

Article Figures & Data
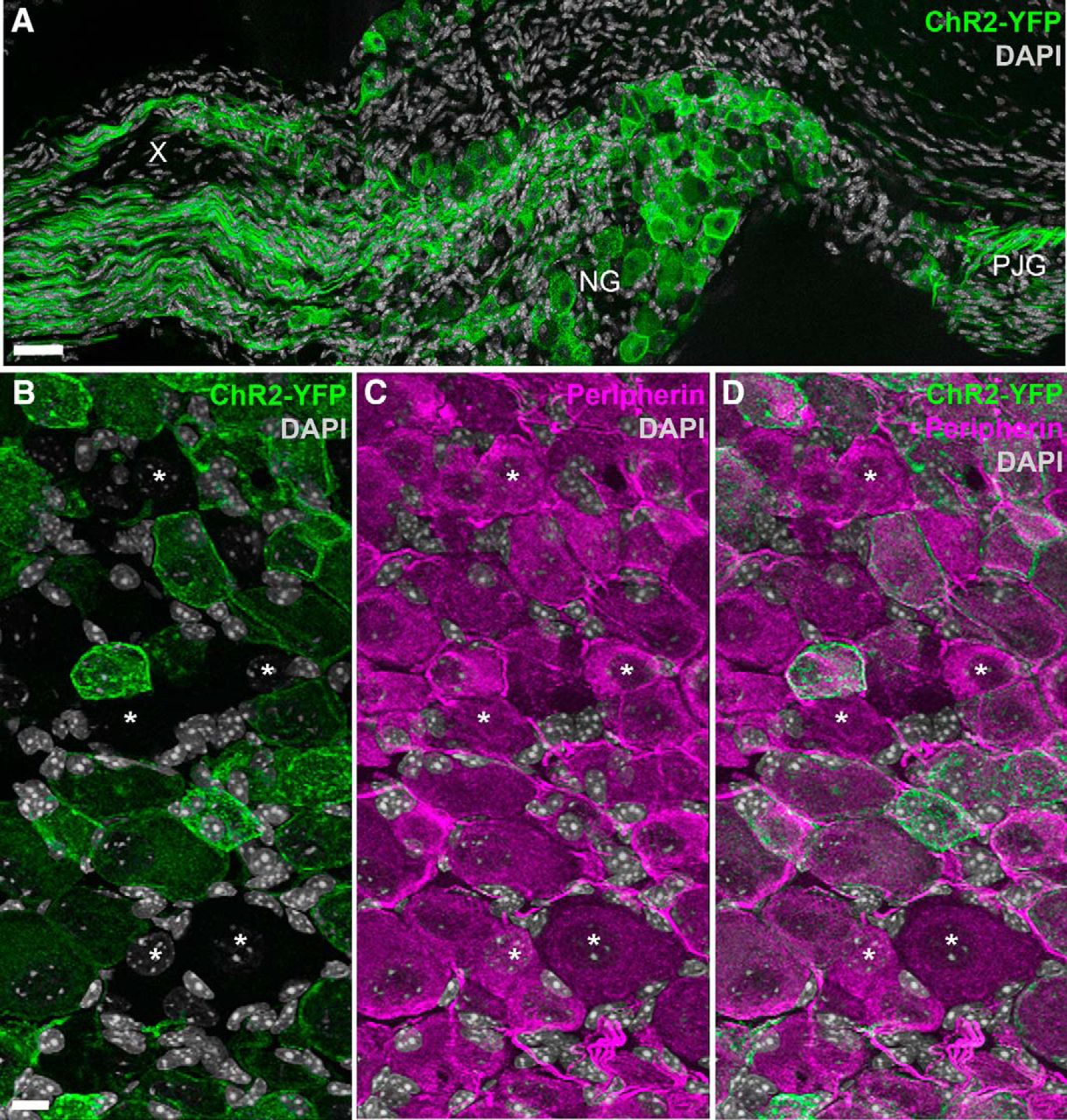
- Figure 1.
Distribution of ChR2-YFP fluorescence in the nodose ganglion of Nav1.8-Cre-ChR2-YFP mice. A, Many neuronal cell bodies and axons were brightly fluorescent in the nodose ganglion (NG) and petrosal-jugular ganglion (PJG) (three images were horizontally stitched together). B–D, Details of NG peripherin-labeled neurons (Alexa Fluor 594). ChR2-YFP is apparent in the membrane and cytoplasm of many NG neurons (but not all) (B). Asterisks are positioned over representative peripherin-positive neurons that were not labeled with ChR2-YFP (B, C). X, cervical vagus nerve. Scale bars, 40 μm in A; 20 μm in B (also applies to C, D).
- Figure 2.
Distribution of ChR2-YFP fluorescence in the dorsovagal complex of Nav1.8-Cre-ChR2-YFP mice. A, Vagal afferents terminating in the AP and NTS were labeled with ChR2-YFP (three horizontally stitched images). B, Highly varicose ChR2-YFP–labeled axons circling the cell body of one TH-positive neuron (Alexa Fluor 594) located in the NTS. C, Single optical section of ChR2-YFP–labeled axons in the AP revealing thin varicose axons of varying sizes. Asterisks are positioned over the presumptive locations of the postsynaptic cell bodies, whereas arrowheads indicate representative vagal varicosities. DMV, dorsal nucleus of the solitary tract; sol, solitary tract; commissural part of the nucleus of the solitary tract; SolDM, dorsomedial part of the nucleus of the solitary tract; SolM, intermediate part of the nucleus of the solitary tract; SubP, subpostrema area. Scale bars, 60 μm in A; 5 μm in B and C.
- Figure 3.
Distribution of ChR2-YFP fluorescence throughout the rostrocaudal extension of the dorsovagal complex of one representative Nav1.8-ChR2-YFP mouse. Several digital images were stitched together (epifluorescence with Apotome filter). The number in the bottom right corner indicates the approximate distance from Bregma according to the Franklin and Paxinos Mouse Brain Atlas (Third Edition). 4v, fourth ventricle; cc, central canal.
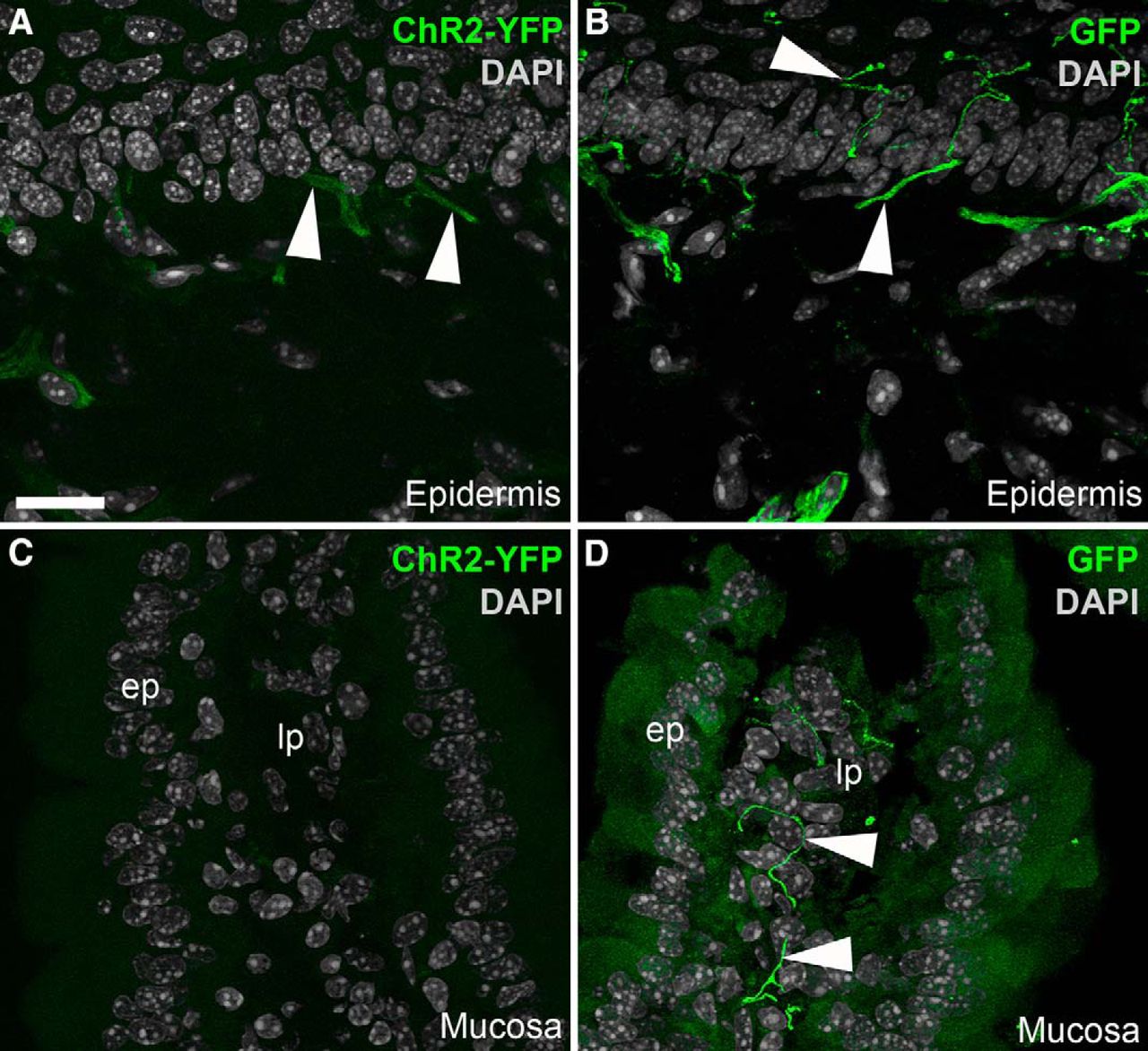
- Figure 4.
Identification of ChR2-YFP–positive fibers in peripheral tissues from Nav1.8-Cre-ChR2-YFP mice. A, Weakly fluorescent structures resembling nerve endings (arrowheads) were observed in the epidermis of the forepaw. B, GFP immunolabeling greatly enhanced the labeling of individual axons ramifying into the epidermis. C, We could not detect endogenous YFP fluorescence in the duodenal mucosa before immunolabeling. D, YFP-labeled axons were evident in the lamina propria (lp) when the tissue was stained for GFP. The antibody produced a small amount of nonspecific background staining in the epithelium (ep). Arrowheads indicate representative YFP-positive fibers. Scale bars, 20 μm in A (also applies to B–D).
- Figure 5.
CART(55–102) detection in the nodose ganglion (NG) of Nav1.8-Cre-ChR2-YFP mice. A, CART-positive cells (Alexa Fluor 594) resembling neurons were observed throughout the NG. The two cells located in the inset are represented at high magnification in C. B, Omission of the primary antibody eliminated CART immunoreactivity. Only a few debris and background could be observed in the NG. C, Two adjacent ChR2-YFP-, CART-positive neurons are shown. CART is detected in vesicle-like structures in the cytoplasm and proximal axon. The larger neuron contained more immunoreactivity than the smaller neuron. Two other YFP-labeled neurons did not contain CART. Scale bars, 50 μm in A and B; 5 μm in C.
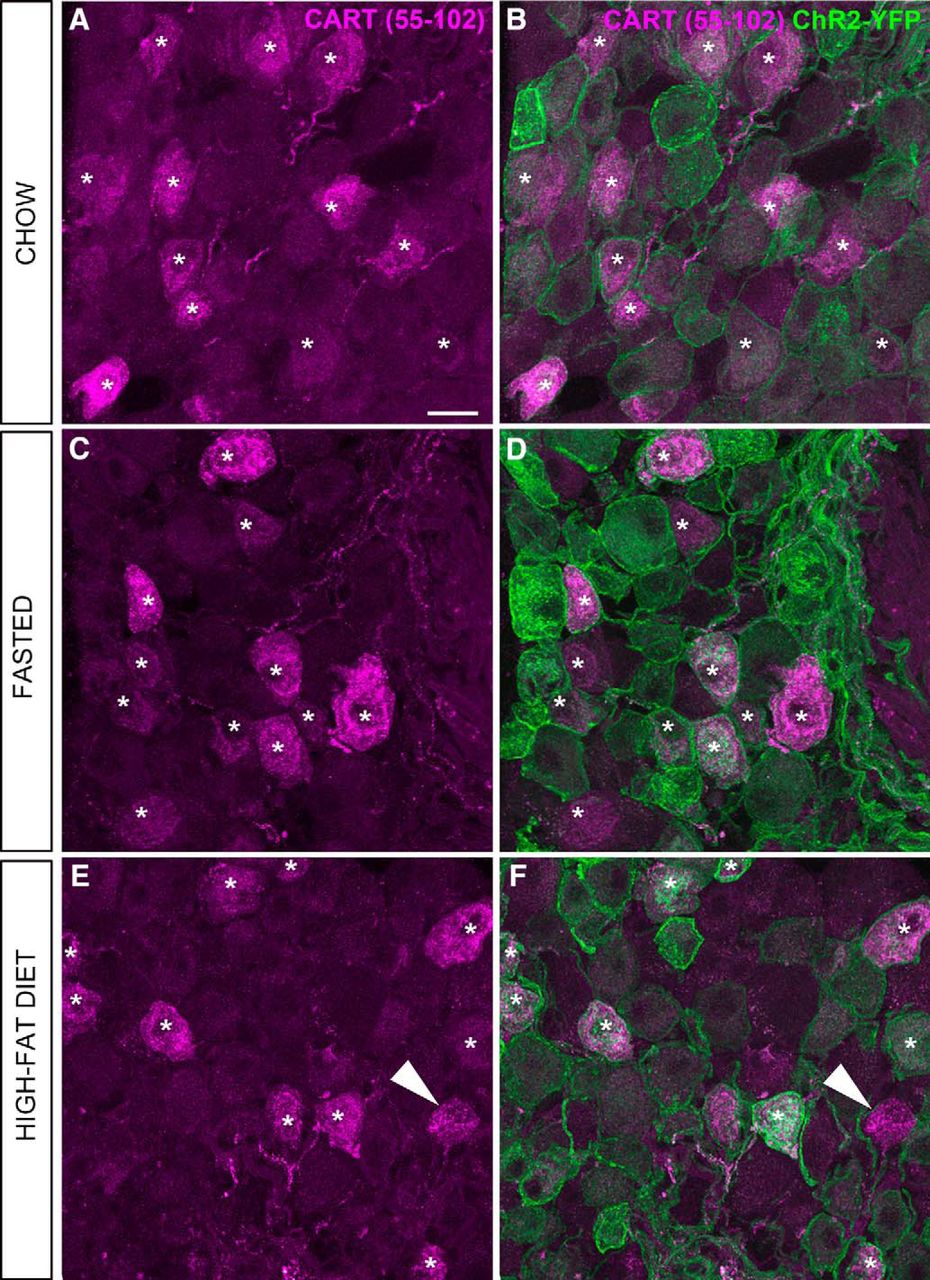
- Figure 6.
CART(55–102) immunolabeling of the nodose ganglion (NG) of metabolically challenged Nav1.8-Cre-ChR2-YFP mice. A, C, E, CART-positive perikarya (Alexa Fluor 594) across the feeding groups. B, D, F, CART staining and endogenous ChR2-YFP fluorescence delineate the outline of the vagal afferents. Asterisks are positioned over representative CART-positive cells. Note that CART immunoreactivity is almost always contained within the cell membrane of the YFP-labeled cells, with a few rare exceptions (arrowhead). By visual inspection, the CART distribution pattern and intensity appeared comparable across the feeding groups. Scale bars, 20 μm in A; applies to all images.
- Figure 7.
Estimates of CART-positive neurons and afferents across the feeding groups. A, The percentage of YFP- and CART-positive neurons in the nodose ganglion (NG) of Nav1.8-Cre-ChR2-YFP mice was not influenced by feeding status (n = 7–8 per group). B, The percentage of CART-positive varicosities in the AP was identical in Nav1.8-Cre-ChR2-YFP mice under different feeding conditions. Data are expressed as the average of the mean percentage ± maximum values (n = 7–8 per group). Each circle represents one value from one mouse. C, Frequency of CART immunoreactivity profiles with varying intensity across feeding groups (n = 7 per group). On the left, frequency scattergraph of distributions of individual CART-positive cell profiles with median and interquartile range. Each gray dot is one cell profile. The total number of profiles examined in each group is annotated. On the right, the same data were represented as stacked bars after categorizations of immunoreactivity level. Data are provided as mean percentage ± SEM. D, Frequency of cells expressing pCART mRNA in the nodose ganglia of fed (n = 4) and fasted (n = 3) wild-type mice. Cells were scaled based on the intensity of hybridization signals. The total number of profiles examined in each group is annotated. Stacked bars provide mean percentage ± SEM.
- Figure 8.
A, B, CART(55–102)-positive neurons in the nodose ganglia of lean Zucker rats. Several digital images were stitched together (epifluorescence with Apotome filter). Many CART-positive neurons (Alexa Fluor 594) were observed in the nodose ganglion of fed and fasted rats. White arrows indicate examples of CART neurons. Scale bar, 100 μm in A; applies to B.
- Figure 9.
CART(55–102) immunolabeling of the dorsovagal complex of metabolically challenged Nav1.8-Cre-ChR2-YFP mice. A, B, C, CART-positive (Alexa Fluor 594) perikarya and fibers of various origins were observed throughout the NTS and AP. We noticed a trend toward increased immunoreactivity in the fasted groups. However, our estimates (see Fig. 6) indicate that the levels of CART in vagal afferents remained unchanged across feeding conditions. D, E, F, CART immunoreactivity combined with endogenous ChR2-YFP fluorescence. G, CART was very abundant in the AP. In particular, many YFP-labeled varicosities of vagal origin were enriched for CART. However, we also observed CART immunoreactivity that was not contained in ChR2-YFP–labeled fibers. H, High magnification of the distribution of CART immunoreactivity in the vagal fibers of the AP in a single optical plan. Of note, CART frequently labeled the cytoplasm contained within the YFP-labeled varicosities, which are indicated by arrowheads. Scale bars, 60 μm in A, applies to B–F; 15 μm in G; 5 μm in H.
- Figure 10.
Detection of prepro-CART (pCART) and prepro-MCH (pMCH) mRNAs using chromogenic ISH. A, pCART hybridization signals (brown DAB; bright-field optics) were strong in select hypothalamic nuclei. B, Throughout the nodose ganglion of fed mice, pCART signals of varying intensity were observed in many cell profiles. C, The nodose ganglion of fasted mice also contained pCART hybridization signals. D, Details of the hybridization signal in the nodose ganglion of one fasted mouse. Please note representative cell profiles without signal (*), or with low (l), medium (m), and high (h) signals. E, Hybridization signals for pMCH were very strong in neurons of the lateral hypothalamus. F, G, In contrast to the hypothalamus, pMCH signals were not observed in the nodose ganglia of fed and fasted mice. H, Details of the nodose ganglion of one fasted mouse showing several neuronal profiles completely devoid of signals (*). Tissue was counterstained with hematoxylin. 3V, third ventricle; Arc, arcuate nucleus; DMH, dorsomedial hypothalamus; LHA, lateral hypothalamus; NG, nodose ganglion; VMH, ventromedial hypothalamus; ZI, zona incerta. Scale bars, 500 μm in A and E; 50 μm in B, C, F, and G; 20 μm in D and H.
- Figure 11.
Absence of MCH staining in the vagal afferents of metabolically challenged Nav1.8-Cre-ChR2-YFP mice. A, An antiserum against prepro-MCH (pMCH) labeled perikarya (Alexa Fluor 594) in the lateral hypothalamus (LHA). B, PMCH was undetectable in the nodose ganglion (NG) of fasted Nav1.8-Cre-ChR2-YFP mice. C, D, An antiserum against the MCH peptide labeled very few axons (arrowhead) in the medial NTS (SolM). MCH immunoreactivity was never observed in the YFP-labeled fibers of vagal origin. Arrowheads point to one MCH-positive axon. sol, solitary tract. Scale bars, 40 μm in A and B; 10 μm in C.
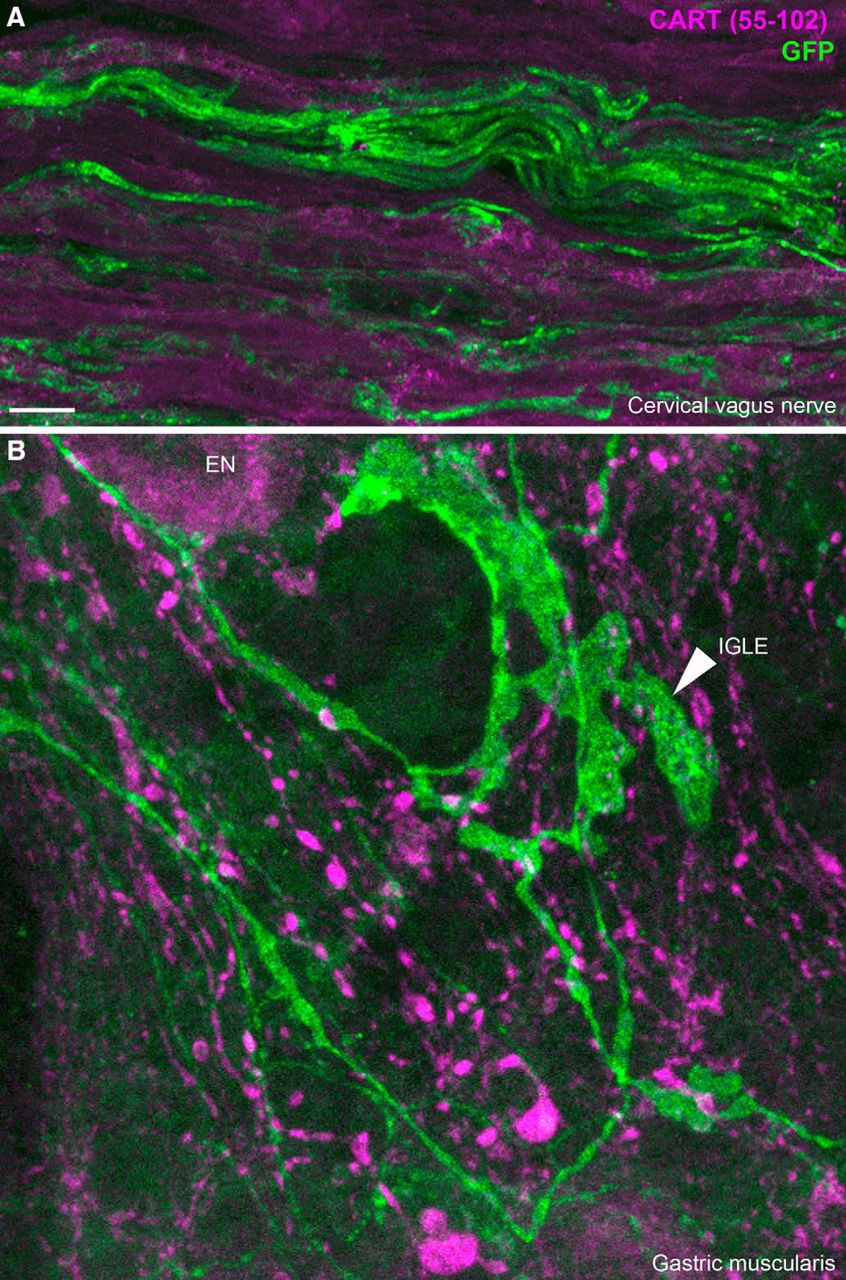
- Figure 12.
CART(55–102) immunolabeling of peripheral ChR2-YFP–labeled fibers. A, ChR2-YFP fibers contained within the cervical vagus nerve. CART was not detected in the vagus nerve. We obtained similar results in the cervical and subdiaphragmatic nerves on both sides. B, In whole mounts of the gastric muscularis, numerous GFP-positive axons and specialized endings can be observed. The leafy structure may correspond to an intraganglionic laminar ending (arrowhead) of vagal origin. CART immunoreactivity was abundant in large varicose fibers and, to a lesser extent, the cell body of enteric neurons (EN). Nonetheless, CART immunoreactivity was not found in GFP-stained fibers. IGLE, intraganglionic laminar ending; EN, enteric neuron. Scale bar, 10 μm in A and B.












Antibody Manufacturer Catalog no. Lot no. Host Working dilution Immunogen Primary antibody Peripherin EMD Millipore AB1530 2446692 Rabbit 1:500 trp-E-peripherin fusion protein containing all but the four N-terminal amino acids of rat peripherin CART(55–102) Phoenix Pharmaceuticals H-003-62 01251-10; 01251-6 Rabbit 1:800–1:1000 Ile-Pro-Ile-Tyr-Glu-Lys-Lys-Tyr -Gly-Gln-Val-Pro-Met-Cys-Asp-Ala-Gly-Glu-Gln-Cys-Ala-Val-Arg-Lys-Gly-Ala-Arg-Ile-Gly-Lys-Leu-Cys-Asp-Cys-Pro-Arg-Gly-Thr-Ser-Cys-Asn-Ser-Phe-Leu-Leu-Lys-Cys-Leu [disulfide bonds between Cys1-Cys3,Cys2-Cys5,Cys4-Cys6] CART 1–39 Phoenix Pharmaceuticals H-003-63 01102 Rabbit 1:800–1:1000 pGlu-Glu-Asp-Ala-Glu-Leu-Gln-Pro-Arg-Ala-Leu-Asp-Ile-Tyr-Ser-Ala-Val-Asp-Asp-Ala-Ser-His-Glu-Lys-Glu-Leu-Pro-Arg-Arg-Gln-Leu-Arg-Ala-Pro-Gly-Ala-Val-Leu-Gln MCH Phoenix Pharmaceuticals H-070-47 01629-3 Rabbit 1:1000 Asp-Phe-Asp-Met-Leu-Arg-Cys-Met-Leu-Gly-Arg-Val-Tyr-Arg-Pro-Cys-Trp- Gln-Val TH Abcam ab101853 GR120879-17 Goat 1:1000 Synthetic peptide corresponding to human TH(30–100) (N terminal) ProMCH Santa Cruz Biotechnology sc-14509 B2415 Goat 1:100–1:1000 20-amino-acid peptide near the C-terminus of pro-MCH precursor of human origin GFP Aves Laboratory GFP-10120 GFP697986 Chicken 1:1000 GFP emulsified in Freund’s adjuvant Secondary antibody Anti-rabbit Alexa Fluor 594 Life Technologies A21207 1256153 Donkey Anti-goat Alexa Fluor 594 Life Technologies A11058 1608643 Donkey Anti-chicken Alexa Fluor 488 Life Technologies A11039 1356650 Goat Gene Accession no. Target region Catalog no., channel Chromogenic label CARTPT NM_013732.7 11–860 432001-c1 DAB, brown PMCH NM_029971.2 4–652 478721-c1 DAB, brown
In this issue

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}



