

Article Figures & Data
Figures
- Figure 1.
TA-induced LTP-D is blocked by the DA receptor antagonist, clozapine. A, The left graph shows the time course of changes in EPSPs following SC HFS (arrow) and depotentiation by TA/perforant path LFS (PLFS, bar). Note that data in A include control slices done in our prior manuscript (Izumi and Zorumski, 2016) with additional slices added. The right graph in A depicts an analysis from a single slice based on changes in the IO curve. Black circles are baseline IO results, while red circles depict changes 60 min after SC HFS, and red squares show reversal of LTP following PLFS. B, The ability of PLFS to depotentiate SC LTP is completely blocked by 1 μM clozapine (white bar). Upper traces to the right show representative EPSPs at baseline (black lines) and 60 min following SC HFS (redlines) while lower traces show baseline (black lines) compared to 60 min following PLFS (red lines). Calibration: 1 mV, 5 ms.
- Figure 2.
TA-induced LTP-D involves D4Rs. A, The graph shows the inability of a selective D2R antagonist (0.2 μM L-741,626) to block PLFS induced depotentiation of SC LTP. SC HFS was delivered at the arrow; PLFS was administered during the hatched bar. B, In contrast, a selective D4R antagonist (0.1 μM L-745,870) completely inhibited depotentiation. Traces to the right show representative EPSPs as in Figure 1. Calibration: 1 mV, 5 ms.
- Figure 3.
A D4R antagonist blocks homosynaptic SC depotentiation. A, The graph shows the ability of SC LFS [SLFS (1 Hz × 15 min), hatched bar] to depotentiate previously established SC LTP. SC HFS was administered at the arrow. B, The D4R antagonist, 0.1 μM L-745,870, blocked homosynaptic SC depotentiation. For reasons that are uncertain, we observed an increase in variance of EPSPs during perfusion of the D4R antagonist in this set of studies but not in the studies shown in Figure 2. Traces to the right show representative EPSPs as in Figure 1. Calibration: 1 mV, 5 ms.
- Figure 4.
Exogenously administered NRG1β, but not a D4R agonist, depotentiates SC LTP. A, A selective D4R agonist,0.2 μM PD-168,077 (black bar), failed to depotentiate SC LTP when administered for 15 min 60 min following LTP induction. SC HFS was administered at the arrow. B, In contrast to the D4R agonist, 1 nM NRG1β (black bar) induced chemical depotentiation of SC LTP. Traces to the right show EPSPs as in Figure 1. Calibration: 1 mV, 5 ms.
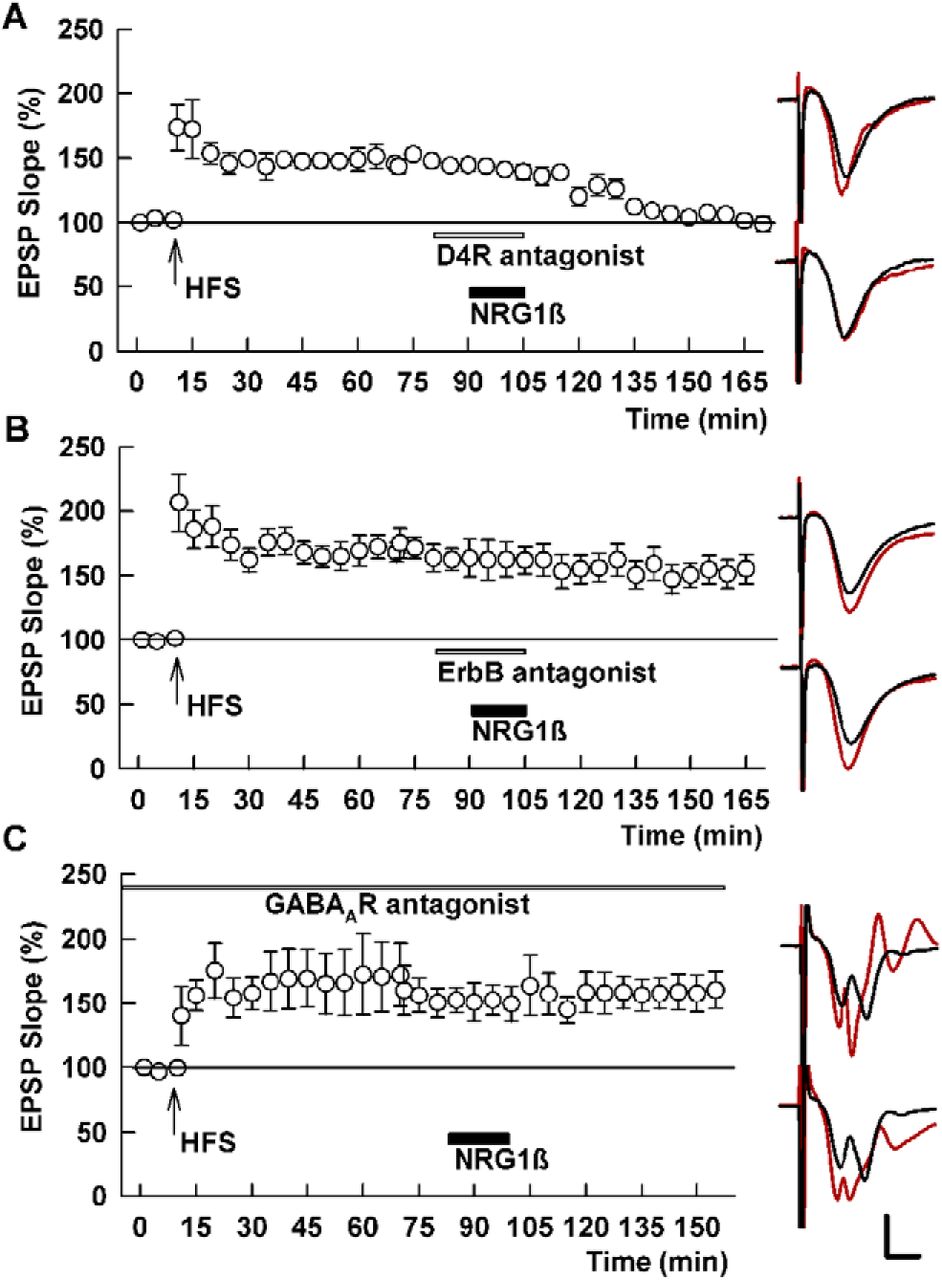
- Figure 5.
Depotentiation by NRG1β is insensitive to D4R antagonism but blocked by an ErbB antagonist and PTX. A, In the presence of 0.1 μM L-745,870 (white bar), 1 nM NRG1β (black bar) induces SC depotentiation. SC HFS was administered at the arrow. B, C, In contrast to the D4R antagonist, a pan-ErbB antagonist (10 μM PD-158,780, white bar) blocks NRG1β-induced depotentiation (B), as does the GABAAR antagonist, 1 μM PTX (C). Traces show representative EPSPs as in Figure 1. Calibration: 1 mV, 5 ms.
- Figure 6.
An ErbB antagonist blocks TA-induced, but not homosynaptic SC depotentiation. A, In the presence of 10 μM PD-158,780, TA stimulation (PLFS, hatched bar) fails to induce persistent SC depotentiation. SC HFS was administered at the arrow. B, In contrast, the ErbB antagonist fails to block depotentiation induced by homosynaptic SC LFS (SLFS, hatched bar). Traces show representative EPSPs as in Figure 1. Calibration: 1 mV, 5 ms.
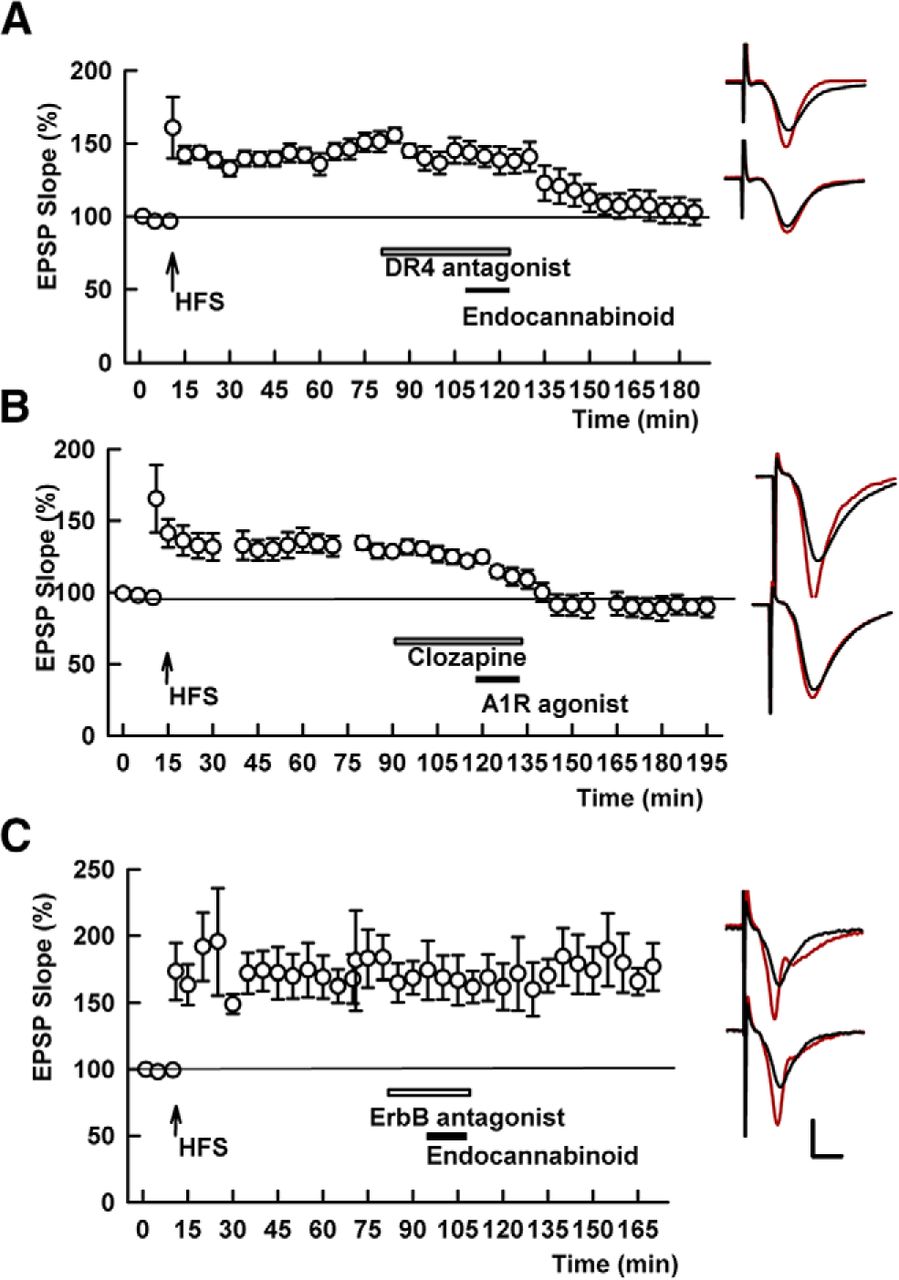
- Figure 7.
D4R antagonism does not block chemical depotentiation by CB1R or adenosine A1R activation, but ErbB antagonism blocks the effects of a CB1R agonist. A, In the presence of the D4R antagonist, L-745,870 (white bar), the endocannabinoid, 20 μM 2-AG (black bar), reversed SC LTP. SC HFS was administered at the arrow. B, Similarly, clozapine, a DAR antagonist with selectivity for D4Rs (white bar), failed to block depotentiation by 10 nM CPA, a selective A1R agonist (black bar). C, The effects of 2AG on SC STP were blocked by the ErbB antagonist, PD-158,780. Traces show representative EPSPs as in Figure 1. Calibration: 1 mV, 5 ms.
- Figure 8.
The diagram depicts a current scheme for TA-induced SC LTP-D based on prior studies (Izumi and Zorumski, 2008, 2016) and the present results. Agents that promote chemical depotentiation are shown in green while agents that inhibit TA-induced SC depotentiation are shown in red.






In this issue

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}



