.9 ± 1.3
n = 1124.4 ± 1.3
n = 1127.4 ± 0.9
n = 1128.5 ± 1.1
n = 1130.1 ± 1.1
n = 1127.4 ± 2.1
n = 1129.0 ± 1.0
n = 1127.5 ± 0.8
n = 11Infragranular 31.1 ± 1.0
n = 1630.3 ± 1.3
n = 1631.7 ± 1.3
n = 1631.9 ± 0.7
n = 1632.4 ± 1.6
n = 1637.4 ± 1.8
n = 1629.4 ± 0.9
n = 1634.2 ± 1.0
n = 1629.2 ± 0.7
n = 16
In this issue

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}



