

Article Figures & Data
Figures
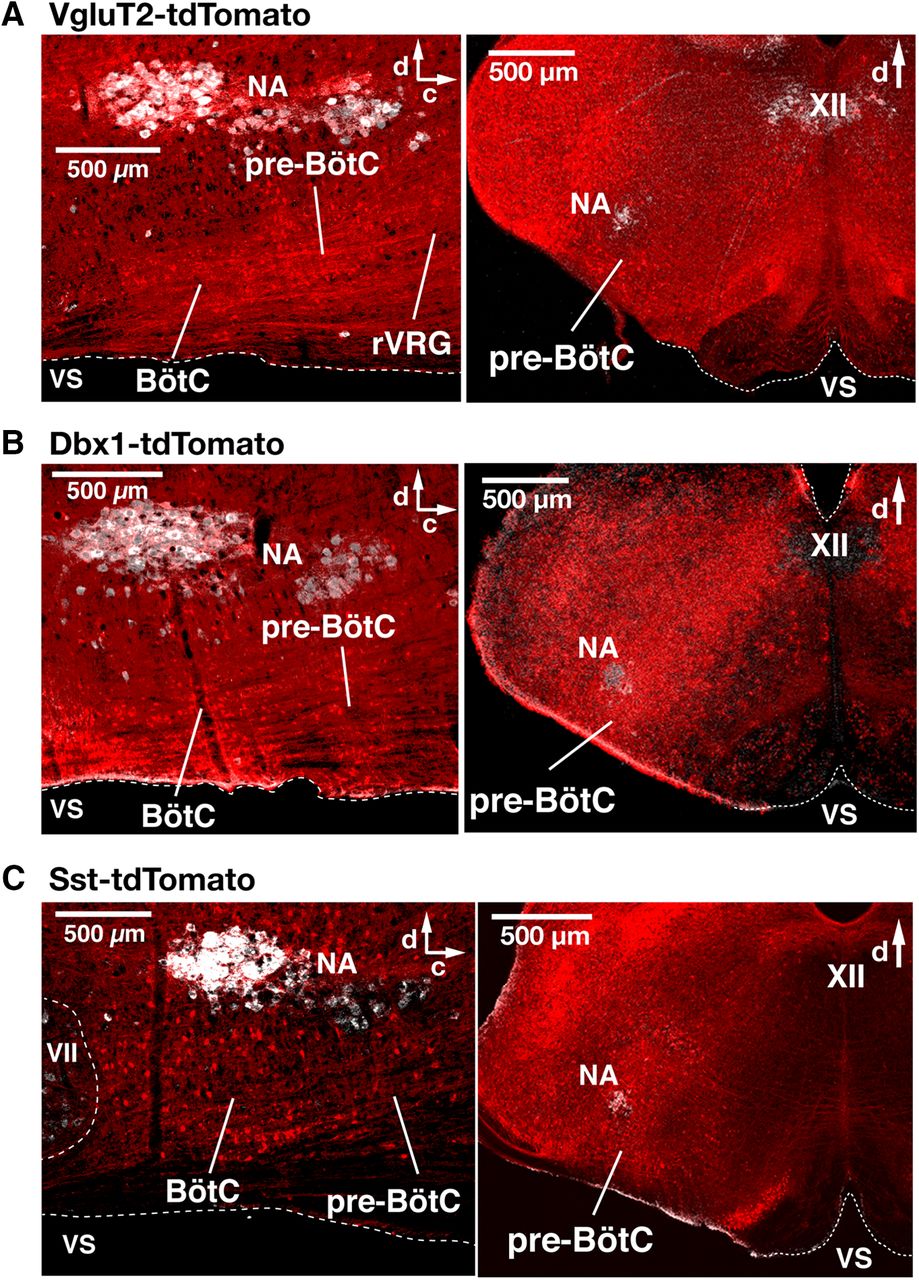
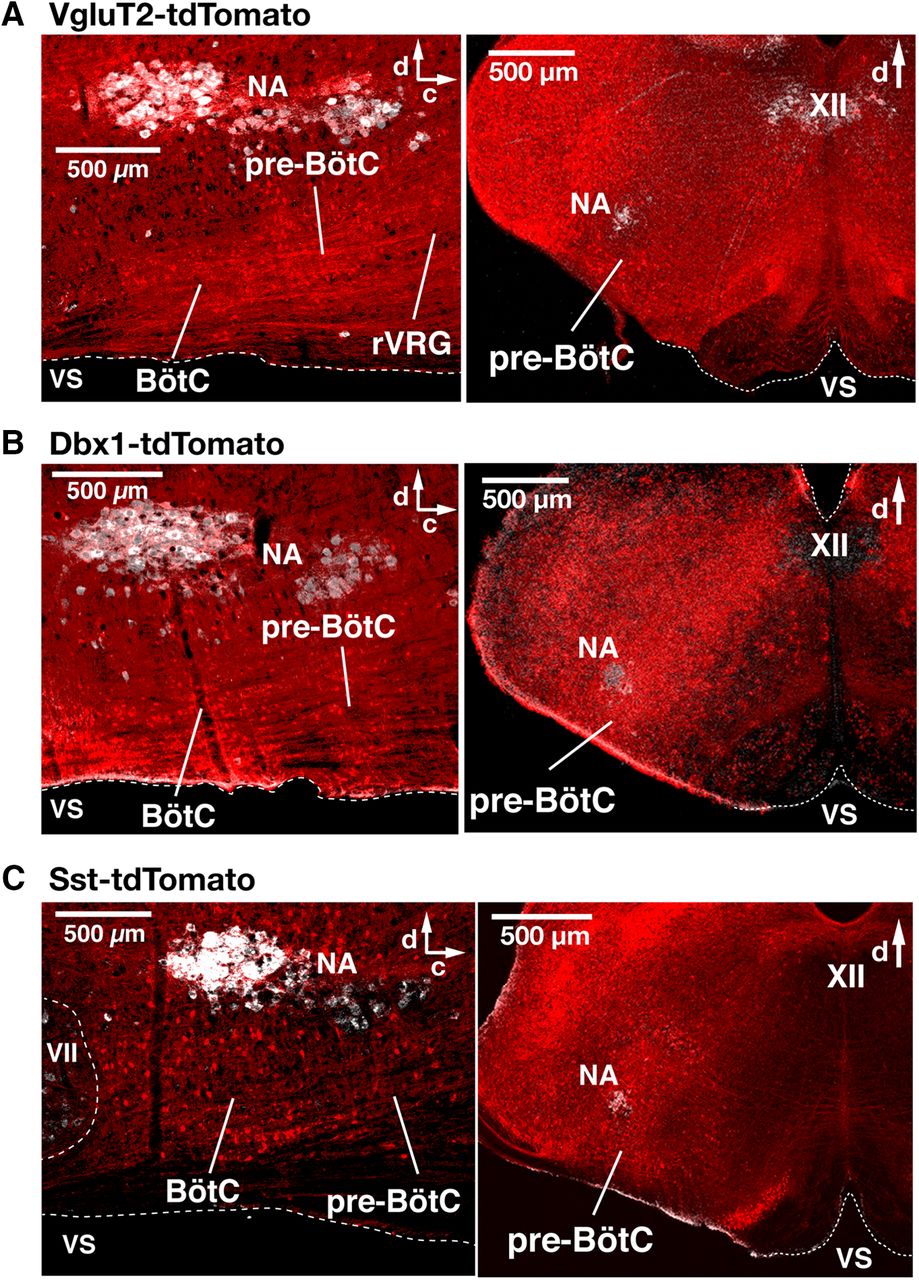
- Figure 1.
Spatial distribution of Cre-dependent tdTomato-labeled neurons in the medullary reticular formation in VgluT2-tdTomato, Dbx1-tdTomato, and Sst-tdTomato double-Tg mouse strains. A–C, Confocal fluorescence microscopy images of parasagittal sections at the level of nucleus ambiguus (NA) and coronal sections at the level of the pre-BötC from adult VgluT2-tdTomato (A), Dbx1-tdTomato (B), and Sst-tdTomato (C) mice, representing an overview of distributions of Cre-dependent tdTomato-labeled neurons (red). Motoneurons of NA and the XII immunostained with ChAT antibody (white) provide reference landmarks for levels of the medulla represented in the sections. The parasagittal sections illustrate the extensive distribution of tdTomato-labeled neurons throughout the ventral medullary reticular formation, including within the pre-BötC, the BötC, and the rVRG. Coronal sections illustrate that tdTomato-labeled neurons are distributed within the pre-BötC, and adjacent regions of the reticular formation (RF), including the intermediate RF dorsomedial to NA, where hypoglossal premotor neurons are located. VS, Ventral surface; d, dorsal; c, caudal.
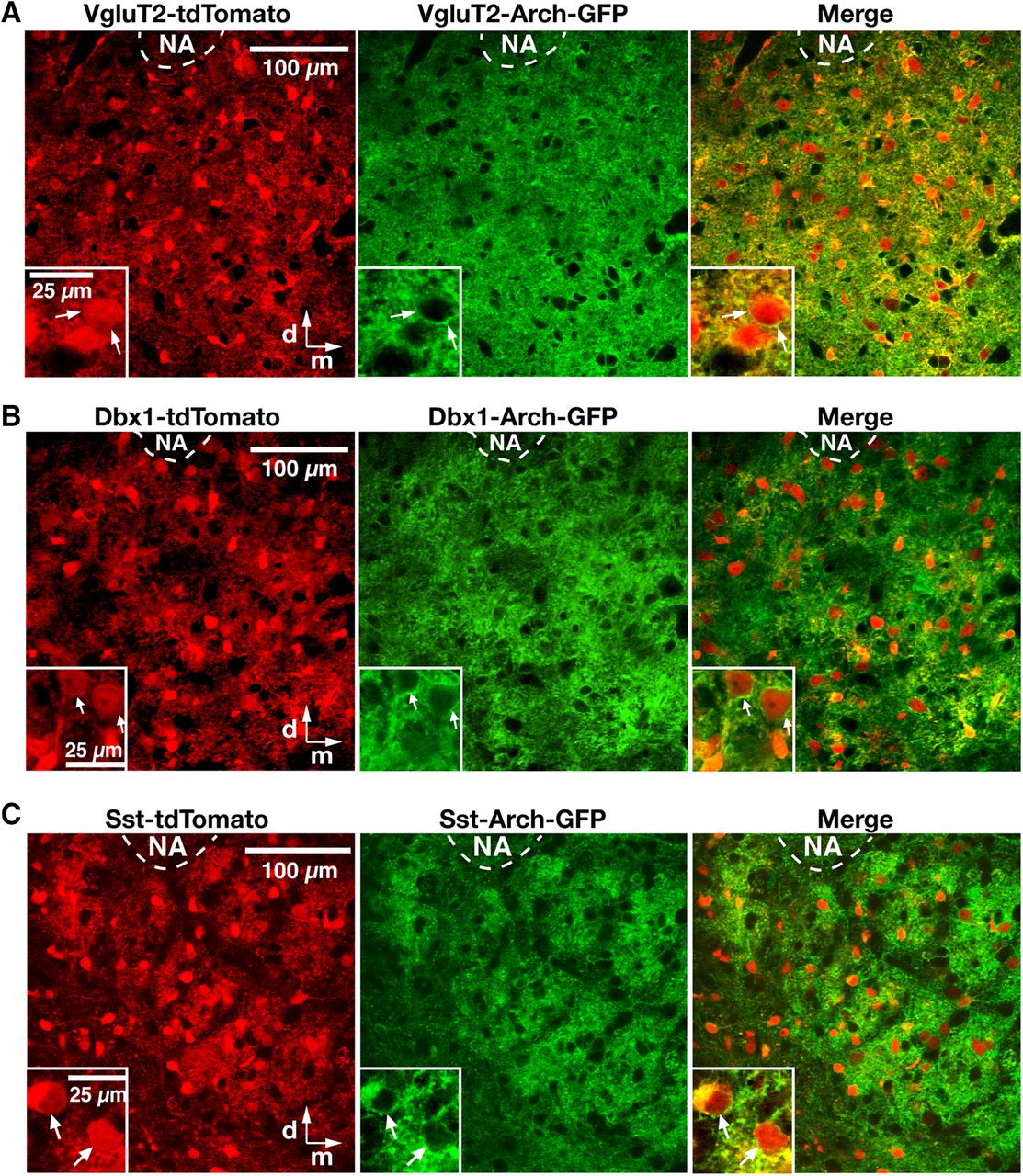
- Figure 2.
Cre-dependent tdTomato reporter neuronal labeling and Arch-GFP expression in the pre-BötC region in triple-transgenic mice. A–C, Two-photon LSM single optical plane “live” images of the pre-BötC region ventral to nucleus ambiguus (NA) on one side of the medulla in an in vitro neonatal medullary slice from the VgluT2-tdTomato-Arch-GFP (A), Dbx1-tdTomato-Arch-GFP (B), and Sst-tdTomato-Arch-GFP (C) triple-Tg mouse lines, presenting an overview of distributions of Cre-conditional tdTomato reporter protein labeling (red) and Arch-GFP fusion protein expression (green) of neurons throughout the pre-BötC region. Insets, Higher-magnification images illustrating neurons with Arch-GFP expression in somal membranes, as confirmed in the merged image for tdTomato-labeled VgluT2-expressing, Dbx1-derived, and Sst-expressing pre-BötC neurons. All images have the same dorsomedial anatomical orientation. d, Dorsal; m, medial.
- Figure 3.
Validation of VgluT2-Cre line and glutamatergic phenotype of Dbx1-derived and Sst-expressing pre-BötC neurons. A, Confocal fluorescence microscopy images of a subregion within the pre-BötC in a fixed coronal section from VgluT2-tdTomato double-Tg strains, showing Cre-dependent tdTomato labeling (red) and immunolabeling with glutaminase antibody (green) in pre-BötC neurons. Merged image illustrates that essentially all of the Cre-mediated tdTomato-labeled VgluT2 neurons coexpress glutaminase, which is consistent with a glutamatergic phenotype of these neurons. B, C, Confocal fluorescence microscopy images of a pre-BötC subregion in a fixed coronal section from Dbx1-tdTomato (B) and Sst-tdTomato (C) double-Tg strains, showing neuronal tdTomato labeling (red) and immunolabeling with glutaminase antibody (green). Merged image illustrates tdTomato-labeled Dbx1-derived (B) and Sst-expressing (C) pre-BötC neurons that coexpress glutaminase (arrows). Examples of glutaminase antibody-labeled neurons without somal tdTomato expression are also indicated (arrowheads). All images have the same dorsomedial anatomical orientation. d, Dorsal; m, medial.
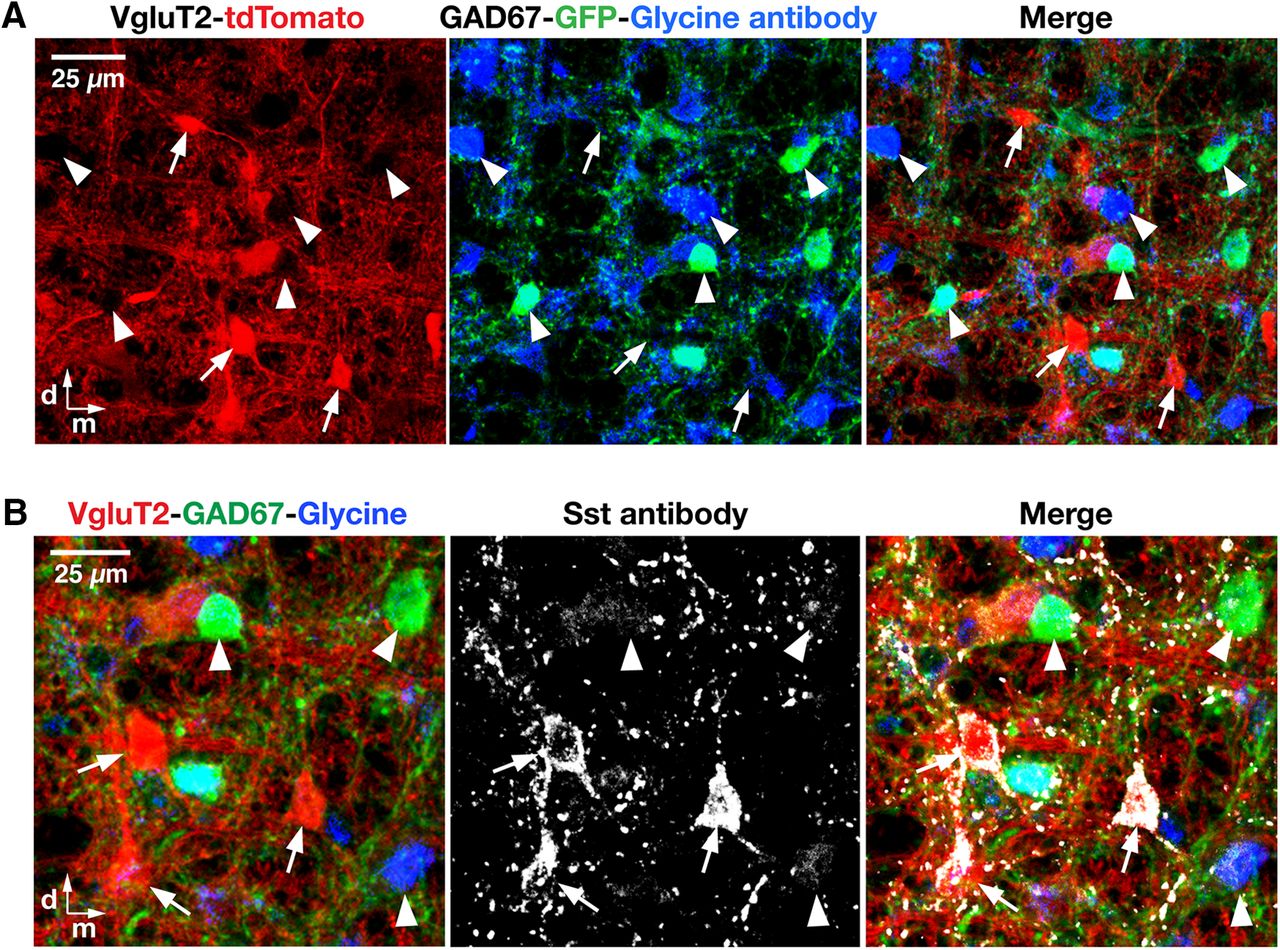
- Figure 4.
Glutamatergic phenotype of VgluT2- and Sst-expressing pre-BötC neurons in the VgluT2-tdTomato-GAD67-GFP triple-transgenic mouse line. A, Confocal images of a subregion within the pre-BötC in a fixed coronal section from the VgluT2-tdTomato-GAD67-GFP triple-Tg strain, showing Cre-dependent VgluT2-tdTomato neuronal labeling (red), GAD67-knock-in GFP neuronal labeling (green), and immunolabeling with glycine antibody (blue) in the pre-BötC neurons. Merged image confirms no overlap of tdTomato neuronal somal labeling (examples indicated by arrows) with GAD67-GFP or glycine antibody expression (arrowheads), except for labeling of terminals in some cases, also verifying that all the Cre-driven tdTomato-labeled VgluT2 neurons are glutamatergic neurons. B, Confocal images of a pre-BötC subregion in an adult VgluT2-tdTomato-GAD67-GFP mouse, illustrating Cre-dependent VgluT2-tdTomato neuronal labeling (red), GAD67-knock-in GFP neuronal labeling (green), immunolabeling with glycine antibody (blue), and Sst antibody labeling (white) of pre-BötC neurons. Merged image illustrates that tdTomato-labeled VgluT2 neurons coexpress Sst (arrows), but GAD67-GFP- or glycine antibody-positive neurons do not express Sst (arrowheads). All images have the same dorsomedial anatomical orientation. d, Dorsal; m, medial.
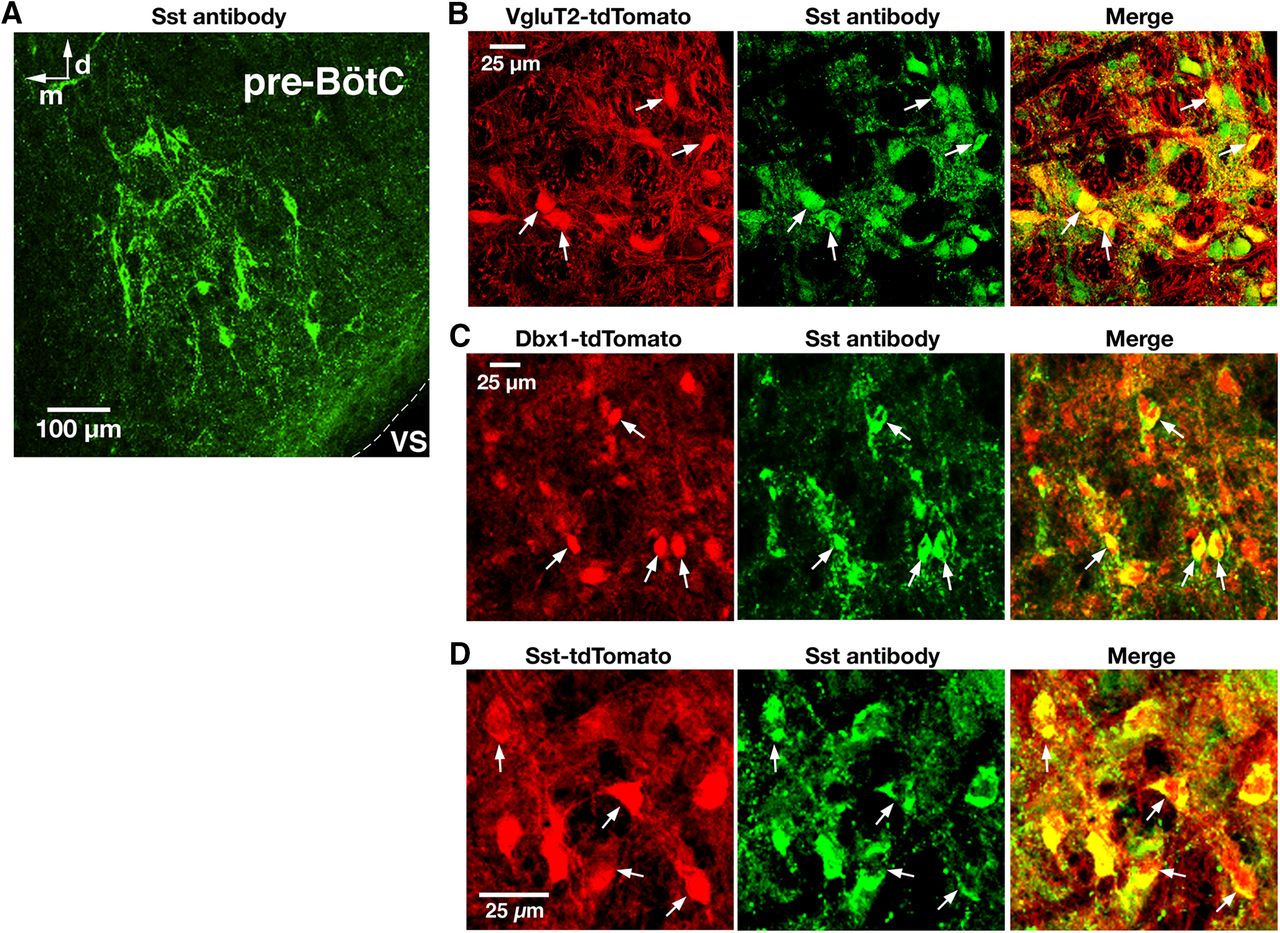
- Figure 5.
Validation of Sst-Cre line and Sst antibody labeling of pre-BötC glutamatergic and Dbx1-derived neurons. A, Confocal fluorescence microscopy images at low magnification showing Sst antibody-labeled neurons within the pre-BötC in a representative fixed coronal section from adult VgluT2-tdTomato mouse. VS, Ventral surface; d, dorsal; m, medial. B, C, Confocal fluorescence microscopy images of a pre-BötC subregion at higher magnification showing VgluT2 Cre-dependent tdTomato neuronal labeling (red; B), Dbx1 Cre-dependent tdTomato labeling (red; C), and immunolabeling with Sst antibody (green) in pre-BötC neurons. Arrows in the merged image in B and C indicate the coexpression of Sst in tdTomato-labeled VgluT2-expressing and Dbx1-derived neurons. D, Confocal images of a pre-BötC subregion in a fixed section from the Sst-tdTomato double-Tg mouse strain illustrating tdTomato neuronal labeling (red) and immunolabeling with Sst antibody (green) colocalized with tdTomato (arrows and merged image). All images have the same dorsomedial anatomical orientation.
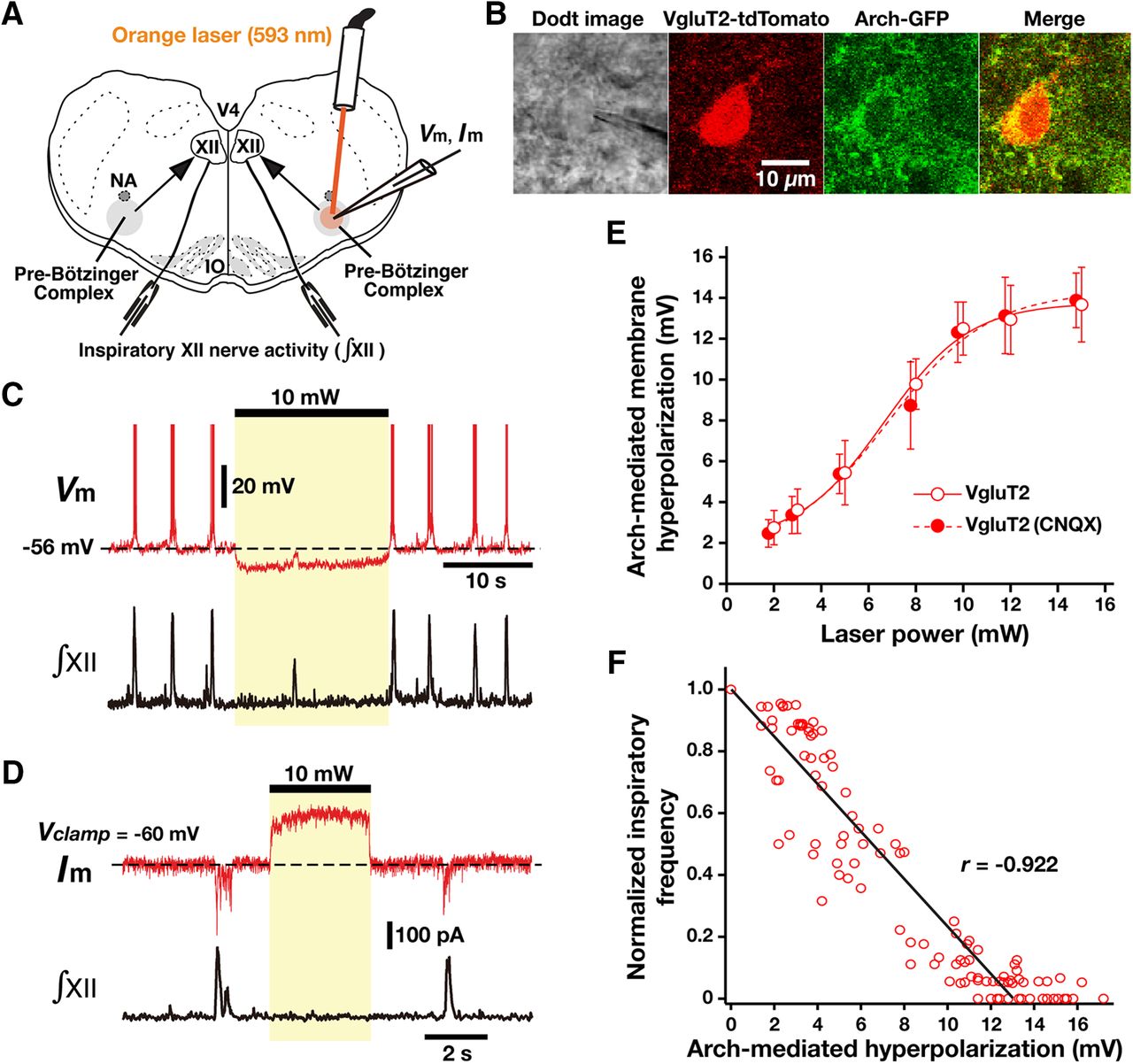
- Figure 6.
Optogenetic inhibition of rhythmically active Arch-expressing pre-BötC inspiratory glutamatergic neurons in vitro. A, Overview of experimental in vitro neonatal mouse rhythmic slice preparation with unilateral pre-BötC laser illumination (2-15 mW). NA, Nucleus ambiguus; V4, fourth ventricle. B, Two-photon single-optical plane images of pre-BötC glutamatergic neuron targeted for whole-cell recording, showing a Dodt structural image, VgluT2-Cre-mediated tdTomato labeling, and Arch-GFP expression (coexpression confirmed in merged image). C, Current-clamp recording from pre-BötC inspiratory neuron in B illustrates inspiratory bursting synchronized with integrated inspiratory hypoglossal activity (∫XII). The membrane potential was hyperpolarized by ∼10 mV at 10 mW laser power. This unilateral illumination was sufficient to reduce the frequency and amplitude of ∫XII. D, Under voltage-clamp, the same neuron as in C exhibited rhythmic inward synaptic currents synchronized with ∫XII, whereas laser illumination (10 mW) induced outward currents of ∼200 pA. E, Summary data showing Arch-mediated hyperpolarization in VgluT2-expressing pre-BötC inspiratory neurons (n = 22) was laser power dependent. After blocking fast glutamatergic synaptic transmission with CNQX, laser-induced hyperpolarization of VgluT2-positive pre-BötC inspiratory neurons (n = 6) was also laser power dependent, which was not significantly different from the cases with the synaptically coupled network (unpaired t test, p = 0.938). F, Linear correlation between laser-induced hyperpolarization of VgluT2-positive pre-BötC inspiratory neurons (n = 22) and XII inspiratory frequency, showing the monotonic reduction of frequency as a function of membrane hyperpolarization. Data are represented as the mean ± SD.
- Figure 7.
Rhythmically active Dbx1- and Sst-expressing pre-BötC inspiratory neurons identified by Ca2+ activity fluorescence imaging and whole-cell patch-clamp recordings. A1, Two-photon single optical plane live images of the pre-BötC region in an in vitro neonatal medullary slice from Dbx1-tdTomato Tg mouse line, showing Cre-driven tdTomato reporter protein labeling in Dbx1-derived neurons (red) and Ca2+-sensitive fluorescent dye (Oregon Green BAPTA-1 AM, OGB-1AM)-labeled neurons (green). Merged image shows tdTomato-labeled Dbx1-derived neurons colabeled with OGB-1AM by local microinjection. NA, Nucleus ambiguus; d, dorsal; m, medial. A2, Left, Example of rhythmic fluorescence intensity signals from Dbx1-derived pre-BötC inspiratory neuron (A1, arrow) illustrating Ca2+ transients (top trace) synchronized with electrophysiological signals of inspiratory network activity represented by integrated inspiratory hypoglossal activity (∫XII; bottom trace). Whole-cell current-clamp recording from the same neuron confirms neuronal activity in phase with the ∫XII (right). B1, Two-photon single-optical plane live images of the pre-BötC region in an in vitro neonatal medullary slice from Sst-tdTomato Tg mouse line, showing Cre-driven tdTomato reporter protein labeling of Sst neurons (red) and Ca2+-sensitive fluorescent dye (OGB-1AM)-labeled neurons (green). Merged image shows tdTomato-labeled Sst-expressing neurons colabeled with OGB-1AM by local microinjection. NA, Nucleus ambiguus; d, dorsal; m, medial. B2, Left, Example of rhythmic fluorescence intensity signals from Sst-expressing pre-BötC inspiratory neuron (A1, arrow) with Ca2+ transients (top trace) synchronized with electrophysiological signals of inspiratory network activity represented by ∫XII (bottom trace). Right, Inspiratory bursting activity of the same Sst-expressing pre-BötC inspiratory neuron synchronized with ∫XII under whole-cell current-clamp recording.
- Figure 8
Arch-mediated hyperpolarization in VgluT2-, Dbx1-, and Sst-positive pre-BötC inspiratory neurons. Summary data showing Arch-mediated hyperpolarization in VgluT2-positive (n = 22 from Fig. 6), Dbx1-positive (n = 28), and Sst-positive (n = 19) pre-BötC inspiratory neurons were laser power dependent, and the data fits with a logistic function were not significantly different among these neurons (one-way ANOVA: xhalf: F(2,56) = 0.45, p = 0.64; slope: F(2,56) = 0.74, p = 0.48).
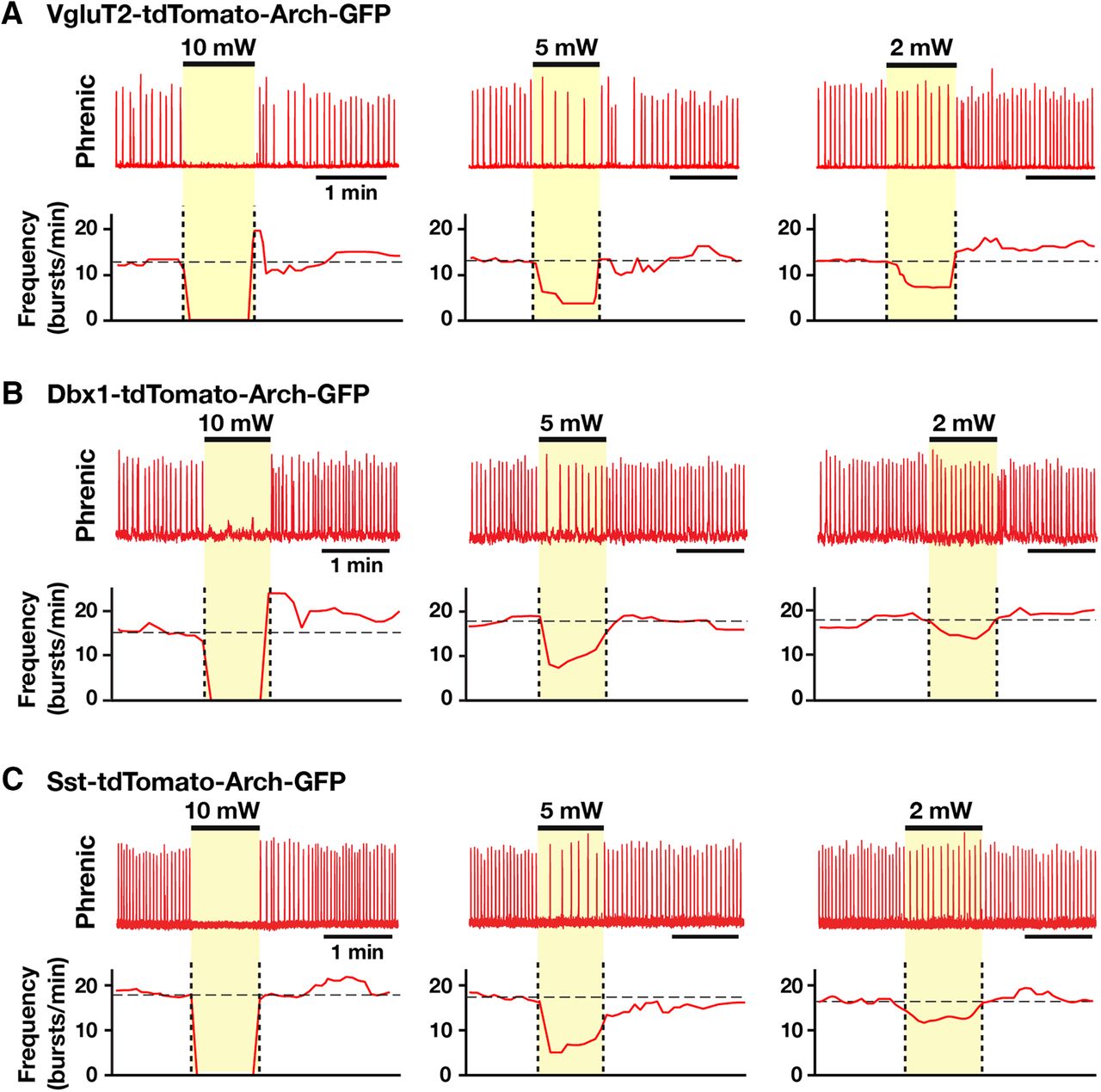
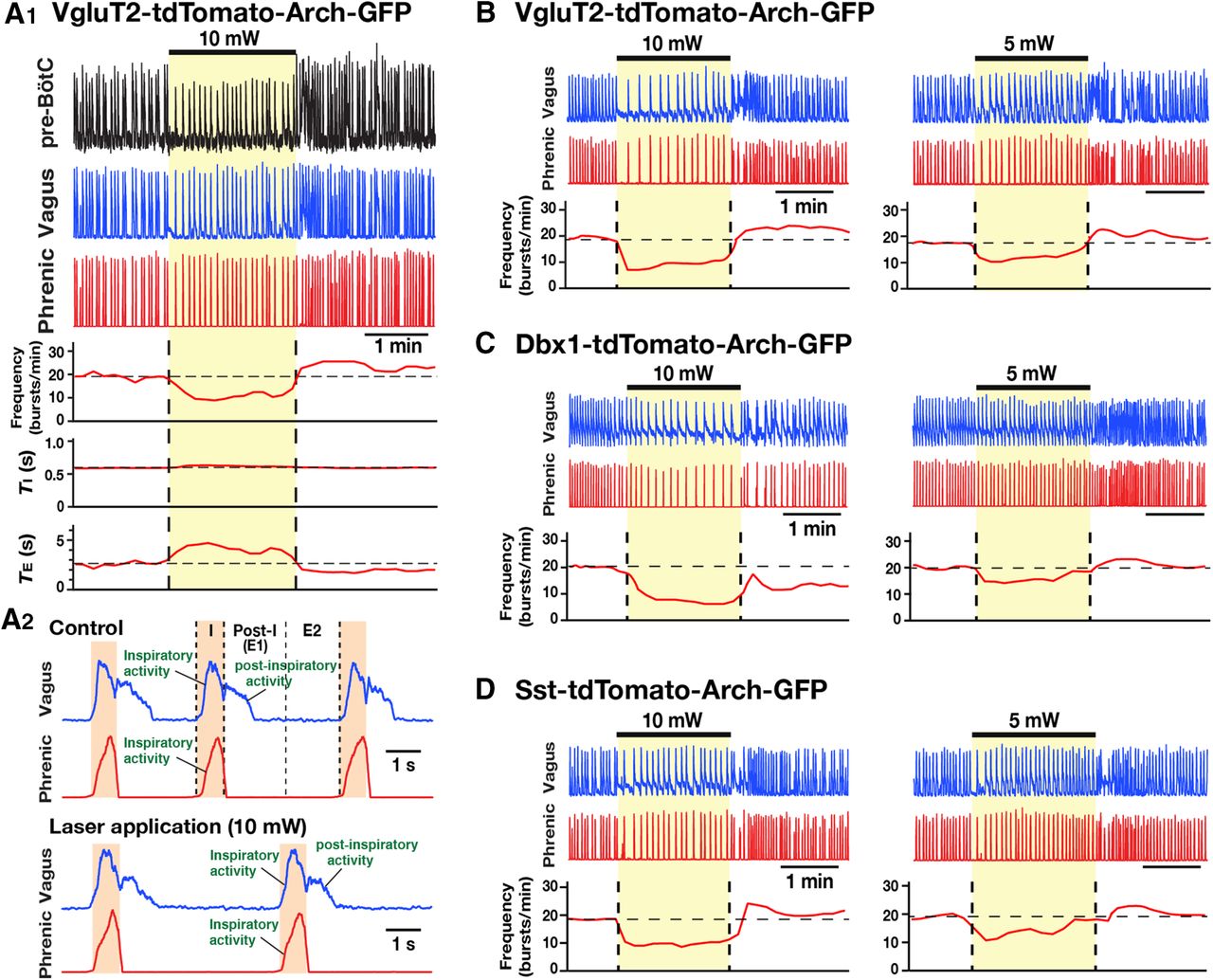
- Figure 9.
Population-specific Arch-mediated optical perturbations and inhibition of inspiratory rhythm in rhythmically active neonatal mouse in vitro medullary slice preparations. A–C, Experiments from VgluT2-tdTomato-Arch-GFP (A), Dbx1-tdTomato-Arch-GFP (B), and Sst-tdTomato-Arch-GFP (C) triple-Tg mouse lines. To systematically analyze the relationship between steady-state frequency and laser power (2-10 mW), we applied single epochs of continuous orange (593 nm) laser illumination (1-2 min). Representative traces of inspiratory network activity monitored by integrated XII nerve (∫XII) recordings, illustrating laser-induced perturbations of inspiratory frequency during bilateral pre-BötC illumination (2, 5, and 10 mW). In all cases, laser application caused rapid and reversible reductions of inspiratory frequency (time-based moving median, solid red lines in panels below the ∫XII traces in A, B, and C) in a laser power-dependent manner with the complete cessation of the inspiratory rhythm at the maximum applied power (10 mW).
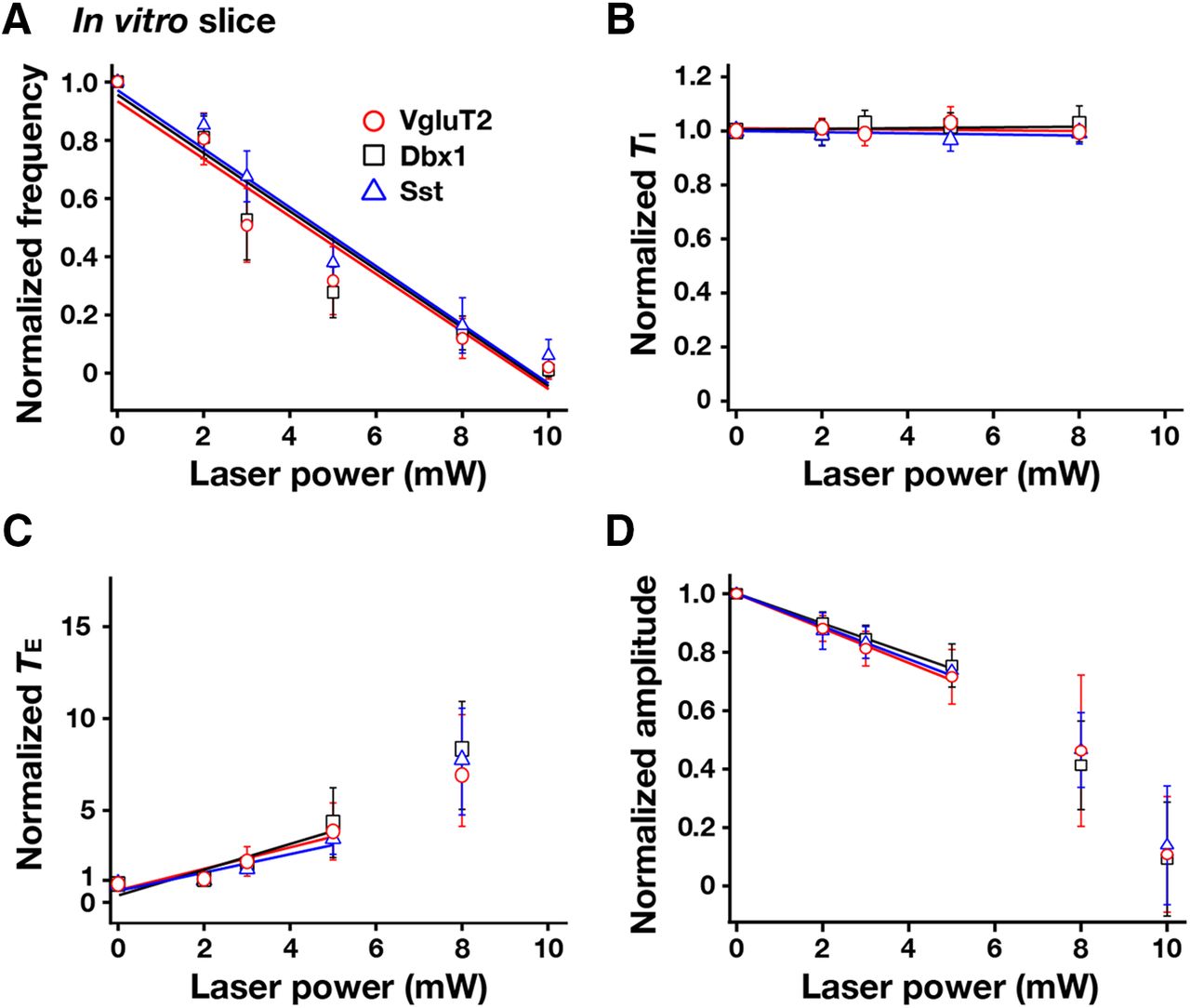
- Figure 10.
Summary of bilateral pre-BötC optical perturbations of inspiratory frequency, amplitude of inspiratory XII activity, and TI and TE in rhythmically active in vitro slice preparations from VgluT2-Arch-GFP (n = 12), Dbx1-Arch-GFP (n = 10), and Sst-tdTomato-Arch-GFP (n = 10) Tg lines. A, Laser application significantly reduced inspiratory frequency monotonically in a laser power-dependent manner in all cases. These frequency–power relations were not statistically different among VgluT2-, Dbx1-, and Sst-expressing neuronal populations (one-way ANOVA: F(2,29) = 0.71, p = 0.499). B, TI did not significantly change during laser application (F(3,30) = 1.72, p = 0.182). C, TE increased in a laser power-dependent manner, which accounts for the progressive reductions of inspiratory frequency. D, The amplitude of XII inspiratory bursts was reduced monotonically as a function of laser power compared with the control mice (data not shown; F(3,30) = 235.77, p < 0.0001; Tukey’s test, p < 0.01), but these relations (over the power range with nonzero frequency values, see regression lines in D) were not statistically different among the three Arch-expressing lines (Tukey’s test, nonsignificant). Data are represented as the mean ± SD.
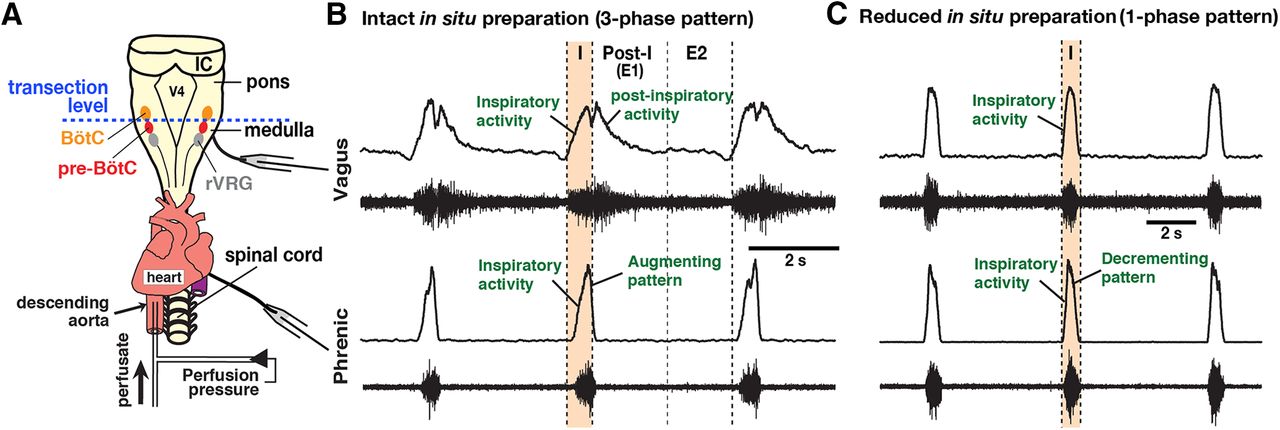
- Figure 11.
Representative activity patterns of central vagus and phrenic nerves recorded from the intact and the reduced in situ arterially perfused brainstem-spinal cord preparations of adult VgluT2-tdTomato-Arch-GFP Tg mice. A, Dorsal view of the intact in situ perfused brainstem-spinal cord preparations, also showing the transection level at the rostral boundary of the pre-BötC to obtain the reduced in situ preparations. V4, Fourth ventricle; IC, inferior colliculus. B, The intact in situ preparations generate a normal three-phase rhythmic pattern with inspiratory (I), postinspiratory (E1), and later expiratory (E2) phases. Each panel shows raw (bottom trace) and integrated (top trace) vagus and phrenic motor nerve discharge. The vagus nerves exhibit robust postinspiratory activity as well as inspiratory activity starting before phrenic inspiratory activity. Phrenic inspiratory discharges have an augmenting pattern. C, The physically reduced in situ preparations generate a reduced respiratory pattern with one (active)-phase inspiratory rhythmic motor output, generate no postinspiratory activity in the vagus nerve, exhibit the onset of inspiratory activity coincident with the onset of phrenic inspiratory activity, and generate phrenic inspiratory discharges that are converted to a decrementing neonatal slice-like pattern in the adult system.
- Figure 12.
Population-specific optogenetic inhibition of pre-BötC glutamatergic, Dbx1-derived, or Sst-expressing neurons in adult brainstem-spinal cord preparations in situ. A–D, Experiments from VgluT2-tdTomato-Arch-GFP (A, B), Dbx1-tdTomato-Arch-GFP (C), and Sst-tdTomato-Arch-GFP (D) mouse lines. A1, Representative traces of simultaneous extracellular recordings of integrated pre-BötC inspiratory neuronal population activity (top trace), as well as integrated inspiratory and postinspiratory activities recorded from vagus nerve (middle trace; represented in A2 with expanded time base) and inspiratory activities from phrenic nerves (bottom trace), with laser-induced effects on inspiratory frequency and TI and TE (solid red lines; time-based moving median). Bilateral pre-BötC illumination (10 mW, 2 min) caused a rapid and reversible reduction (∼50% in this example) of the frequency of integrated inspiratory activity. The decrease in frequency was due to the prolongation of TE without significant changes of TI. Simultaneous extracellular recordings of pre-BötC inspiratory population activity showed perturbations of inspiratory frequency. A2, The three-phase organization of the respiratory pattern [with inspiratory, postinspiratory (E1), and later expiratory (E2) phases] was not perturbed during bilateral pre-BötC illumination (bottom trace), when compared with the organization before laser application (top trace). B–D, Representative traces of inspiratory and postinspiratory vagus activities, and inspiratory phrenic activity with laser-induced perturbations of inspiratory frequency during bilateral pre-BötC laser illumination (5 and 10 mW). Inspiratory frequency was reduced significantly in a laser power-dependent manner in all cases. The frequency of inspiratory activity was maximally reduced by ∼50% in all cases at 10 mW, but was not eliminated in these in situ preparations.
- Figure 13.
Summary of bilateral pre-BötC optical perturbations of inspiratory frequency, amplitude of inspiratory phrenic activity, and TI and TE in the intact in situ preparations from VgluT2-Arch-GFP, Dbx1-Arch-GFP, and Sst-tdTomato-Arch-GFP (n = 8 each) Tg lines. A, Laser application significantly reduced inspiratory frequency monotonically in a laser power-dependent manner in all cases. These frequency–power relations were not statistically different among VgluT2-, Dbx1-, and Sst-expressing neuronal populations (one-way ANOVA: F(2,21) = 1.51, p = 0.246). B, TI did not significantly change (F(3,22) = 0.69, p = 0.568). C, TE increased in a laser power-dependent manner in all cases, which accounts for the progressive reductions in inspiratory frequency. D, The amplitude of phrenic inspiratory bursts was not significantly changed during laser application (F(3,22) = 0.89, p = 0.462). Data are represented as the mean ± SD.
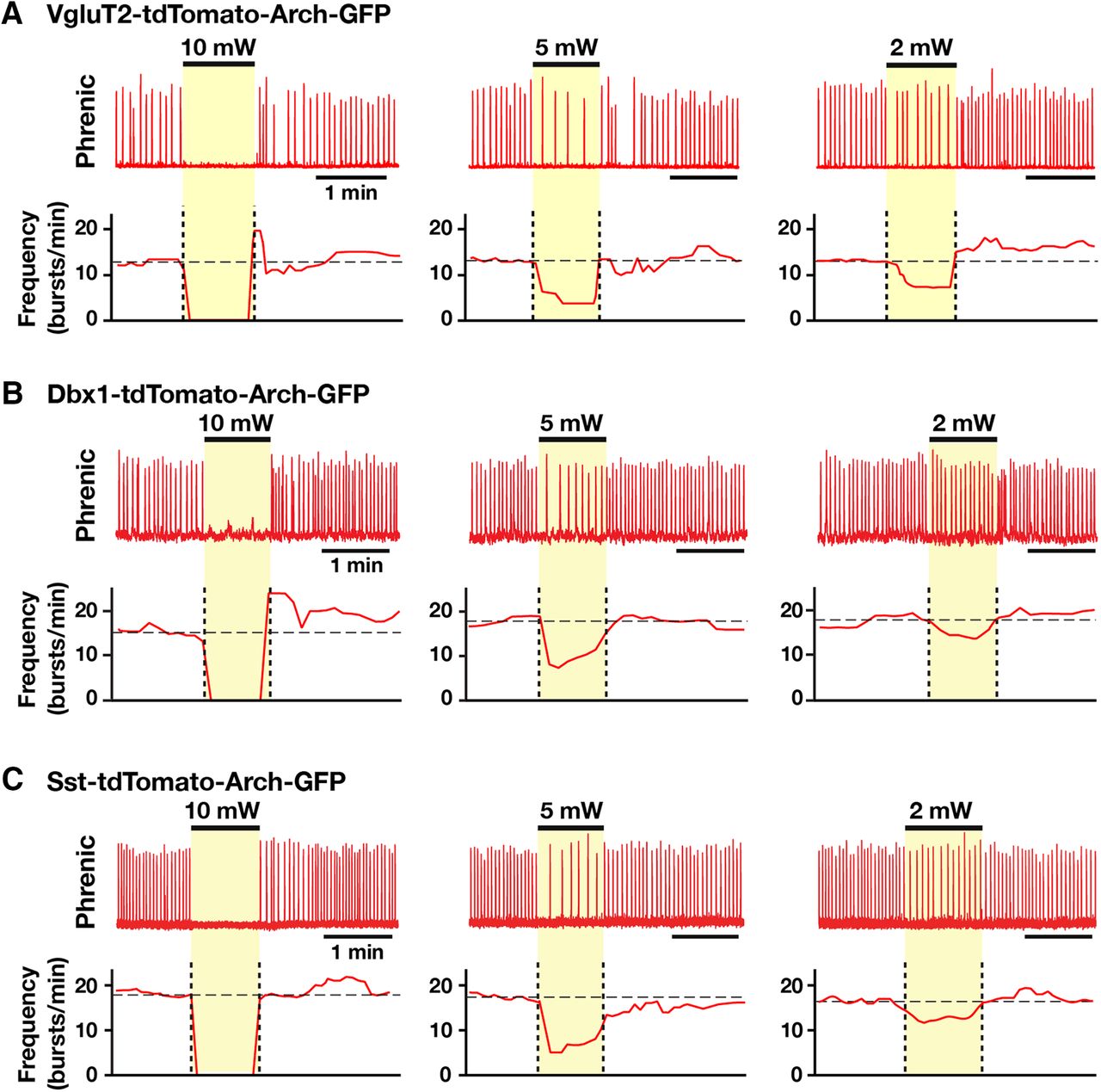
- Figure 14.
Population-specific Arch-mediated inhibition of pre-BötC glutamatergic, Dbx1-derived, and Sst-expressing neurons in the reduced in situ brainstem-spinal cord preparations. A–C, Experiments from adult VgluT2-tdTomato-Arch-GFP (A), Dbx1-tdTomato-Arch-GFP (B), and Sst-tdTomato-Arch-GFP (C) mouse lines. The reduced in situ preparation, with transection at the rostral boundary of the pre-BötC, generates a reduced respiratory pattern that is represented by phrenic nerve activity with one (active)-phase (inspiratory but no postinspiratory phase activity; Fig. 11C) and is similar to that generated in the in vitro neonatal slice preparations. Representative traces of integrated phrenic activities and laser-induced perturbations of inspiratory frequency during bilateral pre-BötC laser illumination (2, 5, and 10 mW), which caused a rapid and reversible laser power-dependent reduction of the inspiratory frequency with complete cessation of the inspiratory rhythm at the maximum applied power (10 mW).
- Figure 15.
Summary of bilateral pre-BötC optical perturbations of inspiratory frequency, the amplitude of inspiratory phrenic activity, and TI and TE in the reduced in situ preparations from VgluT2-Arch-GFP, Dbx1-Arch-GFP, and Sst-tdTomato-Arch-GFP (n = 6 each) Tg lines. A, Laser application significantly reduced inspiratory frequency monotonically in a laser power-dependent manner in all cases. These frequency–power relations were not statistically different among VgluT2-, Dbx1-, and Sst-expressing neuronal populations (one-way ANOVA: F(2,15) = 0.60, p = 0.564). B, TI did not significantly change (F(3,16) = 0.56, p = 0.649). C, TE increased in a laser power-dependent manner, which accounts for the progressive reductions of inspiratory frequency. D, The amplitudes of phrenic inspiratory bursts were not significantly changed (F(3,16) = 0.66, p = 0.589). Data are represented as the mean ± SD.
- Figure 16.
Site specificity in the ventrolateral respiratory column of laser-induced perturbations of the inspiratory rhythm in the intact in situ preparations from VgluT2-tdTomato-Arch-GFP mice. A, Representative traces of integrated phrenic inspiratory activity (top trace) illustrating a reduction of inspiratory burst amplitude without perturbations of inspiratory frequency by laser illumination (10 mW) in the rVRG region ∼200 µm caudal to the caudal boundary of the pre-BötC. B, Summary data from five in situ preparations showing that, in contrast to when orange light is applied to the pre-BötC region, laser application to the rVRG (2-10 mW) did not perturb inspiratory frequency compared with the frequency before laser application (paired t test, p = 0.607 at 10 mW), but significantly reduced the amplitude of integrated phrenic inspiratory activity compared with averaged amplitudes before application of the laser (paired t test: 2 mW, p = 0.016; 5 mW, p = 0.0012; 10 mW, p = 0.00017), as predicted for the partial inhibition of rVRG bulbospinal excitatory inspiratory neurons. These results indicate that laser illumination specifically in the pre-BötC is required for perturbations of the inspiratory rhythm. Data are represented as the mean ± SD.











Tables
Figure Data structure Type of test p value 6E (xhalf) Normally distributed Student’s t test 0.904 6E (slope) Normally distributed Student’s t test 0.938 8 (xhalf) Normally distributed One-way ANOVA 0.640 8 (slope) Normally distributed One-way ANOVA 0.480 10A Normally distributed One-way ANOVA 0.499 10B Normally distributed One-way ANOVA 0.182 10C Normally distributed One-way ANOVA 0.512 10D Normally distributed One-way ANOVA 0.448 13A Normally distributed One-way ANOVA 0.246 13B Normally distributed One-way ANOVA 0.568 13C Normally distributed One-way ANOVA 0.288 13D Normally distributed One-way ANOVA 0.462 15A Normally distributed One-way ANOVA 0.564 15B Normally distributed One-way ANOVA 0.649 15C Normally distributed One-way ANOVA 0.594 15D Normally distributed One-way ANOVA 0.589 16B Frequency, 2 mW Normally distributed Student’s t test 0.891 Frequency, 5 mW Normally distributed Student’s t test 0.482 Frequency, 10 mW Normally distributed Student’s t test 0.607 Amplitude, 2 mW Normally distributed Student’s t test 0.016 Amplitude, 5 mW Normally distributed Student’s t test 0.0012 Amplitude, 10 mW Normally distributed Student’s t test 0.00017
In this issue

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}



