

Abstract
In this study, we investigated the effect of transcranial alternating current stimulation (tACS) on voluntary risky decision making and executive control in humans. Stimulation was delivered online at 5 Hz (θ), 10 Hz (α), 20 Hz (β), and 40 Hz (γ) on the left and right frontal area while participants performed a modified risky decision-making task. This task allowed participants to voluntarily select between risky and certain decisions associated with potential gains or losses, while simultaneously measuring the cognitive control component (voluntary switching) of decision making. The purpose of this experimental design was to test whether voluntary risky decision making and executive control can be modulated with tACS in a frequency-specific manner. Our results revealed a robust effect of a 20-Hz stimulation over the left prefrontal area that significantly increased voluntary risky decision making, which may suggest a possible link between risky decision making and reward processing, underlined by β-oscillatory activity.
- 20-Hz stimulation
- frontal hemisphere
- reward
- risky decision making
- task switching
- transcranial alternating current stimulation
Significance Statement
This is the first study that shows a frequency-specific effect on risky decision making demonstrated by online 20-Hz transcranial alternating current stimulation (tACS) applied to left dorsolateral prefrontal cortex (PFC). Our results suggest that left frontal 20-Hz tACS specifically modulates risky decision making, perhaps by entraining endogenous β-activity underlying a frontal-striatal network associated with gain anticipation.
Introduction
Much research has been conducted on the neurobiological mechanisms of risky decision making demonstrating a large neural network comprised of the ventral striatum, amygdala, insula, cingulate, and prefrontal cortices (PFCs; Knutson et al., 2001a,b; O'Doherty et al., 2001; Kuhnen and Knutson, 2005; Rao et al., 2008; Fujiwara et al., 2009; Mohr et al., 2010; Kohls et al., 2013). In particular, the PFC plays an important role in voluntary risky decision making. For instance, Rao et al. (2008) demonstrated a link between the PFC and voluntary decisions to accept greater risk. They suggested that the PFC mediates the active volitional control or agency of the risk taker by means of an executive control component.
The PFC also plays a prominent role in executive control (Derrfuss et al., 2005; Swick et al., 2011; Kim et al., 2012; Rottschy et al., 2012; also, see Owen et al., 2005, for fMRI meta-analyses on executive functions), which in turn comprises of three separate, independent components; working memory updating, inhibition, and set shifting/task switching (Miyake et al., 2000; Diamond, 2013). Risky decision making and executive control have been thoroughly investigated. Inspired by Kahneman’s dual process theory, that irrational decision making increases when cognitive resources become depleted (Kahneman, 2003; Kahneman and Frederick, 2007; Kahneman, 2011), some have tested the influence of executive control on risky decision making by administering the n-back task, a popular working memory task, in parallel with various risky decision-making tasks (Whitney, Rinehart and Hinson, 2008; Starcke et al., 2011; Farrell et al., 2012; Pabst et al., 2013; Gathmann et al., 2014a,b). Likewise, many have examined inhibitory processes and risky decision making by employing the Go No-Go (Verdejo-García et al., 2007; Yeomans and Brace, 2015; Ba et al., 2016; Welsh et al., 2017). However, to date few have examined the link between set-switching and risky decision making (Verdejo-García et al., 2007; Fröber and Dreisbach, 2016); therefore, we proposed to investigate this link by using brain stimulation of the PFC.
θ-Related activity (4–8 Hz) has been inferred to reflect aspects of risky decision making and executive control. While numerous accounts have associated θ-band oscillations with executive control functions (e.g., working memory, set-switching, conflict monitoring, error detection; Jensen and Tesche, 2002; Sauseng et al., 2006; Cunillera et al., 2012; Cavanagh and Frank, 2014), a recent EEG study reported fronto-central θ-oscillations inferred to reflect an action monitoring system that compares potential outcomes of high- and low-risk options (Zhang et al., 2014). Furthermore, θ-band transcranial alternating current stimulation (tACS) applied on the left PFC was demonstrated to increase risky decision making (Sela et al., 2012). This stimulation technique allegedly entrains ongoing electrophysiological oscillatory activity (Veniero et al., 2015; Vosskuhl et al., 2015; Thut et al., 2011; Helfrich et al., 2014a), suggesting that theta tACS entrains frontal-central theta oscillations. However, a disadvantage to this study is that frequency specificity could not be assessed since the authors did not control for other stimulation frequencies. In other words, the increase in risky decision making may have been driven by the stimulation alone and not necessarily by theta stimulation (for further details, see Feurra et al., 2012).
For this study, we tested whether voluntary risky decision making under varied levels of executive control can be modulated by applying online tACS at various frequencies (sham, 5, 10, 20, and 40 Hz) to the left and right frontal hemispheres. To isolate these components of decision making, we adopted and modified a task-switching paradigm that allows participants to choose between risky and safe (certain) decisions depending on the decision to switch or repeat between task sets (Arrington and Logan, 2004, 2005; Weaver and Arrington, 2013; Arrington et al., 2014; Orr and Banich, 2014; Poljac and Yeung, 2014; Fröber and Dreisbach, 2016). Although relatively new for cognitive neuroscience (Orr and Banich, 2014; Poljac and Yeung, 2014), the voluntary task-switching paradigm is well established within the cognitive psychological literature (Arrington and Logan, 2004, 2005; Weaver and Arrington, 2013; Fröber and Dreisbach, 2016). However, unlike typical executive tasks, in which participants are rated on response time and accuracy (e.g., N back, Go Go-No task, Eriksen Flanker task, Wisconsin Card Sorting task), the voluntary task-switching paradigm investigates voluntary executive control by considering choice as a dependent variable. By combining the voluntary task-switching paradigm with two-choice financial decision-making task between lotteries involving risk, it is possible to measure how much executive control participants are willing to exert under the condition of risk. The advantage of this task design was the possibility to measure voluntary executive control and voluntary risky decision making within a single response, thus allowing us to test whether tACS can modulate voluntary risky decision making under varied levels of voluntary executive control. Given that voluntary, but not involuntary, risky decision making yields frontal-ventral striatum activity (Rao et al., 2008), we hypothesize that θ-band tACS should modulate voluntary risky decision making under high levels of executive control.
Materials and Methods
Participants
Thirty-four healthy right-handed participants (21 females; mean age 21; age range 18–26 years; SD = 2.54) with normal or corrected to normal vision and with no neurologic disorders participated in the study. All participants provided a written consent approved by a local ethics committee in accordance with the Declaration of Helsinki. All participants were screened for psychological/psychiatric disorders and none of them reported use of drugs or alcohol in the days preceding the experiment. Participants were divided into two groups: those who received stimulation on the left frontal area (n = 17; 10 females; mean age 20.52; age range 18–25 years; SD = 2.52) and those who received stimulation on the right frontal area (n = 17; 11 females; mean age 21.17; age range 18–26 years; SD = 2.78).
Stimuli and procedure
Participants performed a novel neuroeconomic risky decision-making task that combines binary lotteries with equal expected value (Selten et al., 1999; Engelmann and Tamir, 2009; Harrison et al., 2013), and the voluntary task-switching paradigm (Arrington and Logan, 2004, 2005) that allows participants to select between risky or certain decisions by switching or repeating task sets between trials. Each trial began with a centered fixation cross which remained between 500 and 1000 ms followed by the stimuli screen, composed of a randomly selected single digit (1, 2, 3, 4, 6, 7, 8, or 9) centered on the screen until the participant responded. For each trial participants had to select one of the two games: odd/even game (participants indicated whether the digit was odd or even) or higher/lower game (participants indicated whether the digit was higher or lower than 5) by pressing one of the corresponding buttons (odd, even, high, low). Using a randomized Latin-square blocked design, the instruction varied across blocks as described below.
In the basic version of the task (Fig. 1A, “switch = risk” blocks) participants were instructed that if they chose to repeat the same game in successive trials they would make a certain decision, e.g., they select “odd” button for the digit 3 on trial N-1, and then “even” button for the digit 8 on trial N, repeating the odd/even game. If the participant decided to alternate between game types, participants made a risky decision, e.g., they select “odd” button for the digit 3 on trial N-1, and then “high” button for the digit 8 on trial N, switching to the higher/lower game. Across half of the blocks these instructions were counterbalanced such that switching between games led to the certain decision and repeating the same game would yield the risky decision. In the Results section these block instructions are referred to as switch = risk blocks and “repeat = risk” blocks. In other words, to select a risky decision, participants had to switch between games (switch = risk blocks), while in the other blocks (repeat = risk blocks), participants had to repeat the same game.

A, Rewarded voluntary switch task, a combined risky decision-making and task-switching paradigm. In trial N, subjects may select the certain decision (25 RUB with a probability of 100%) or the risky decision (50 RUB or 0 RUB with a probability of 50%) depending on decision to switch or repeat task sets from trial N-1. Figure represents trial in the reward switch = risk block. B, tACS montage. Active electrodes were placed on F3 and F4 electrode, representing left and right frontal area. Placement of the reference electrode was the ipsilateral deltoid for F3 and F4.
Since gain-framed and loss-framed decisions differentially affect risk preferences (Tversky and Kahneman, 1985), the experiment was also divided into gain and loss blocks. In gain blocks, certain decisions were defined and instructed as “100% probability that you would receive 25 Russian rubles (RUB)”, while risky decisions were defined and instructed as “50% probability that you would receive 50 RUB” (or alternatively 0 RUB). In loss blocks, the certain decision indicated “100% probability that you would lose 25 RUB” while risky decisions indicated “50% probability that you would lose 50 RUB” (alternatively 0 RUB). For each response that determined the game they selected, a feedback screen displayed for 1000 ms indicated the amount of money gained or lost for that particular trial. If response time exceeded 4000 ms or participants responded erroneously, feedback for that particular trial displayed negative feedback (e.g., 0 RUB for gain block, −50 RUB for loss blocks).
Similarly to the voluntary switching task, response buttons were counterbalanced across participants (Arrington and Logan, 2004). Block condition were counterbalanced in random order. Presentation of stimuli and recording of responses were controlled by E-Prime 2.0 software. All text was displayed in black font on a gray scale background and all participants were instructed to use two hands to respond. Due to the difficulty of the task and to avoid learning effects, participants received two rounds of training, which consisted of eight blocks of 10 trials, resulting in 80 trials in total. If accuracy was below 95% additional training sessions were given. This learning phase was reflected in the actual experiment in which accuracy for all participants throughout the task was above 92%. After training, participants received 20 blocks of 20 trials each. At the end of the experiment participants were shown the total cumulative feedback on the computer screen. Participants received 500 RUB for participation (500 RUB ≈ 7 United States dollar) and an additional bonus, between −300 and +300 RUB, based on the feedback outcomes of six randomly selected trials to maintain an equal motivation for risky decision making across blocks (Krajbich et al., 2012).
tACS procedure
By using the international electroencephalography 10-20 system, tACS was applied on the left or right frontal areas by placing a 7 × 5-cm saline-soaked electrode on F3 or F4 locations (Fig. 1B). For both location sites, a reference electrode was placed on the ipsilateral deltoid to the target electrode (Im et al., 2012; Bai et al., 2014). The order of stimuli was randomized across 20 blocks. Standard protocols were employed as in previous frequency-controlled tACS experiments on motor and cognitive tasks (sham, 5, 10, 20, and 40 Hz; Feurra et al., 2011, 2016; Santarnecchi et al., 2013, 2016), accounting for mean center frequencies (Klimesch, 2012). Furthermore, tACS set at a fixed frequency has been shown to entrain individualized α-oscillations converging to a 10-Hz stimulation (Helfrich et al., 2014a). Therefore, we contend that these frequency stimulations suffice to entrain endogenous neural oscillations within standard θ, α, β, and γ ranges, respective of individual frequency ranges.
Stimulation was delivered online during task performance, with exception to sham stimulation, which lasted for 30 s. To implement a sham stimulation, instead of using a fixed frequency that may bias a single stimulation protocol over another, we applied sinusoidal low-frequency transcranial random noise stimulation (tRNS) between 0.1 and 100 Hz for 30 s. This sham stimulation protocol was necessary in the current experiment due to the unconventional use of multiple stimulation protocols reflecting the harmonics of mean center frequencies (Klimesch, 2012). Furthermore, it is important to emphasize that low-frequency tRNS was applied only for a short duration, compared to all other protocols that were applied throughout the entire block; sham stimulation was delivered for 30 s with 10-s fade-in/fade-out, while all other stimulation protocols lasted between 5 and 10 min. Moreover, low-frequency tRNS has been shown not to affect cortical excitability (Paulus, 2011). Stimulation current was set at 1 mA (500 mA peak-to-peak). The maximum current density at the stimulation electrode was ∼14 μA/cm2. The wave form of the stimulation was sinusoidal, and there was no direct current offset. The low intensity of stimulation was used to avoid a perception of flickering lights (Paulus, 2010). Stimulation was delivered using a battery-operated stimulator system (BrainStim, EMS Medical). Impedance was kept below 10 kΩ. All protocols began one minute before each block. Due to abundant evidence that tACS affects physiologic activity during stimulation (Antal et al., 2008; Helfrich et al., 2014a,b; Strüber et al., 2015), breaks of 5 min were given after each set of four blocks. In total, stimulation lasted ∼40 min.
Statistical analysis
Analysis was performed using R software (R Core Team, 2016) with the software package lme4 (Bates et al., 2014) and lmertest (Kuznetsova et al., 2016). Two separate logistic regression mixed models (Generalized Linear Mixed Model) on the raw data were performed on the following variables: (1) selection of risky decisions and (2) selection of switches between trials. Each model included the following categorical predictors: valence (gain, loss blocks), switch condition (switch = risk blocks and repeat = risk), frequency of stimulation (sham, 5, 10, 20, and 40 Hz) with sham as a reference variable, and hemisphere of stimulation (left, right). Before analysis error trials and trials exceeding response time of four seconds were omitted. Wald tests (Kuznetsova et al., 2016) were performed on all levels up to two interactions. To account for possible group differences, sham stimulation was used as a reference variable for each effect associated with frequency. In the logistic regression model participants, valence, switch condition, and frequency of stimulation were modeled with random effects, while hemisphere of stimulation (a between-subjects factor) was modeled with fixed effects. The R command lme4 function is as follows: glmer(Risk ∼ (Frequency + Valence + Hemi) ^ 2 + (1 + Frequency + Condition + Valence:Condition | Subject), family = “binomial”, data = D, control = glmerControl (optimizer=“bobyqa”, optCtrl = list (maxfun = 2e5))). Significance for the regression coefficients was corrected for false positives by using Holm-Bonferroni procedure.
For the following analyses we used SPSS software version 20 (IBM Corp, 2011). A mixed ANOVA was performed on the mean response time of the following variables: valence (gain, loss blocks), frequency of stimulation (sham, 5, 10, 20, and 40 Hz), switch condition (switch = risk blocks and repeat = risk blocks), and hemisphere of stimulation (left- and right-stimulated group), in which switch condition, valence, and frequency of stimulation were within-participants factors and hemisphere of stimulation was treated as between participants factor. Sphericity was not violated across any of these effects (all p > 0.05). To assess whether participants selected more risky decisions than chance level, a one-sample t test was performed.
Results
Figure 2 displays the percentage of risky decisions in all stimulation conditions. The logistic regression mixed model for risky decision making revealed an increase in risky decision making during 20 Hz of stimulation particularly when stimulating the left PFC (β = 0.989; p = 0.00194; p’ = 0.043). The effects of other tACS frequencies on risky decision making did not survive Holm-Bonferroni correction for multiple comparisons (Table 1). The frequency- and hemisphere-specific effect of a 20-Hz stimulation was confirmed by a nonsignificant main effect of hemisphere of stimulation (β = 0.072, p = 0.885; p’ > 0.999). Figure 2 displays means and standard error for each of the comparisons with regards to the frequency of stimulation × hemisphere of stimulation interaction effect.

Mean percentage of risky decisions for each tACS condition with respect to sham; 20-Hz stimulation of the left frontal area increased selection of voluntary risky decisions. Error bars correspond to SEM.
Results of the logistic regression for risky decision making and for voluntary switching
In addition, separate logistic regression models were performed using sham as a reference for each stimulation group (Table 2). The model for the left-stimulated group (Table 2) revealed a statistical significant increase in risky decision making from a 20-Hz stimulation (β = 0.610, p = 0.001; p’ > 0.021). Follow-up analysis using 20 Hz as a reference (Table 3) revealed that 20 Hz of stimulation applied to the left hemisphere increased risky decision making with respect to sham (β = −0.989, p = 0.001; p’ > 0.021) and 40-Hz stimulation (β = −1.265, p < 0.001; p’ > 0.015). When separately testing hemisphere stimulation groups with 20 Hz as the reference variable, 20 Hz increased risky decision making compared to sham for the left-stimulated group (β = −0.610, p = 0.001; p’ > 0.021; Table 4). No effects were found for the right-stimulated group (Table 4).
Logistic regression model of risky decision making for each group with sham as a reference variable
Logistic regression model of risky decision making with 20 Hz as a reference variable
Logistic regression model of risky decision making for each group with 20 Hz as a reference variable
After Holm-Bonferroni correction, the logistic regression mixed model yielded no significant effects of tACS on voluntary switching, yet revealed a main effect of switch condition (β = 2.005; p ≤ 2 × 10−16; p’ < 0.001), an interaction effect of switch condition × valence (β = −0.311; p = 4.47 × 10−5; p’ = 0.001), and an interaction effect of switch condition × hemisphere of stimulation (β = 0.908; p ≤ 2 × 10−16; p’ < 0.001). These effects indicate an increase in voluntary switching in the switch = risk blocks compared to repeat = risk blocks, especially for loss blocks; perhaps reflecting an influence of executive control on the framing bias. The interaction effect of switch condition and Hemisphere may demonstrate an increase in voluntary switching during switch = risk blocks compared to repeat = risk blocks from the left-stimulated group, yet should be interpreted with caution since Hemisphere of stimulation was modeled with fixed effects (see Discussion for details). Analysis of response times revealed that participants responded more slowly in trials in which switching between tasks led to risk (μ = 1112.43 ms) compared to trials in which repeating led to risk (μ = 988.98 ms; F(1,32) = 17.455; p < 0.001, partial η2 = 0.353). Since participants overall were more likely to select risky decisions (μ = 63.6%; SE = 0.004; one-sample t test: t = 33.037; p < 0.001), we infer that this observed difference in response time is likely due to switching costs (for a detailed account on the voluntary switch cost, see Arrington et al., 2014).
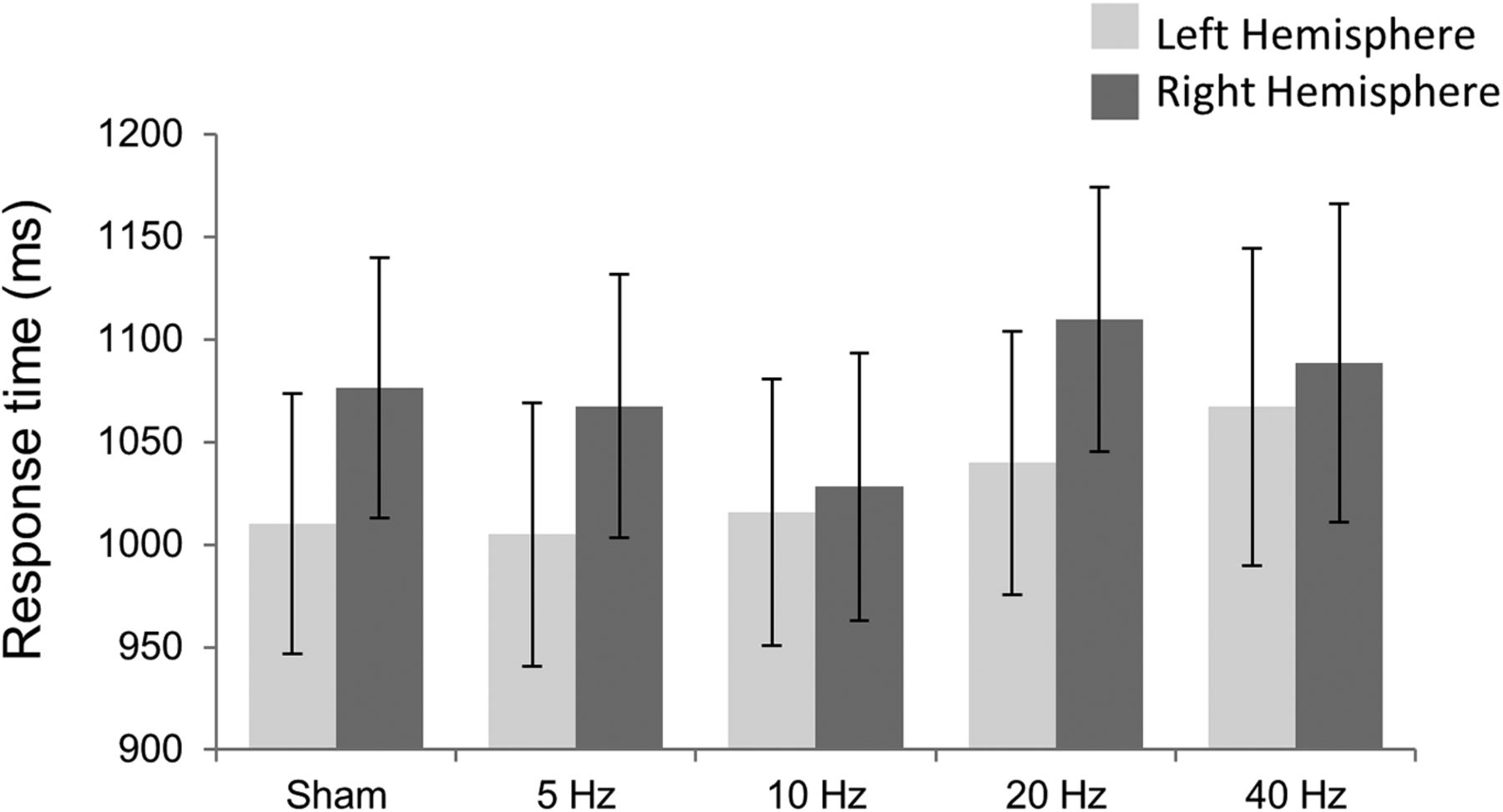
In addition, the mixed ANOVA on response time revealed a main effect of valence (F(1,32) = 25.842; p < 0.001, partial η2 = 0.447), showing slower mean response times in loss blocks (μ = 1085.96 ms) compared to gain blocks (μ = 1015.44 ms). This significant difference may indicate increased deliberation in loss blocks. No other effects on reaction times were significant. See Figure 3 and Table 5 for list of response times for each condition.

Mean response times for each Hemisphere group across frequency stimulation.
Mean response times associated with each hemisphere group across frequency stimulation
Discussion
In the attempt to modulate oscillatory activity underling voluntary risky decision making and executive control we applied tACS (sham, 5, 10, 20, and 40 Hz) to the left and right PFC while participants performed a modified risky decision-making task that requires choosing between risky and certain decisions by switching or repeating task sets. The analyses of risky decision making revealed several significant effects, yet the influence of a 20-Hz stimulation on risky decision making was the most robust, surviving Holm-Bonferroni correction. Although frequency specificity has been demonstrated with 20-Hz tACS for motor (Pogosyan et al., 2009; Feurra et al., 2011, Joundi et al., 2012) and sensory functions (Kanai et al., 2008, 2010; Turi et al., 2013), the current experiment is the first to reveal a frequency-specific increase in voluntary risky decision making from 20-Hz tACS.
Within recent years, EEG studies investigating oscillatory activity in gambling tasks have demonstrated a correspondence between frontal β-oscillations (20–35 Hz) and anticipation of probable rewards (Bunzeck et al., 2011), as well as receiving unexpected rewarded feedback (Marco-Pallares et al., 2008; HajiHosseini et al., 2012; HajiHosseini and Holroyd, 2015; Mas-Herrero et al., 2015). Marco-Pallarés et al. (2015) proposed that frontal β-oscillatory activity during gambling paradigms might signify the functional coupling between cortical and subcortical regions such as the ventral striatum, known to be involved in reward processing. This was recently confirmed in an EEG-fMRI study that reported correspondence between mid-frontal β-oscillatory activity and engagement of the fronto-striatal-hippocampal network (Mas-Herrero et al., 2015). This may indicate that 20 Hz of stimulation increased motivation to select risky decisions by indirectly affecting brain regions of the reward system, such as the ventral striatum. Importantly, the ventral striatum is a key subcortical region for risky decision making since the activation of this area predicts risky decision making and increases in activation as rewards become more probable (Knutson and Greer, 2008; Niv et al., 2012). Taken together, we speculate that stimulation of the frontal cortex with 20-Hz tACS may have resulted in a boost in reward-related processes involving the ventral striatum, thus resulting to an increase in voluntary risky decision making. Further support for this claim derives from electrical simulations of the left PFC (F3, EEG 10–20 system) with an extracephalic electrode placed on the shoulder demonstrating modulation of the PFC and deep medial structures (Bai et al., 2014).
It is important to underline that although several effects involving hemisphere of stimulation were statistically significant, these effects should be generalized to the population cautiously since we used a between group design combining random effects (valence, switch condition, and frequency of stimulation) with fixed effects (Hemisphere of stimulation). Importantly, the specific effect of 20-Hz tACS of the left PFC on risky decision making was further confirmed by separate statistical analyses for the left- and right-side stimulation (Table 2). Another potential caveat to the study is that potential after-effects of tACS cannot be ruled out as no simultaneous EEG recording took place. Despite the growing evidence that tACS effects neural oscillatory activity online (Antal et al., 2008; Helfrich et al., 2014a,b; Strüber et al., 2015), it was not possible to control within the current experiment.
The results of the current study seem contradictory to a previous study using tACS on risky decision making (Sela et al., 2012). However, the effect of θ-band tACS in the previous study (Sela et al., 2012) could be due to a modulation of feedback-related adjustments (Cavanagh et al., 2010; Cavanagh et al., 2012; Luft, 2014; Zhang et al., 2014) since the previous tACS paper used the Balloon Analog Risk Task, which measures risk-taking propensity across a cumulative number of responses, as opposed to measuring risky decision making within a single response, as in the current study. A possible explanation for the alternate results may be due to the differences in montage. For instance, a previous study that modulated executive functions, specifically working memory, stimulated both frontal and parietal areas using an F3–P3 montage (Polanía et al., 2012). We suggest that stimulation of the frontal lobe may modulate either a frontal-striatal network associated with voluntary risky decision making (Rao et al., 2008) or a frontal-parietal network in association with voluntary executive control (Orr and Banich, 2014) depending on the placement of the reference electrode (Bai et al., 2014). Whereas the F3–EC (extracephalic) montage used in the current study likely modulates frontal and deep medial structures, an F3–P3 montage likely modulates frontal and parietal structures (Bai et al., 2014). Therefore, modulation of voluntary executive control may require an F3–P3 montage. Some have reported that the ratio of θ- and β-oscillations at resting state can be used to predict risk preferences in individuals (Schutter and van Honk, 2005; Massar et al., 2014). Therefore, it is also plausible that both θ- and β-band stimulation may modulate different cognitive components of the decision-making process within different states and/or contexts. Alternatively, one may suggest that the 20-Hz stimulation could modulate working memory during risky decision making. Some studies suggest that risky decision making is associated with the capacity to maintain and organize information in working memory as an estimation of executive processes (Brevers et al., 2014a,b). Unfortunately, our study design did not allow testing of this hypothesis. However, we think that a modulation of working memory should not affect our results since subjects continued to receive training until their performance became above 95%, as specified in the stimuli and procedure section, thereby eliminating potential confound learning effects and an overload of working memory.
Finally, our findings are consistent with the previous studies demonstrating that laterality (left and right frontal hemisphere) strongly influences the effect of voluntary risky decision making (Knoch et al., 2006; Fecteau et al., 2007a,b; Sela et al., 2012; Cheng and Lee, 2016). Together, these previous studies show that exciting the left and/or inhibiting the right PFC increasing risky decision making and vice versa. This suggests that a 20-Hz stimulation increases cortical excitability of the left frontal area, presumably by entraining the frontal-striatal network. Together these results offer novel insight into the role of β-oscillatory activity in neural mechanisms of risky decision making.
Acknowledgments
Acknowledgements: We thank Bhuvanesh Awasthi for his recommendations and expertise in task programming and for data collection.
Footnotes
The authors declare no competing financial interests.
The work was supported by the Russian Academic Excellence Project 5-100.
This is an open-access article distributed under the terms of the Creative Commons Attribution 4.0 International license, which permits unrestricted use, distribution and reproduction in any medium provided that the original work is properly attributed.
References
In this issue

{kind=link}
{kind=link}
{kind=link}



