

Article Figures & Data
Figures
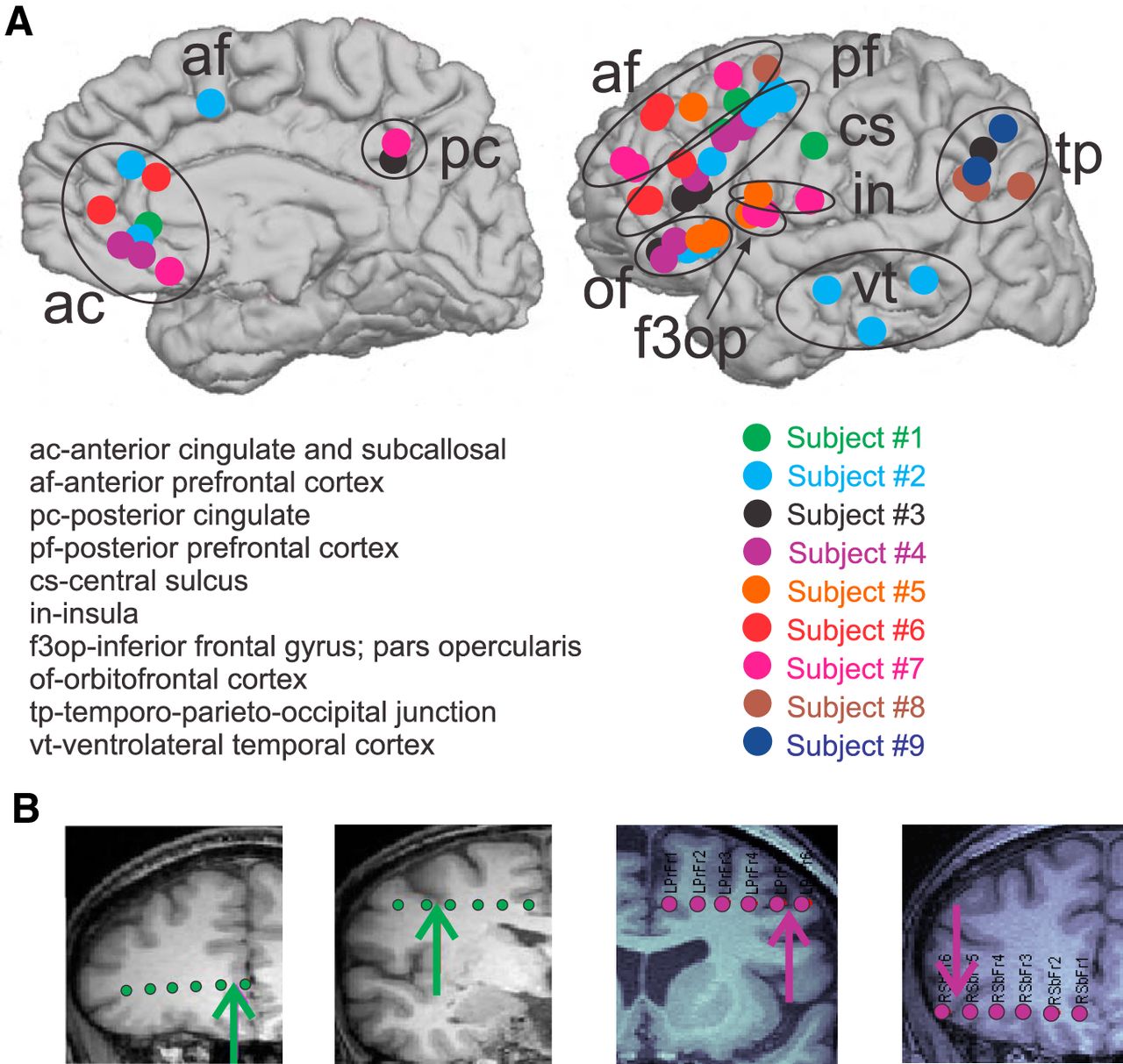
- Figure 1.
Bipolar SEEG contacts. A, Bipolar SEEG contacts, color coded by subject, are marked by circles on the medial (left) and lateral (right) surfaces of the brain. Bipolar SEEG contacts are grouped into 1 of 10 anatomical groups, as demarcated by black circles and/or one of the following labels: ac (anterior cingulate and subcallosal); af (anterior prefrontal cortex); pc (posterior cingulate); pf (posterior prefrontal cortex); cs (central sulcus); in (insula); f3op (inferior frontal gyrus; pars opercularis); of (orbitofrontal cortex); tp (temporo-parieto-occipital junction); vt (ventrolateral temporal cortex). B, Example SEEG electrodes for subject 1 (green) and subject 4 (purple) are superimposed on the subjects’ MRIs. The arrows indicate the location of the bipolar contact pair chosen on each electrode. From the MRI, it is possible to see that the two contacts in each pair are spanning the local cortical mantle.
- Figure 2.
Example KC localizations, waveforms, and down state confirmations. KCs are plotted from representative sites across the cortical areas sampled. Individual KCs for each bipolar location are plotted in black with the average KC overlaid in red. The number of individual KCs plotted is listed above the waveforms. Below the KC waveforms, time frequency plots from 5 to 120 Hz with a −1.5 to −0.5 s baseline relative to the most negative peak of the KC at time zero indicate a significant drop (p < 0.01, uncorrected) in high gamma power at the time of the KC. Arrows indicate the bipolar locations where the plotted waveforms and time–frequency plots were recorded. Circles indicate the subject color-coded bipolar channel locations, as in Figure 1. The four additional red squares indicate sites where KCs were located, but not included for additional analysis because they were part of an epileptic region or potentially in entorhinal cortex.
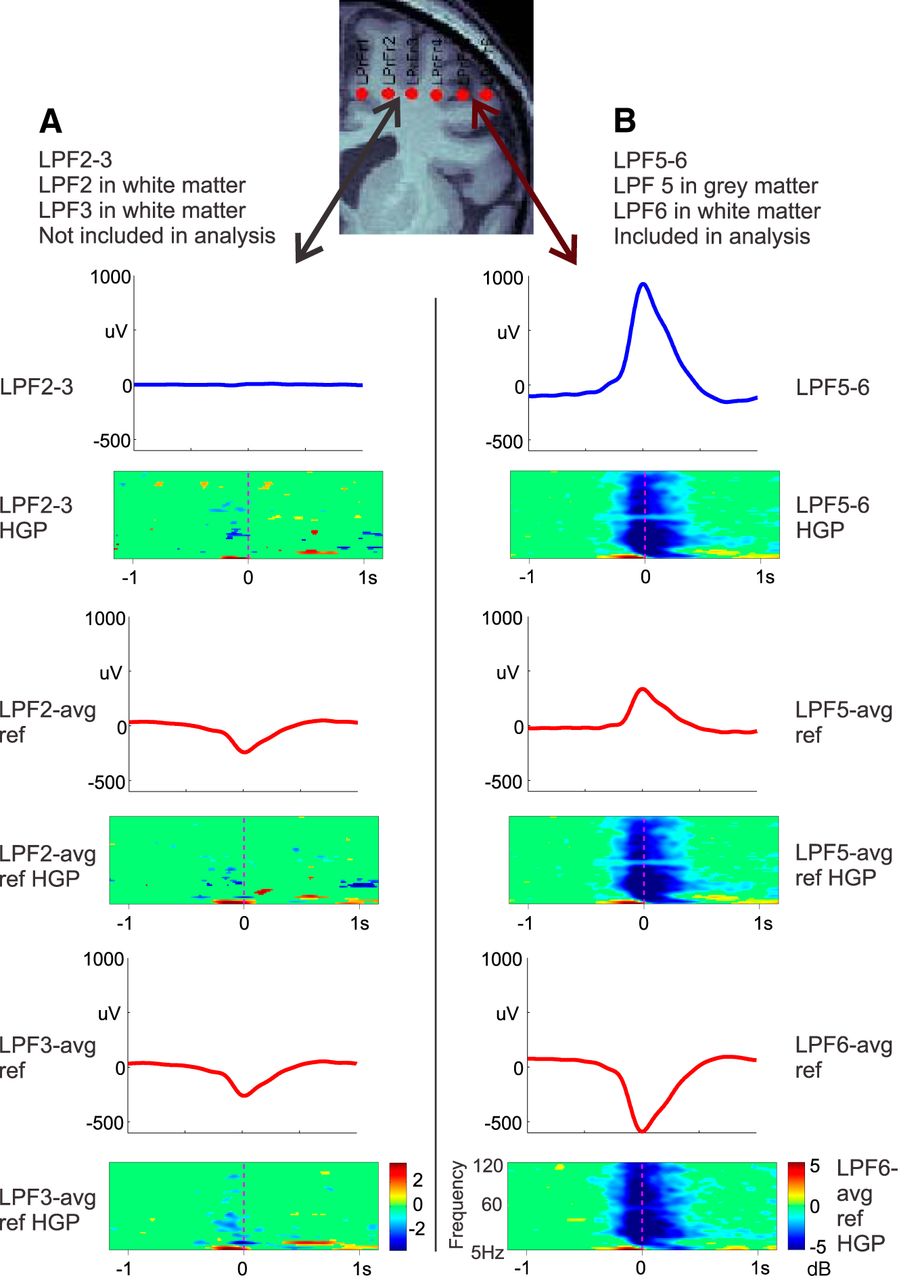
- Figure 3.
Bipolar versus common average reference recordings. Bipolar KC recordings (blue waveforms) were compared with monopolar KC recordings (red waveforms) referenced to a common average reference (T3, T5, F3, C3, P3, O1, F4, C4, P4, O2, T4, and T6) as in Wennberg (2010). Two pairs of adjacent contacts located on the same electrode of subject 4 were examined. A, Bipolar (blue) and referential (red) recordings from two adjacent contacts, both located in the white matter, as confirmed by MRI. Bipolar recordings show no KCs, but referential recordings do, despite the fact that no decrease in high gamma power is present in the time–frequency plots below each waveform. Thus, referential recordings show KCs even when they are not locally generated. B, Bipolar (blue) and referential (red) recordings from two adjacent contacts that span the gray matter, as confirmed by MRI. Both bipolar and referential recordings show KCs, together with decreased high gamma power. All recordings were averaged with respect to the peaks of KCs identified in the bipolar recordings in B.
- Figure 4.
KC amplitudes vary across the cortex. A, The average amplitude of manual KCs at each channel, color coded by subject, is highly variable across the cortex. However, no significant effects are found when a linear mixed-effects model is applied. B, The addition of KC-like activity shows a similar pattern, but with the increased sample, the same linear mixed-effects model now finds that the anterior prefrontal channels highlighted by the blue circle are ∼30% smaller than the orbitofrontal reference (p = 0.045). Black circles in the top right indicate the amplitude scale represented by the size of each circle.
- Figure 5.
KC occurrence rates vary across the cortex. A, Manually marked KCs color coded for each subject. The size of the circle indicates the KC occurrence rate at each location, normalized to the maximum KC occurrence rate seen on any channel for that subject. Black circles in the top right indicate this normalized occurrence rate scale. Locations identified by a linear mixed-effects model to have a significantly higher occurrence rate than the orbitofrontal reference are highlighted by pink circles. Locations identified as occurring significantly less frequently are highlighted by blue circles. B, KC-like activity in addition to manually marked KCs. The locations showing a lower occurrence rate are the same as in A. The locations showing a higher occurrence rate differ with the addition of the template-detected KCs.
- Figure 6.
Observed manual KCs versus those expected under the null hypothesis of independent occurrence, for representative nights for each subject. A, KC events observed for each subject were classified into two groups: KCs with one participating channel (light orange); and KCs with two or more participating channels (dark orange). B, KC events derived from simulating an expected distribution for each subject were classified into the same two groups: one channel (light purple); and two or more channels (dark purple). For all subjects, there are more KCs that occur only in one channel expected under the null hypothesis than were actually observed. These distributions are tested statistically using a Fisher’s exact test, and these results are listed in Table 3.
- Figure 7.
Lack of systematic sequential order in KCs. Sequence pairs that are significant with manually marked KCs are plotted with red arrows, while sequence pairs that are significant only after the addition of KC-like activity are plotted with blue arrows. Sequence pairs tested but not significant are plotted in yellow. A, Subject 3 has electrodes in frontal and parietal cortices, but only shows significant sequences within the frontal lobe, and only after the addition of KC-like activity. B, Subject 2 shows a large number of significant sequence pairs (16) in manually marked KCs, the majority of which lead to the temporal lobe (11). This pattern is strengthened with the addition of KC-like activity (18 of 29 additional significant pairs). A variety of other significant sequences are seen within the frontal lobe.
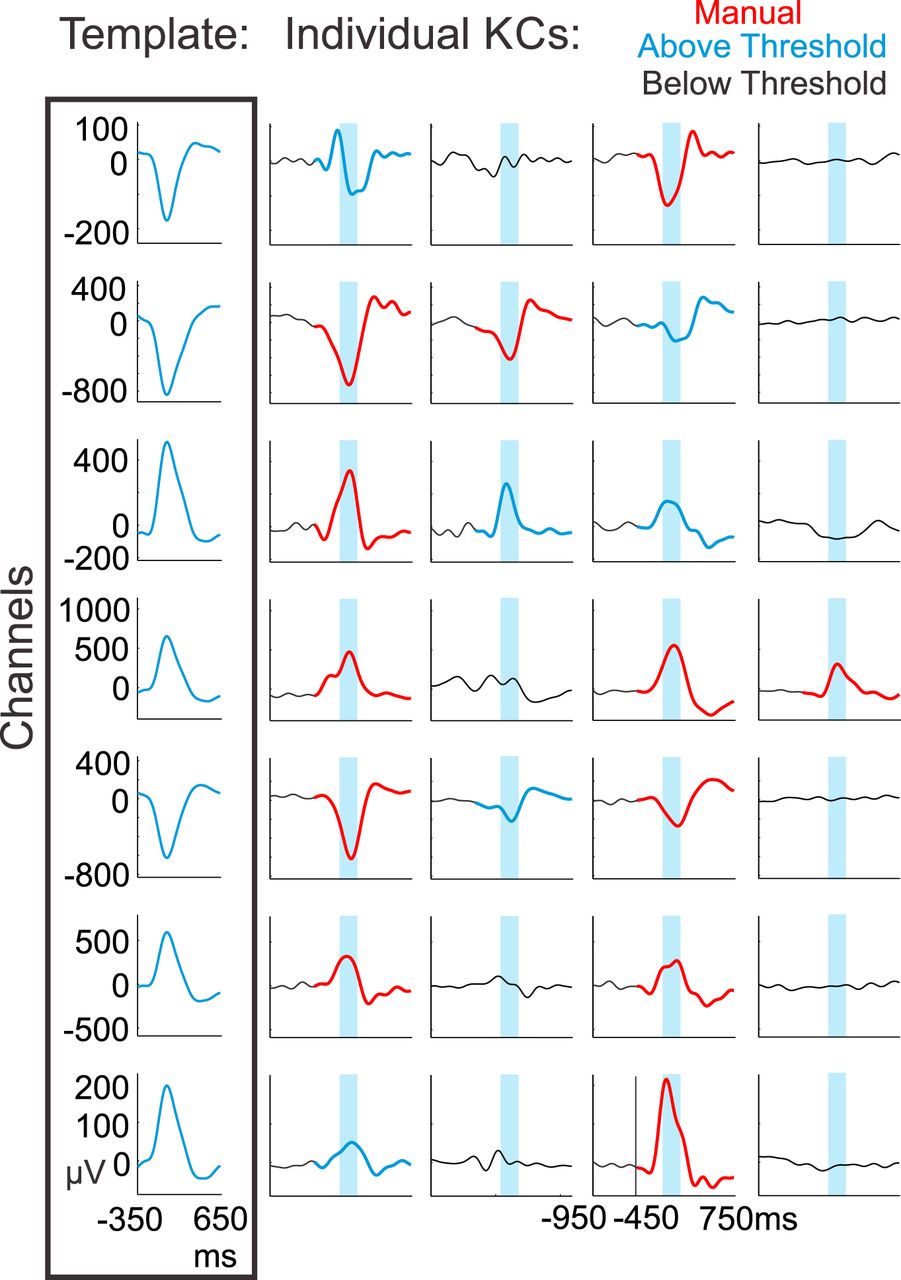
- Figure 8.
Template application to detect KC-like activity. Channel-specific templates were created for each subject by averaging from −350 to +650 ms on the most negative peak at time zero over the manually detected KCs in each channel (box at left). The channel-specific templates were then applied when a manually marked KC occurred in at least one channel. The templates were applied from −450 to +750 ms, with time zero representing the average time between the first and last manually chosen channel peaks within a KC. Four such examples from subject 6 are plotted vertically. Manually marked KCs are plotted in red. The maximum value of the sliding inner product between the signal and the template was taken over the blue highlighted window. If the value was above the 99th percentile of the null distribution for that channel and corresponded to the largest (or smallest) peak over the entire 1200 ms window, the KC was recorded as KC-like activity (KCs in blue), and if it was below the threshold, it was not (signal in black).
- Figure 9.
Manually detected KCs have smaller pre-KC period variance and larger amplitude than template-detected KCs. A, The probability density of variance in the pre-KC period (−1.4 to −0.4 s prior to peak) is plotted for the null times (black), manually detected KCs (red), and template-detected KCs (blue) for each of subject 1’s four channels. B, The KC peak amplitude probability density of null times (black), manually detected KCs (red), and template-detected KCs (blue) are plotted for the same four channels as in A. C, For each channel, the manually detected (red circles) and template-detected (blue triangles) z-scores were calculated for the KC peak amplitude and the pre-KC period variance referenced to the null values. A line connects the manually detected and template-detected values for each channel. A green line indicates that the amplitude of template-detected KCs is smaller than that of manually detected KCs in that channel; a black line indicates the opposite relationship. On the right is an expansion of the lower left corner, which is outlined by a gray box.
- Figure 10.
Including KC-like activity increases multichannel KC participation. The addition of KC-like activity (blue columns) to the manually marked KCs (red columns) increased the number of channels that were participating in KCs for all subjects. A, In subjects where a small number of manually marked KCs included the participation of all channels, the addition of KC-like activity increased the percentage of such KCs. B, Subjects who did not show maximum channel participation with manually marked KCs displayed KCs that included all channels after the addition of KC-like activity. In subjects 2 and 6 (pictured), and subject 3 (not pictured), the classification of KCs using a 200 ms crawl window slightly shifted the distribution of manual multichannel participation to the right (gray columns), but not to the extent of the addition of KC-like activity (blue columns).
- Figure 11.
Anterior-to-posterior contact pair distance versus KC latency delay. A, The distance between anterior-to-posterior channel pairs was plotted as a function of the average KC latency difference between the two channels, for all contact pairs (black circles and red triangles). The linear mixed-model regression between anterior-to-posterior distance and delay for all pairs is plotted as a purple line (p = 0.007), and, after excluding the temporal pairs (plotted with red triangles), is plotted as a green line (p = 0.08). B, The distance between the same anterior-to-posterior channel pairs as in A was plotted as a function of each individual KC latency difference (black circles and red triangles). A linear mixed-model regression calculated to determine whether a significant relationship existed for individual KCs on these pairs was significant at p < 10−4 (purple line). Excluding the temporal pairs plotted with red triangles, the linear mixed-model regression was still significant at p = 0.003 (green line).
- Figure 12.
Comparison of N2 and N3 KCs. A, Waveforms of averaged N2 and N3 KCs overlaid for subject 1’s four channels exhibit highly similar KC morphology. B, Multichannel KC participation is not consistently greater in N2 or N3. The percentage of KCs occurring in each number of channels for N2 (red) and N3 (blue) are plotted for subjects 4, 6, and 9. Subjects 6 and 9 show N2 KCs involving more channels, but this is not the pattern seen in all subjects, as shown with subject 4. When a χ2 test was performed for each subject on these distributions, only subjects 6 (p = 0.011) and 9 (p = 0.000056) exhibited significant differences.
- Figure 13.
Summary of current KC findings compared with previous views. A, Previous scalp EEG and intracranial studies of the KC and isolated SOs (i.e., KCs) find the KCs with the largest amplitudes in midline prefrontal regions (orange star), as well as strong (blue arrow), one-directional (faded orange background) propagation from anterior to posterior cortical regions. B, Our current findings indicate that when measured locally, KCs are actually smallest in amplitude in anterior prefrontal regions (orange star). KCs may co-occur (uniform yellow background) across variably small or large areas of cortex (stars and arrows). Propagation may occur from anterior to temporal regions (gray arrow), but also occurs in a variety of patterns across the cortex (groups of colored arrows). This variability in KC onset, extent, and spread could be used by the cortex during replay to consolidate memories encoded with correspondingly variable spatiotemporal encoding patterns.












Tables
Subject Gender Age (years) Handedness Clinical diagnosis Pathologic diagnosis Imaging Focus IQ (FSIQ) 1 M 45 L CPS; bitemporal No pathology obtained Left and right mesial temporal lobes 83 2 F 58 R CPS; possible generalized epilepsy No pathologyobtained Likely generalized 80 3 F 45 R CPS; multifocal Multifocal: temporal, parietal, occipital Temporal Average 4 F 65 R CPS; temporal lobe epilepsy with two foci: left mesial temporal structures and right subfrontal region No pathology obtained Right subfrontal and anterior temporal 101 5 F 27 R Right temporal lobe epilepsy No pathology obtained Right hippocampal sclerosis Hippocampus, entorhinal cortex, amygdala Average 6 F 32 R Left temporal lobe epilepsy Non-specific gliosis Normal Hippocampus, entorhinal cortex, amygdala 77 7 F 23 R Right temporal lobe epilepsy Type I focal dysplasia Normal Hippocampus, entorhinal cortex,anterior insula 86 8 F 28 R Right temporal occipital epilepsy Type II focal dysplasia Focal cortical dysplasia in right fusiform Fusiform gyrus, entorhinal cortex Average 9 F 50 R Right temporal occipital epilepsy No pathology obtained Normal Right fusiform gyrus 110 FSIQ, Full Scale Intelligence Quotient; CPS, Complex Partial Seizures.
- Table 2.
Mean and SD of maximum KC peak differences between manually chosen KCs for each night for each subject
Subject Channels (n) Mean of maximum KC peak differences (ms) SD (ms) 1 4 61.1 49.5 2 13 99 59.6 3 5 86.3 59.5 4 7 66.3 54.1 5 N1 5 24.8 23.8 N2 5 27.8 26.4 N3 5 32.2 27.7 6 N1 7 71.3 48.5 N2 7 66.4 45.6 N3 7 81.2 52.8 N4 7 60.3 54 7 N1 8 68.6 59.1 N2 8 63.2 57.2 N3 8 73.4 61.2 N4 8 77.7 63.1 N5 8 70.7 51.1 N6 8 74.9 59.7 8 N1 4 49.8 53.4 N2 4 79.2 42 N3 4 67.5 59.7 9 N1 2 95.2 59.4 N2 2 115.2 0 N3 2 52.2 39 N4 2 78.6 83 - Table 3.
Fisher’s exact test for co-occurrence for one channel and two or more channels; three or more channels; four or more channels; and five or more channels for all nights for all subjects
Subject 1 Channel and ≥2 channels 1 Channel and ≥3 channels 1 Channel and ≥4 channels 1 Channel and ≥5 channels 1 < 2.2E-16 9.49E-12 0.000445 2 < 2.2E-16 < 2.2E-16 < 2.2E-16 < 2.2E-16 3 < 2.2E-16 < 2.2E-16 6.24E-09 0.00493 4 < 2.2E-16 < 2.2E-16 5.38E-08 5 N1 < 2.2E-16 N2 5.40E-14 N3 2.96E-13 0.0135 6 N1 < 2.2E-16 < 2.2E-16 < 2.2E-16 1.10E-15 N2 < 2.2E-16 < 2.2E-16 < 2.2E-16 < 2.2E-16 N3 < 2.2E-16 < 2.2E-16 < 2.2E-16 1.25E-15 N4 3.92E-10 0.0125 7 N1 < 2.2E-16 2.33E-06 N2 < 2.2E-16 1.20E-05 N3 < 2.2E-16 1.61E-05 0.0140 N4 < 2.2E-16 2.00E-08 N5 9.50E-06 N6 8.09E-08 8 N1 < 2.2E-16 0.000355 N2 0.00227 N3 0.000107 9 N1 0.215 N2 0.497 N3 0.000312 N4 0.0140 Bonferroni correction at p < 0.05 was applied across all tests. Values in bold are significant after this Bonferroni correction.
In this issue

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}



