

Abstract
The olfactory cortex is part of the mammalian cerebral cortex together with the neocortex and the hippocampus. It receives direct input from the olfactory bulbs and participates in odor discrimination, association, and learning (Bekkers and Suzuki, 2013). It is thought to be an evolutionarily conserved paleocortex, which shares common characteristics with the three-layered general cortex of reptiles (Aboitiz et al., 2002). The olfactory cortex has been studied as a “simple model” to address sensory processing, though little is known about its precise cell origin, diversity, and identity. While the development and the cellular diversity of the six-layered neocortex are increasingly understood, the olfactory cortex remains poorly documented in these aspects. Here is a review of current knowledge of the development and organization of the olfactory cortex, keeping the analogy with those of the neocortex. The comparison of olfactory cortex and neocortex will allow the opening of evolutionary perspectives on cortical development.
Significance Statement
The olfactory cortex is an evolutionarily conserved paleocortex implicated in odor processing. While several studies addressed how the olfactory cortex encodes and processes odorant information, little is known about its precise cellular origin, diversity, and identity. Unraveling where the cells are born and how they migrate toward and settle the olfactory cortex during development is of main importance in understanding its circuit organization and function. In addition, comparing the development of the olfactory cortex with that of the neocortex will help in identifying common evolutionarily conserved developmental mechanisms as well as new mechanisms specific to the neocortex that appeared later during evolution and participate in building the mammalian cortex.
Introduction
Mammalian cerebral cortex comprises the neocortex, the hippocampus, and the olfactory cortex. The olfactory cortex is an evolutionarily conserved paleocortex located in the ventrolateral part of the telencephalon and shares common characteristics with the general cortex of reptiles, which are composed of three thin layers (Aboitiz et al., 2002; Shepherd, 2011; Fournier et al., 2015). This three-layered organization, also called allocortex, is conserved in both the hippocampus (medial pallium derivative) and the olfactory cortex (lateral pallium derivative; Aboitiz et al., 2002). The mammalian olfactory cortex is subdivided into several structures along anteroposterior axis, as follows: the anterior olfactory nucleus, the olfactory tubercle, the piriform cortex, the olfactory amygdala [cortical amygdala and nucleus of the lateral olfactory tract (LOT)], and the lateral entorhinal cortex (lENT; Haberly, 2001; Brunjes et al., 2005; Sanchez-Andrade and Kendrick, 2009). In contrast to the classic sensory pathway where sensory information is first relayed in the thalamus before reaching the neocortex, the olfactory cortex receives direct sensory inputs from the mitral and tufted cells of olfactory bulb (OB) via the LOT, and is thought to play a key role in olfaction (Wilson and Mainen, 2006; Isaacson, 2010; Leinwand and Chalasani, 2011; Bekkers and Suzuki, 2013). The piriform cortex is the most studied structure of the olfactory cortex. It receives inputs from the OB, as well as from the other regions of the olfactory cortex cited above, and sends projections to the anterior olfactory nucleus, the olfactory tubercle, the cortical amygdala (CoA), and the lENT within the olfactory cortex, as well as to the mediodorsal nucleus of the thalamus and to several subdivisions of the prefrontal cortex [including the infralimbic (IL), orbitofrontal, and agranual insular cortices; Johnson et al., 2000; Ekstrand et al., 2001; Meyer et al., 2006; Kerr et al., 2007]. Finally, the piriform cortex sends consequent feedback projections to the OB (Haberly and Price, 1978; Shipley and Adamek, 1984). The piriform cortex has been shown to participate in odor discrimination, association, and learning (Bekkers and Suzuki, 2013); it allows object recognition in a sensory landscape, whose relevant perceptual dimensions are dynamically shaped by sensory experience (Fournier et al., 2015).
While the development of the six-layered neocortex has been extensively described, the development of the olfactory cortex remains poorly understood. Neurons of the olfactory cortex seem to have multiple origins both in the pallial and the subpallial ventricular zones (VZs; Carney et al., 2006; Garcia-Moreno et al., 2008). Moreover, these neurons migrate over long distances and display complex migratory properties before reaching their final destination in the olfactory cortex and organizing into three layers. In this review, we discuss current knowledge on the development and organization of the olfactory cortex and investigate the following questions. How is the olfactory cortex specified during development? Where are the olfactory cortex neurons born? How do they migrate to the olfactory cortex and form the three layers? What is known about neuron diversity within the olfactory cortex, and how do they encode olfactory information? These questions will be addressed keeping the analogy with the neocortex development in order to open evolutionary perspectives on cortical development.
Arealization and neurogenesis
During the first 10 days of mouse embryonic development, the pallial neuroepithelium proliferates, allowing the exponential generation of progenies through symmetric divisions in the VZ (Noctor et al., 2004). From embryonic day 11 (E11), the first postmitotic neurons are produced by asymmetric divisions, generating both progenies, which stay in the VZ, and neuroblasts, which migrate out of the VZ toward the cortical surface.
Arealization of the cortex by signaling centers
Based on gene expression, the pallium is subdivided into four major subregions, namely the medial, dorsal, lateral, and ventral pallium; each pallial region is thought to give rise to specific cortical structures, such as the hippocampus, the neocortex, the olfactory cortex, and the amygdala/endopiriform cortex nuclei, respectively (Puelles et al., 2000; Yun et al., 2001). Intrinsic mechanisms based on morphogens and signaling molecules secreted by patterning centers allow the graded expression of transcription factors by cortical progenitors and the development of the different cortical fields (O'Leary et al., 2007; Arai and Pierani, 2014). The following three patterning centers lie at the borders of the telencephalon and participate in the arealization of the cortex: the cortical hem (between cortical and choroidal fields); the commissural plate (at the rostromedial pole of the telencephalon); and the cortical antihem (at the pallial–subpallial boundary, PSB; Mallamaci and Stoykova, 2006). The LIM homeobox protein 2 (LHX2), expressed in a rostrolateralHIGH to caudomedialLOW gradient, suppresses hem and antihem fates, both of which expanded in Lhx2 mutant mice (Bulchand et al., 2001; Monuki et al., 2001; Nakagawa and O'Leary, 2001; Mangale et al., 2008). The transcription factor FOXG1, expressed in a rostrolateralHIGH to caudomedialLOW gradient, suppresses hem fate and is required for lateral fates, including that of the antihem (Dou et al., 1999; Muzio and Mallamaci, 2005; Hanashima et al., 2007; Shibata et al., 2008). The cortical antihem is of major importance for the specification of the olfactory cortex. In the following section, we will discuss the antihem specification and functions during development (see O'Leary et al., 2007; Subramanian et al., 2009; Montiel and Aboitiz, 2015 for further information about organizing centers and arealization of the cerebral cortex).
The antihem, major signaling center for the determination of the olfactory cortex
The antihem is located at the PSB, between the ventral pallium and the dorsal lateral ganglionic eminence (dLGE; Yun et al., 2001). The antihem is delineated from adjacent regions through the exclusive expression of the transcription factor Dbx1 in the ventral pallium VZ, as well as the enriched expression of the secreted Frizzled related gene sFrp2 (Kim et al., 2001; Yun et al., 2001; Assimacopoulos et al., 2003; Medina et al., 2004). The ventral pallium and the adjacent dLGE both display an enriched expression of Pax6 transcription factor, which is required for the development of lateral and ventral pallial identities (olfactory cortex and amygdaloid complex; Stoykova et al., 1996; Toresson et al., 2000; Kim et al., 2001; Yun et al., 2001; Hirata et al., 2002; Tole, 2005; Piñon et al., 2008; Cocas et al., 2011).
The position and the specification of the antihem rely on the expression of transcription factors Pax6, Tlx, and Gsh2. In Pax6 mutant mice, the ventral and the lateral pallium ectopically express subpallial markers, such as Mash1, Gsh2, and Dlx2 (Stoykova et al., 1996, 1997, 2000; Toresson et al., 2000; Kim et al., 2001; Yun et al., 2001). Tlx mutants display a similar but less dramatic phenotype (Stenman, 2003). In line with Gsh2 expression in the subpallium, Gsh2 mutants display pallial gene expression in subpallial domains, such as the dLGE (Toresson et al., 2000; Yun et al., 2001). PAX6 is therefore required for the expression of ventral pallium-specific genes, while GSH2 suppresses their expression in the subpallium (Carney et al., 2009).
The organizer function of the antihem is poorly understood compared with that of the cortical hem in hippocampus development (Subramanian et al., 2009). A prominent palisade of radial glial fibers delineates the PSB. These fibers originate in the corticostriatal junction of the VZ and extend up to the pial surface in the piriform cortex (Molnár and Butler, 2002). In the absence of the antihem or in both Pax6 and Tlx mutants, the radial glial palisade is severely affected at the PSB. Moreover, Pax6 mutants show a higher number of subpallium-derived interneurons in the cortex, suggesting a role of the radial glial palisade in restricting tangential migration of interneurons during development (Chapouton et al., 1999). In addition, while the antihem expresses Wnt signaling inhibitor sFrp2, Wnt7b is expressed in the dLGE adjacent to the antihem (Kim et al., 2001; Assimacopoulos et al., 2003). This restriction of the Wnt signaling to the subpallial side of the PSB instructs the position of the radial glial palisade. The antihem further expresses specific molecules, like the epidermal growth factor family members TGF-α, Neuregulin 1 (NRG1) and NRG3, and fibroblast growth factor 7 (Kim et al., 2001; Assimacopoulos et al., 2003). NRG1 has been shown to be essential in the formation and maintenance of the radial glial cells (Anton et al., 1997; Schmid et al., 2003).
Signaling centers and production of different lineages of pioneer Cajal-Retzius cells
The three organizing centers at the pallial borders are known to be the main origins of Cajal-Retzius (CR) cells (Takiguchi-Hayashi, 2004; Bielle et al., 2005; Yoshida, 2006; Zhao et al., 2006). These early-born pallial cells differentiate between E10.5 and E12.5 in mice and populate the marginal zones of all cortical areas (Smart and Smart, 1977; Wood et al., 1992; Marín-Padilla, 1998). Many CR cells express Emx1 and Tbr1 pallial transcription factors (Gorski et al., 2002; Hevner et al., 2003), and Reelin, a secreted glycoprotein that guides radial migration of neocortical neurons (Caviness, 1982; Howell et al., 1997; Alcántara et al., 1998). All CR cells express Reelin, but only hem lineage-derived CR cells express the tumor protein 73 (p73), while antihem- or septum-derived CR cells specifically express Dbx1. This diversity of CR cell lineages may play a role in the development of cytoarchitectonic differences between the neocortex, the olfactory cortex, and the hippocampus (Bielle et al., 2005).
Migration and layer formation
Migration and layer formation in the neocortex
Neuron migration in the neocortex has been well studied. From E11, the first neuroblasts accumulate superficially in the neocortical neuroepithelium, forming the preplate beneath the CR cell layer (Marin-Padilla, 1978; Stewart and Pearlman, 1987; Narboux-Nême and Gaspar, 2008; Vitalis and Rossier, 2011). The preplate is a transitory developmental structure in the neocortex of mouse embryos. Axons of preplate neurons are thought to play the role of pioneers for the development of several fiber tracts (Supèr et al., 1998). From E13, new excitatory neurons settle the preplate, dividing it in two layers, the marginal zone at the surface and the subplate near the VZ. Simultaneously, the radial glia develops and its processes reach the pial surface (Rakic, 2003; Borrell and Götz, 2014). These radial glia processes allow the subsequent migration of neuroblasts, which invade the cortical plate in between the marginal zone and the subplate from E13 to E16 (Marin-Padilla, 1978; De Carlos and O’Leary, 1992). Excitatory neurons of cortical layers are then produced in an inside-out manner from the deep to the superficial layers, such that late-born neurons migrate throughout early-born neurons (Molyneaux et al., 2007; Rakic et al., 2009). Interneurons of the neocortex are produced in the ganglionic eminences of the subpallium [medial ganglionic eminence (MGE), LGE, and caudal ganglionic eminence; Anderson et al., 1997] and migrate first tangentially and then radially to settle the cortical layers, starting from E13 (see Fig. 2; Marín and Rubenstein, 2001; Métin et al., 2006; Gelman et al., 2011). MGE-derived interneurons born at different stages settle distinct layers of the neocortex (Miyoshi and Fishell, 2011).
Early migration to the olfactory cortex
In contrast to the neocortex, the olfactory cortex is located ventrolaterally and therefore does not line the pallial VZ. Complex waves of cell migration from various regions of the VZ toward the olfactory cortex have been described in early mouse development, from E9.75 to E12 (Nomura et al., 2006; Garcia-Moreno et al., 2008). Olfactory neurons generated in the dorsal part of the telencephalon at E9.75 migrate ventrally and align the PSB through Ephrin-A5 repulsive activity, which expression is regulated by PAX6 transcription factor. These ventrally migrating neurons differentiate both in lot cells and olfactory cortex neurons (Nomura et al., 2006).
Lot cells are a specific subpopulation of Reelin+/p73+ CR cells born from E9.5 to E11.5, which were named after their specific expression of lot1 (mGluR1β; note that lot cells are distinguished from LOT axons by small and capital letters, respectively; Sato et al., 1998; Jiménez et al., 2002; Dixit et al., 2014). Proneural genes Neurogenin1 and Neurogenin2 are coexpressed in dorsal and ventral pallial progenitors and required for the differentiation of lot cells (Dixit et al., 2014). These cells are produced in the whole VZ of the pallium from E9.5 to E11.5 and next migrate tangentially and ventrally toward the cortex surface to organize themselves around the presumptive territory of the LOT at E12.5 (Tomioka et al., 2000). The migration and position of lot cells rely on the expression of guidance cues. The neocortex displays dorsoventral gradients of cues, which position the first steps of lot cell migration. The subpallium expresses repulsive cues, which prevent lot cells from invading ventral territories and maintain them superficially at the PSB. Among these cues, Netrin-1 has been shown to locally attract lot cells around the PSB, while Semaphorin 3F is secreted by deep regions of the ventral telencephalon and keeps lot cells at the cerebral surface by its repulsive action (Kawasaki, 2006; Ito et al., 2008). Lot cells act as guideposts for the establishment and positioning of LOT axons, which occur from E12.5 to E13.5 in mice (Squarzoni, 2015). At these early stages, lot cells display long processes and respond to electric stimulation of the OB (Sato et al., 1998; Hirata et al., 2012). Except for lot cells, a general role of CR cells as guideposts for axon development still remains to be elucidated, but CR cells have been involved in the guidance of entorhinal and thalamocortical axons (Ceranik et al., 2000; Del Río et al., 2002; Barber et al., 2015). Recently, de Frutos et al. (2016) discovered a later and unexpected role for lot cells during development: between E13.5 and E15.5, lot cells retract their processes, adopt a rounded-up morphology with small filopodia, and initiate amoeboid-like migration away from the LOT territory to dorsal neocortical regions (de Frutos et al., 2016). This results in a doubled density of CR cells in the neocortex and a 60% reduction of lot CR cell density in the LOT territory. This reallocation of lot CR cells is regulated by the NMDA glutamate receptor and is required for the proper organization of neocortex layer 1 and for layer 2/3 pyramidal cell apical dendrite morphology and spine density (de Frutos et al., 2016). Since the olfactory cortex predates the emergence of the neocortex in evolution, the lot cell reallocation into the neocortex could reflect an evolutionary co-option of these ancestral guideposts. In the neocortex, CR cells are progressively eliminated by apoptosis and disappear by the end of the second postnatal week (Del Río et al., 1996). It is yet unclear whether lot cells remaining in the olfactory cortex are integrated in the olfactory network or whether they are a transient population as well.
Garcia-Moreno et al. (2008) described other migration maps and destination areas of olfactory cortex cells by DiI injections at E10 to E12 followed by in toto embryo culture for 1 day (Fig. 1). Neuroblasts are produced in the pallial VZ of the dorsal telencephalon and of the rostromedial telencephalic wall, as well as in the subpallial VZ of the lateral ganglionic eminence and of the septoeminential sulcus. These neuroblasts display tangential or radial migration, with some cells displaying both types of migration sequentially. The authors showed that the cell destination and identity in the olfactory cortex differ depending on their origin in the VZ (Fig. 1). Surprisingly, some neurons from the pallial rostromedial telencephalic wall VZ differentiate in interneurons, while others generated in the dorsal telencephalon differentiate in excitatory neurons. In addition, olfactory cortex neurons from the subpallial dorsal lateral ganglionic eminence VZ differentiate in excitatory neurons, while others generated in the septoeminential sulcus mainly differentiate in interneurons (Garcia-Moreno et al., 2008; Fig. 1). In which olfactory cortex layer these early cells do settle and what are their specific functions in the circuit are not yet elucidated. Further investigations are needed to decipher whether cells coming from different regions in the VZ express specific lineage markers, which could help to identify cell populations and their specific functions within the olfactory cortex.

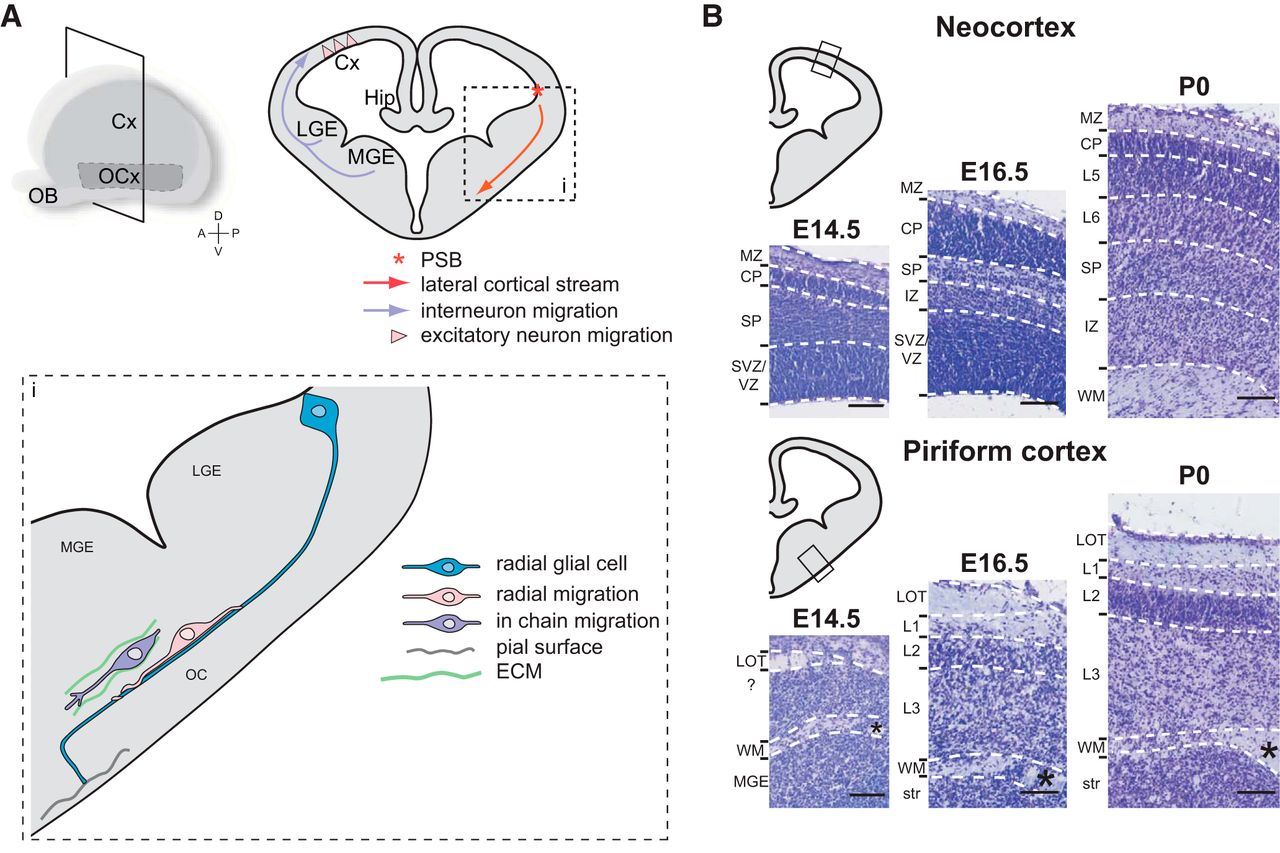
Early migration routes to the olfactory cortex (E10–E12). OCx, Olfactory cortex; PC, piriform cortex; OT, olfactory tubercle; vLGE, ventrolateral ganglionic eminence; SES, septoeminential sulcus; RDT, rostromedial telencephalon; MDT, mediodorsal telencephalon; CDT, caudodorsal telencephalon; RMTW, rostromedial telencephalic wall. In blue, OT migrating neurons; in red, PC migrating neurons. Adapted from Garcia-Moreno et al., 2008, with permission.
Late migration routes to the olfactory cortex
The olfactory cortex neurons born in the lateral and ventral pallium at embryonic stages E12 to E15 in rat embryos (corresponding to E11 to E14 in mice) have been initially described to migrate radially toward the surface of the ventrolateral telencephalon (Bayer, 1986; Valverde and Santacana, 1994; De Carlos et al., 1996). The most described migratory route for olfactory cortex neurons is the lateral cortical stream. This migratory route is formed by cells migrating to the piriform cortex, the olfactory tubercle, and the olfactory amygdala. Neuroblasts taking this route are born in the VZ at the PSB and migrate tangentially to the brain surface through ventral regions of the telencephalon, and then radially to settle in the piriform cortex (Fig. 2). In Pax6 mutant mice (which display an impaired PSB formation), the routing of the lateral cortical stream toward the amygdala and the olfactory cortex is altered (Chapouton et al., 1999; Tole, 2005). In contrast to other known tangential migratory streams described for interneuron migration, the neuroblasts of the lateral cortical stream mainly differentiate into excitatory neurons in the piriform cortex (Corbin et al., 2001). Bai et al. (2008) studied this migratory stream in rat embryos by in utero electroporation of a plasmid, allowing monomeric RFP expression by the cells born at E13 in the VZ at the level of the PSB. After 3 days, the cells migrate tangentially toward ventral regions along the lateral cortical stream. These cells settle the piriform cortex after 6 days and mainly differentiate into neuronal transcription factor TBR1+ cells (Hevner et al., 2001; Bai et al., 2008). However Carney et al. (2006) showed that the identity of cells migrating along the lateral cortical stream is not homogeneous. The authors described two populations of progenies migrating along the lateral cortical stream in mouse embryos: PAX6+ progenies, which derive from the pallium; and DLX2+ progenies, which derive from the subpallium. These two populations are generated at precise and distinct temporal windows during development. From E11.5, Pax6 is highly expressed in the pallium and at the PSB of the VZ. At this stage, some PAX6+ cells are visible along the lateral cortical stream and in the piriform cortex. At E15.5, PAX6+ cells settle the piriform cortex and the amygdaloid complex. DLX2+ cells arose from the PSB about 2 days after PAX6+ cells, at E13.5. DLX2+ cells are actively migrating from E15.5, and by E18.5 they accumulate in the piriform cortex and the olfactory amygdala. PAX6+ cells display a migration along GFAP+ radial glial processes found between the PSB of the VZ and the pial surface of the ventrolateral telencephalon at E13.5 and E15.5 (Carney et al., 2006; Fig. 2). Paradoxically, these cells display a tangential migration along radial glial processes. Some DLX2+ cells display migration along radial glial processes as well, but “in chain” migration has also been observed in this population (Carney et al., 2006; Fig. 2). The authors hypothesize that PAX6+ cells differentiate in excitatory neurons, while DLX2+ cells differentiate in inhibitory neurons, since DLX2+ cells express the interneuron marker Calbindin (Ghanem et al., 2007).

Comparison of developing neocortex and piriform cortex. A, Main migratory routes to the neocortex and to the piriform cortex. i, Lateral cortical stream and modes of migration to the piriform cortex. B, Prenatal development of layers in the neocortex and in the piriform cortex illustrated by cresyl violet stainings. Cx, neocortex; OCx, olfactory cortex; Hip, hippocampus; ECM, extracellular matrix; MZ, marginal zone; CP, cortical plate; SP, subplate; SVZ, subventricular zone; IZ, intermediate zone; WM, white matter; L, layer; str, striatum; *anterior commissure. Scale bars, 200 µm.
Zhao et al. (2008) unraveled novel diencephalon-to-telencephalon migrations into the septum, but also into the piriform cortex and the amygdala. By genetically labeling the Foxb1 diencephalic lineage, the authors identified labeled cells from the caudal hypothalamus, which migrate into ventral levels of the telencephalon through the continuity between the ventral diencephalon and the telencephalon. This migration occurs after the previously described migratory streams: it starts from E15.5 and is substantial at E18.5 in mouse embryos (Zhao et al., 2008). At birth, abundant Foxb1-lineage cells migrated from the thalamic region into the globus pallidus, the amygdala, and the piriform cortex. These cells express interneuron markers: most of Foxb1-lineage cells in the cortex and in the amygdala express glutamate decarboxylase (Zhao et al., 2008). Some Foxb1-lineage cells in the cortex were Calretinin+, but none of them expressed Calbindin (Zhao et al., 2008). In rodents, as a rule, cortical interneurons are generated in the ganglionic eminences (Marín and Rubenstein, 2003). The migratory stream uncovered by Zhao et al. (2008) would, therefore, allow a specific pool of interneurons to settle the olfactory cortex during late embryonic stages. Whether these interneurons settle a specific layer in the olfactory cortex or whether they are homogeneously distributed remain an open question, as does their specific function in the neuronal network.
Layer formation in the olfactory cortex
Reminiscent of the neocortex layer inside-out development, injections of [3H]thymidine in E14 to E22 timed-pregnant female rats showed significantly more early-born cells settled in piriform cortex deep layer 3 compared with layer 2 (Bayer, 1986; Valverde and Santacana, 1994). The piriform cortex layer 2 is easily delineable with its high density of pyramidal cell bodies. This layer starts to organize from E16 and is well definable by E18 in mouse embryos (Klingler et al., 2015; Fig. 2). BrdU injections at different mouse development stages and analyses of cell identity at postnatal day 7 showed that cell birth date significantly affects not only the laminar position of cells but also their cellular fate (Sarma et al., 2011). In mice, most layer 2 pyramidal cells are born at E12. E14-born cells are less numerous but still differentiate mainly in pyramidal cells. However, E16-born cells display more heterogeneous identities, as follows: 43% differentiate in pyramidal neurons, 24% differentiate in nonpyramidal neurons (interneurons), and 33% differentiate in non-neuronal cells (astrocytes; Sarma et al., 2011). These results show the conservation of a fundamental developmental chronology in both paleocortices and neocortices, with the sequential generation of pyramidal cells, interneurons, and glia. Labeling of cells generated at precise time points during development could be helpful to identify genetic markers of cell populations in the olfactory cortex. So far, nothing is known about the developmental dynamics of layer 3 neurons.
Axons from OB mitral and tufted cells fasciculate to form the LOT and develop collaterals, which invade the ipsilateral olfactory cortex layer 1 from E15.5 to E17.5 in a caudal to rostral sequence (Fig. 3; Haberly, 1983, 2001; Haberly and Feig, 1983; Hirata and Fujisawa, 1999). Layer 1 can be divided into the following two sublayers: superficial layer 1a formed by the terminals of LOT axons, which make synaptic contacts with the apical dendrites of layer 2/3 pyramidal cells; and layer 1b, which is formed by the apical dendrites of layer 2/3 pyramidal cells (Schwob and Price, 1984; Suzuki and Bekkers, 2011; Fig. 3). In the adult mouse brain, layers 1a and 1b are delineable by MAP-2 and Calretinin immunostainings, which label layer 2/3 pyramidal cell dendrites and LOT axons, respectively (Sarma et al., 2011). At birth, layer 1 is not yet subdivided. Layer 1a starts to be distinguishable from postnatal day 7 with visible costaining of MAP-2 and Calretinin (Sarma et al., 2011). These observations suggest that the synaptic contacts between LOT terminals and layer 2/3 pyramidal cell apical dendrites develop during the first postnatal week. During mouse embryonic development, LOT axons develop collaterals, which first invade the CoA at E15.5, then the piriform cortex 1 day later, followed by the more rostrally located regions of the olfactory cortex (Hirata and Fujisawa, 1999). These collaterals allow one LOT axon to make synaptic contacts with layer 2/3 pyramidal cells of different regions of the olfactory cortex (Ojima et al., 1984). Since LOT axons develop after birth, it is rational to assume that the maturation of the circuit is experience dependent, as it has been well described for the primary somatosensory barrel cortex (for review, see Vitali and Jabaudon, 2014). It would be of interest to investigate the consequences of postnatal odorant stimulus depletion in the organization and maturation of olfactory cortex layer 1 connectivity.
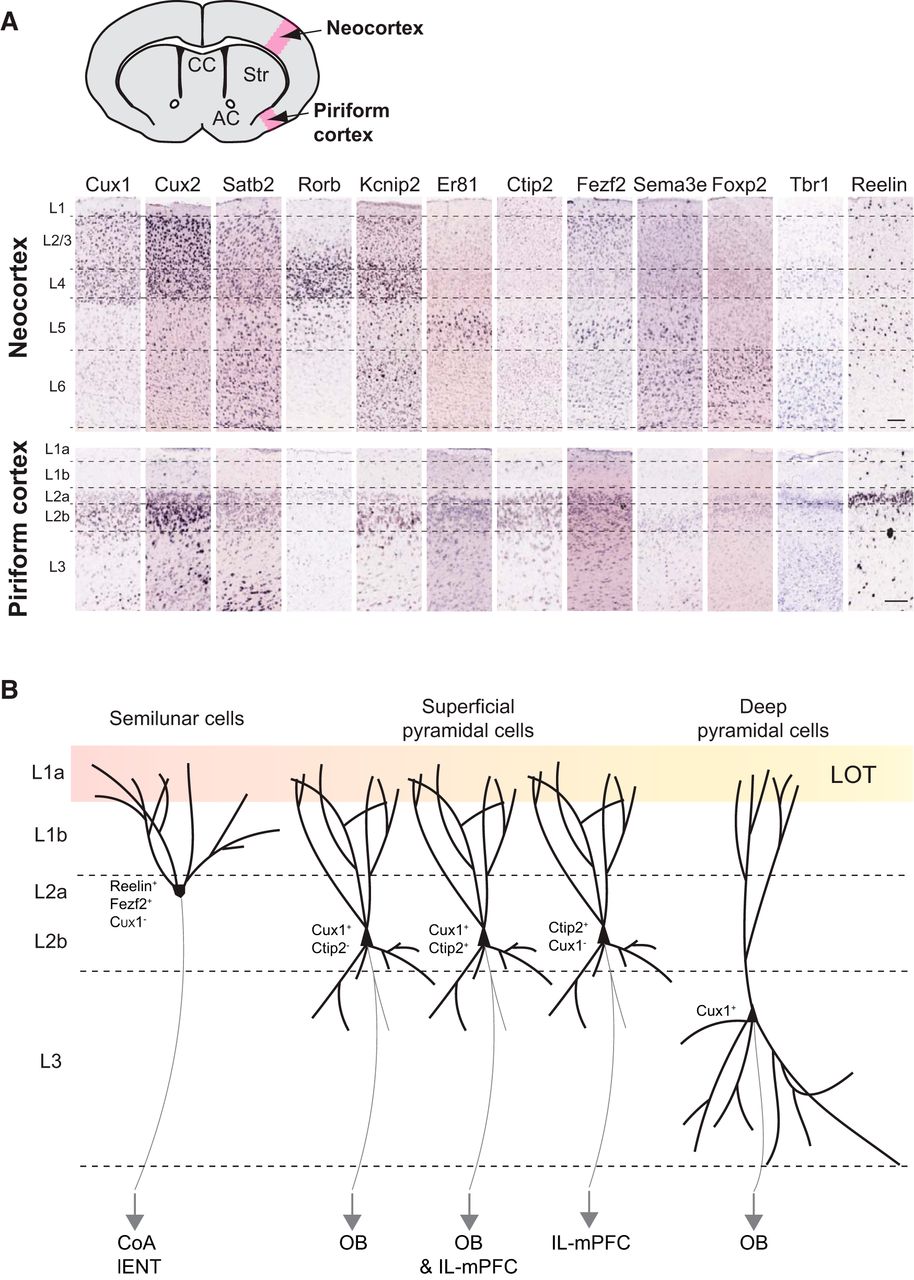
Piriform cortex neuron identities. A, Expression of principal neocortical layer markers in the piriform cortex. In situ hybridizations from Allen Brain Atlas database (postnatal day 56). CC, Corpus callosum; AC, anterior commissure; Str, striatum; L, layer. Scale bar, 200 µm. B, Organization, molecular identities, and known targets of projection neurons in the piriform cortex. LOT, lateral olfactory tract; CoA, cortical amygdala; lENT, lateral entorhinal cortex; OB, olfactory bulb; IL-mPFC, infralimbic medial prefrontal cortex.
Cell types and organization of the circuit implicated in odor processing
Cell types and circuit organization within the piriform cortex
In the piriform cortex, layer 2 pyramidal cells have been initially classified into two populations based on their distinct functions in odor processing: the semilunar and the superficial pyramidal cells (Fig. 3). The semilunar cells are located in the upper part of layer 2 (2a) and display a high spine density in the distal regions of their apical dendrites. These cells receive predominantly afferent excitation from LOT axons, their activity being highly correlated with OB stimulation (Suzuki and Bekkers, 2011). The superficial pyramidal cells are located deeper in layer 2 (2b) and display basal dendrites extending into layer 3 (Fig. 3). They receive weaker afferent inputs from the OB and stronger intracortical excitatory drive through associative fibers, their activity therefore depending on local feedback loops (Haberly and Feig, 1983; Suzuki and Bekkers, 2011; Wiegand et al., 2011; Hagiwara et al., 2012; Fig. 3). Layer 3 is less dense in cell bodies compared with layer 2 and is composed of deep pyramidal cells, which receive minimal afferent inputs from LOT axons, but substantial intracortical excitation (Fig. 3).
To date, molecular identity of the distinct pyramidal neuron within the piriform cortex has been underinvestigated. Most markers defining layers in the neocortex are expressed in the piriform cortex as well (Molyneaux et al., 2007; Fig. 3). Upper layer markers CUX1 and CUX2 (Cubelos et al., 2010) are enriched in piriform cortex layer 2b. Some CUX2+ cells are also present in layer 3. Callosal projection neuron marker SATB2 (Alcamo et al., 2008; Leone et al., 2015) is expressed in piriform cortex layers 2 and 3. It would be of interest to know whether SATB2+ piriform cortex neurons project to the contralateral hemisphere through the anterior commissure and are therefore the counterparts of SATB2+ neocortical callosal projection neurons. Thalamic recipient layer 4 neuron marker RORβ (Jabaudon et al., 2012) shows very weak expression in the piriform cortex. This could be associated with the fact that piriform cortex neurons do not receive sensory input from the thalamus but directly from the OBs. FEZF2 known to be enriched in neocortex layer 5 (Rouaux and Arlotta, 2010; De la Rossa et al., 2013), is enriched in piriform cortex layer 2a (Diodato et al., 2016). Layer 5/6 marker CTIP2 is expressed in piriform cortex layer 2 (Fig. 3). Gene expression comparison between the neocortex and the piriform cortex has been extensively analyzed in the study by Luzzati (2015). He showed that the neocortex layers 2/3 share 42% of enriched genes with the piriform cortex, while deep layers 4 and 6 are less related to the piriform cortex with only 29% of coexpressed genes (Luzzati, 2015). Piriform cortex neurons seem therefore to express common molecular markers enriched in neocortical neurons, but they do not display the same organization within layers. Ramsden et al. (2015) developed a new computational pipeline for high-throughput analysis and comparison of Allen Brain Atlas in situ hybridizations at laminar resolution to study gene expression in the medial entorhinal cortex (MEC). As observed for the piriform cortex, very few genes are uniquely expressed in the medial entorhinal cortex. In addition and contrary to the piriform cortex, deep layers of the medial entorhinal cortex are relatively similar to those of the neocortex, while superficial layers are substantially more divergent at a molecular level (Ramsden et al., 2015). The authors proposed that the medial entorhinal cortex is a type of periarchicortex (paleocortex), a transitional structure between the six-layered neocortex and the three-layered archicortex (Ramsden et al., 2015).
A recent study by Diodato et al. (2016) addressed for the first time the molecular identities of piriform cortex projection neurons using laser capture microdissection of piriform cortex layers and RNA deep sequencing to identify genes differentially expressed within piriform cortex layers in combination with retrograde labeling from piriform cortex targets. With these approaches, they showed that layer 2a semilunar cells project to CoA and lENT and express Reelin and Fezf2 (Fig. 3). The expression of Reelin by excitatory projection neurons in adult brain is specific to the piriform cortex, since in the neocortex, Reelin is mostly expressed by interneurons (Alcántara et al., 1998; Pesold et al., 1998; Ramos-Moreno et al., 2006). These Reelin+ piriform cortex semilunar cells resemble the CR cells from the developing brain (Carceller et al., 2016). Carceller et al. (2016) proposed that the secretion of Reelin by upper piriform cortex layer 2 could participate in the maturation of an immature neuron subpopulation expressing polysialylated-neural cell adhesion molecule and Doublecortin located deeper in layer 2 (Nacher et al., 2001; Luzzati et al., 2009; Klempin et al., 2011). Interestingly, Reelin+ stellate principal cells have been described in the adult MEC layer 2 (Varga et al., 2010). The MEC layer 2 also comprises pyramidal cells, some of which express Doublecortin late in postnatal development (Ray and Brecht, 2016). A role for Reelin in the late maturation of immature neuron subpopulations can therefore also be considered in the case of the MEC.
Besides, piriform cortex pyramidal neurons sending projections back to OB are located in layer 2b and layer 3, display morphology characteristics of superficial and deep pyramidal cells, and express Cux1 (Fig. 3). Interestingly, 3D reconstruction of feedback-projecting neurons after rabies virus injection in the granule cell layer of the main OB showed that these cells are more numerous in the anterior than in the posterior piriform cortex and display a nonrandom organization: piriform cortex neurons next to one another project to similar regions of the OB granule cell layer (Padmanabhan et al., 2016). Neurons projecting to IL subdivision of the medial prefrontal cortex (IL-mPFC) are mainly located in layer 2b and express Ctip2. Moreover, a fraction of layer 2b neurons express both Cux1 and Ctip2 and project to both OB and IL-mPFC (Fig. 3; Diodato et al., 2016). This study points out interesting differences about the molecular organizations of the piriform cortex and the neocortex: while in the neocortex, Cux1 is expressed by superficial layer neurons, and Fezf2 by deep layer 5/6 neurons exclusively; this organization is reversed in the piriform cortex, where Fezf2 is expressed in layer 2a, and Cux1 in layers 2b and 3. Moreover, Cux1 and Ctip2 are expressed in mutually exclusive populations in the neocortex, while a fraction of layer 2b neurons express both markers in the piriform cortex. Interestingly, the authors further showed that in Reelin-deficient mice, where cells of distinct layers are intermingled across the piriform cortex, the molecular identities of neurons projecting to the OB (CUX1+/FEZF2−) compared with neurons projecting to the CoA (CUX1−/FEZF2+) are conserved, despite their position defect, as described in the neocortex (Ogawa et al., 1995; Hevner et al., 2003; Wagener et al., 2010; Boyle et al., 2011; Diodato et al., 2016).
Odor processing in the piriform cortex
In mice, each olfactory sensory neuron in the olfactory epithelium expresses only one olfactory receptor gene, and olfactory sensory neurons expressing a common olfactory receptor send convergent projections to two glomeruli in the main OB (Buck and Axel, 1991; Mombaerts et al., 1996; Malnic et al., 1999, 2010). Despite the precise odotopic organization at these levels of the olfactory circuit, individual odorants evoke a response in sparsely and randomly distributed sets of neurons within the piriform cortex (Rennaker et al., 2007; Poo and Isaacson, 2009; Stettler and Axel, 2009; Ghosh et al., 2011; Miyamichi et al., 2011; Sosulski et al., 2011). Using single-cell recordings from head-restrained awake mice, Zhan and Luo (2010) analyzed odor response profiles of individual neurons in the anterior piriform cortex. Upon odorant presentation, 25% of projection neurons were broadly excited and rarely inhibited, 25% showed no excitation and a clear inhibition, and 50% displayed very selective responses in terms of excitation and inhibition (Zhan and Luo, 2010). The apparent random connections from the OB mitral and tufted cell axons to the piriform cortex suggest that the representations of odors in the olfactory cortex are learned by experience. The piriform cortex would detect odors by comparing them with a previously acquired library of odors (Sullivan and Wilson, 2003) and build odor units from the chemicals identified upstream in the olfactory circuit (Johnson et al., 2000). The piriform cortex can therefore be seen as a memory tool optimized in the storage of odor synaptic representations (Barkai et al., 1994).
Olfactory representations within the piriform cortex are strongly shaped by recurrent excitatory and inhibitory intracortical connections (Franks et al., 2011; Poo and Isaacson, 2011). While each piriform cortex pyramidal cell receives only 200 inputs from mitral/tufted OB cells, it receives at least 2000 recurrent excitatory inputs (Davison and Ehlers, 2011; Franks et al., 2011). Contrary to the primary visual and somatosensory cortices, recurrent piriform cortex axons form synapses with the same probability, whatever the distance from the soma (Franks et al., 2011). Local cortical application of baclofen, a GABAB antagonist, abolishes intracortical associational transmission from excitatory neurons without affecting LOT-evoked excitatory responses (Tang and Hasselmo, 1994; Franks and Isaacson, 2005). Using this pharmacologic approach, Poo and Isaacson (2011) showed that the recruitment of intracortical input, rather than OB input, largely determines the strength of odor-evoked excitatory synaptic transmission in the piriform cortex. Using a novel transgenic mouse model allowing the expression of channel rhodopsin (ChR2) in a subset of semilunar cells, Choy et al. (2015) showed that ChR2-expressing semilunar cells avoid targeting neighboring semilunar cells but provide strong monosynaptic associational excitation of superficial and deep pyramidal cells. Piriform cortex interneurons also play an important role in odor processing. They allow increasing discrimination of an odor and prevent nonspecific excitations. Only 10% of neurons will be activated by a given odor (Rennaker et al., 2007; Poo and Isaacson, 2009; Zhan and Luo, 2010). This signal transformation may allow the piriform cortex to perceive a complex mix of odors as an odorant object separated from its pure components. Feedforward inhibition is mediated by layer 1 horizontal and neurogliaform interneurons, which receive direct inputs from the LOT and synapse onto apical dendrites of pyramidal cells, and are thought to play a prominent role in dendritic integration of afferent input from LOT axons in all piriform cortex excitatory cells. Feedback inhibition is mediated by layer 2/3 bitufted regular spiking interneurons, which receive little direct LOT input and synapse onto pyramidal cell bodies and basal dendrites, with deep pyramidal cells receiving the strongest recurrent inhibition (Stokes and Isaacson, 2010; Suzuki and Bekkers, 2010a,b; Taniguchi, 2014; Large et al., 2016). Semilunar cells have been shown to directly activate layer 3 GABAergic interneurons (fast spiking, regular spiking, and neurogliaform), but neither neurogliaform nor horizontal interneurons of layer 1a (Choy et al., 2015). The feedback inhibition would dominate the feedforward inhibition in controlling the activation of piriform pyramidal cells (Franks et al., 2011).
Synaptic organization of the cortex: from 3 to 6 layers
The cortical type microcircuit consists of a superficial plexiform layer 1, where extrinsic and intrinsic projections meet the apical dendrites of pyramidal neurons located in deeper layers, and is responsible for the generation of recurrent excitation and inhibition (Haberly, 1990). The neocortex shares the basic microcircuits with the three-layered allocortex, but displays a higher number of neurons and layers (Shepherd, 2011; Fournier et al., 2015). The neocortex could then be described as a double allocortex: two couples of pyramidal cell populations, formed by upper layers 2/3 and deeper layers 5/6, are each located below plexiform layers 1 and 4, respectively, carrying extrinsic inputs (Shepherd, 2011; Luzzati, 2015). In contrast to the dorsal cortex of reptiles or to the piriform cortex of mammals, where sensory afferents come from the top, in the neocortex the thalamic afferents ascend from the depth and efferents descend through the same layers. Both inputs and outputs have therefore potential access to all cells in every layer (Shepherd, 2011). Moreover, the generation of superficial and deep layers in the neocortex allows expansion of the combinatorial possibilities for intracortical and subcortical processing (Shepherd, 2011). Neocortex upper layers are evolutionarily the most recent. Interestingly and counterintuitively, piriform cortex neurons express most molecular markers found in these neocortex upper layers. These similarities lead to the hypothesis of an olfactory cortex-based developmental program for the evolution of neocortical layers 2/3 (Luzzati, 2015). The early neocortical column would therefore result of the superposition of the lateral cortex (olfactory cortex) and the dorsal cortex (Luzzati, 2015). Careful studies of piriform cortex cell lineages and neuron molecular identities are needed to understand what specific cell types are found in evolutionary “old” olfactory cortex and neocortex layers.
Concluding remarks
The olfactory cortex, so-called “simple” cortex in the literature when compared with the six-layered neocortex, is often presented as a general model for cortical sensory processing. However, the molecular identities and the specific functions of cells composing each olfactory cortex layer remain poorly described. The sequential generation of pyramidal cells, interneurons, and glia, as well as the inside-out migration of pyramidal cells seem to be conserved processes in both olfactory cortex and neocortex during development. In the neocortex, excitatory neurons derive from pallial VZ and migrate radially, while inhibitory neurons derive from subpallial VZ and migrate tangentially. This seems not as obvious in the olfactory cortex, where some neurons from the pallium differentiate in inhibitory neurons and some neurons from the subpallium in excitatory neurons (Garcia-Moreno et al., 2008; Fig. 1). The routes of migration toward the olfactory cortex are particularly complicated since olfactory cortex neurons have multiple origins in the VZ and some of them migrate over a long distance to reach their final location. These routes implicate tangential as well as radial migrations. The most studied migratory stream to the olfactory cortex is the lateral cortical stream, which is thought to be the main stream for olfactory cortex excitatory neurons. Some radial glial processes (radial glial palisade) have been identified along this stream and possibly allow the tangential-like migration of excitatory neurons along radial glia (Carney et al., 2006; Fig. 2).
The diversity of their VZ origins implies that the cells composing the olfactory cortex come from different cell lineages and suggests that different cell populations should be definable among olfactory cortex layers. So far, two pyramidal cell populations have mainly been described in the piriform cortex layer 2 based on their morphological and electrophysiological properties (Suzuki and Bekkers, 2011; Fig. 3). However, recent studies started decoding the molecular signatures of projection neurons in respect to their connectivity properties (Diodato et al., 2016; Padmanabhan et al., 2016; Fig. 3). Further investigations will allow understanding whether the different VZ origins and migratory properties of olfactory cortex neurons during development are correlated with their heterogeneous identities and functions in odor processing. Addressing olfactory cortex neuron transcriptional identities throughout development will help to molecularly characterize cell populations with specific properties in the olfactory cortex, in order to compare them with populations identified in the neocortex, in terms of layer localization, connectivity (input and output), and function. The piriform cortex and the dorsal cortex of reptiles (corresponding to visual cortex) both seem to process sensory inputs as high-order cortical areas rather than primary sensory neocortex. Computations performed by high-order cortical areas seem therefore to be ancestral, while computations performed at initial stages of neocortical processing appeared later in evolution, possibly linked to the additions of new layers (Fournier et al., 2015). Unraveling precise neuronal origins and identities in both the neocortex and the olfactory cortex will further elucidate the evolutionarily conserved properties of sensory cortices.
Acknowledgments
Acknowledgments: I thank A. Hay, G. Limoni, and U. Tomasello for constructive comments and corrections on the manuscript. I also thank L. Goutebroze and D. Jabaudon for their support.
Footnotes
The author declares no competing financial interests.
This work was supported by University of Geneva (Geneva, Switzerland).
This is an open-access article distributed under the terms of the Creative Commons Attribution 4.0 International, which permits unrestricted use, distribution and reproduction in any medium provided that the original work is properly attributed.
References
In this issue

{kind=link}
{kind=link}
{kind=link}



