

Article Figures & Data
Figures
- Figure 1
Acute effects of decentralization on the pyloric rhythm. A, Schematic diagram of the dissected STNS, as pinned out for recording, showing the location of the STG and its associated anterior modulatory ganglia (CoG, OG). Grey circles identify where Vaseline wells were built for monitoring pyloric activity extracellularly. Decentralization was accomplished by blocking or transecting the stn at the indicated location. B, Example lvn recordings from a preparation before (blue trace) and 30 min following decentralization (red trace). C, D, Histograms of pyloric cycle frequencies immediately before decentralization (C, blue bars) and 30 min after (D, red bars). E, F, Phase relationships among the PD and LP cells in the pyloric rhythm for intact (E, blue) and 30 min decentralized (F, red) preparations. Lines represent fits by linear regression.
- Figure 2
Long-term, continuous extracellular recordings of preparations of the STNS reveal stable behavior. Example 15 s lvn traces from two preparations, one in which with the stn left intact throughout the experiment (blue traces, left), and one in which the stn was transected shortly after the baseline recording (red traces, right).
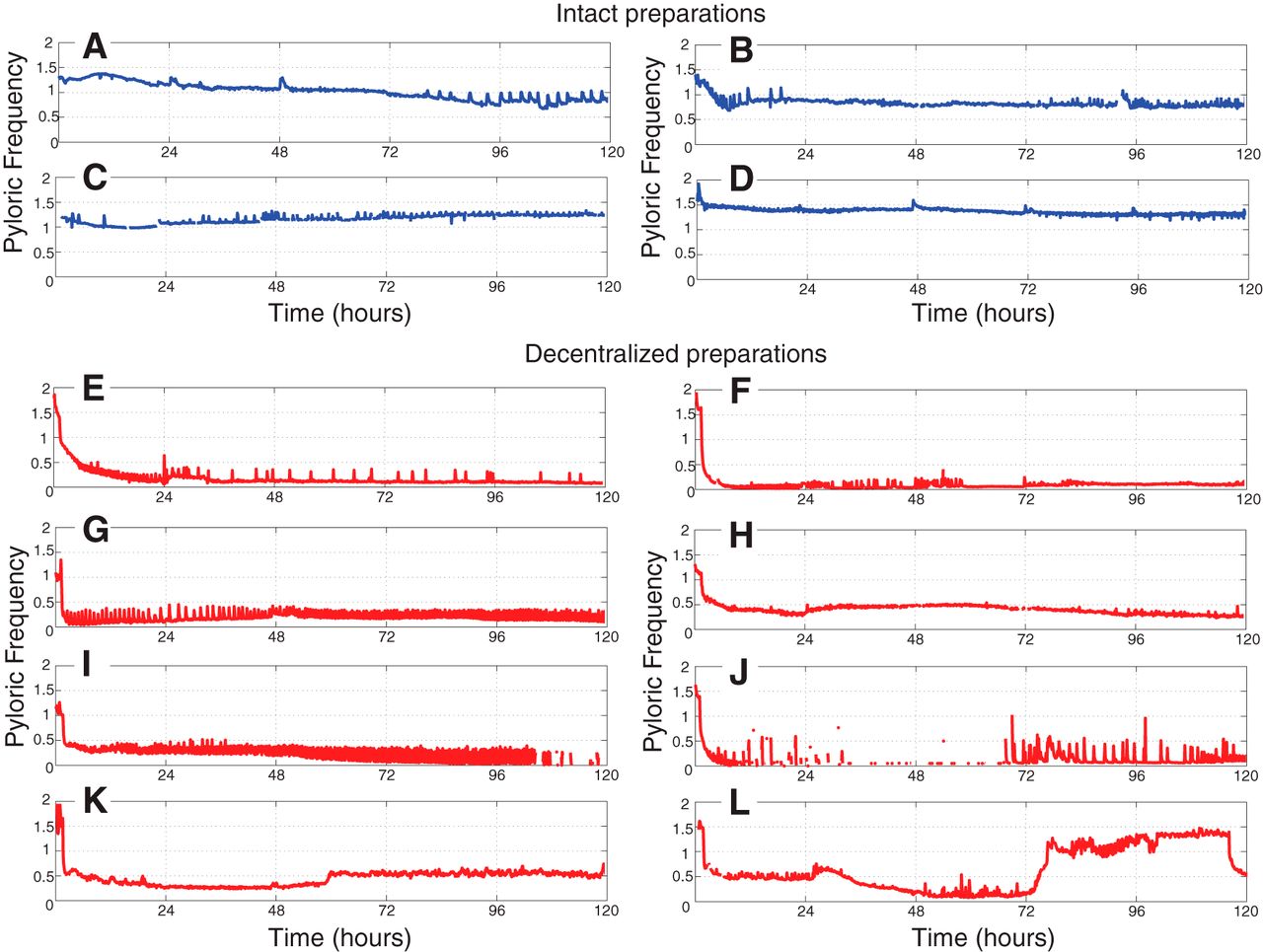
- Figure 3
Example long-term pyloric cycle frequency plots of intact (A−D) and decentralized (E−L) preparations of the STNS. Data are plotted after binning frequencies into 0.1 h bins. Intact preparations (blue) tend to maintain stable pyloric rhythms with frequencies in the normal range of controls. Decentralized preparations (red) display more varied and variable behavior. Decentralization was accomplished approximately 1 h following the beginning of these recordings.
- Figure 4
Long-term population effects of decentralization on the pyloric rhythm. A, Population data for intact (n = 9) and decentralized (n = 19) preparations is shown for the first 6 d in culture. Data are plotted as box plots (see Materials and Methods) for each hour; data from intact preparations are shown in blue while decentralized preparations are shown in red. B−E, Cycle frequency correlations within preparations across time (dc, decentralized). Black diagonal line represents unity. Insets report R and p values of Pearson correlations. B, Following decentralization, all preparations exhibit a lower pyloric cycle frequency. C, All examined preparations also exhibit a lower frequency on the second day following decentralization (average frequency, 24−48 h). D, E, On subsequent days, preparations are more correlated, and show an equal tendency to increase or decrease frequency. F−K, Measures of frequency (F), across-preparation variability (coefficient of variation (C.V.), G), within-preparation variability (H), and pyloric phase relationships (I−K) are plotted for intact (blue) and decentralized (red) preparations. Box plots are created from population data consisting of the average value for each preparation in 24 h bins following decentralization.
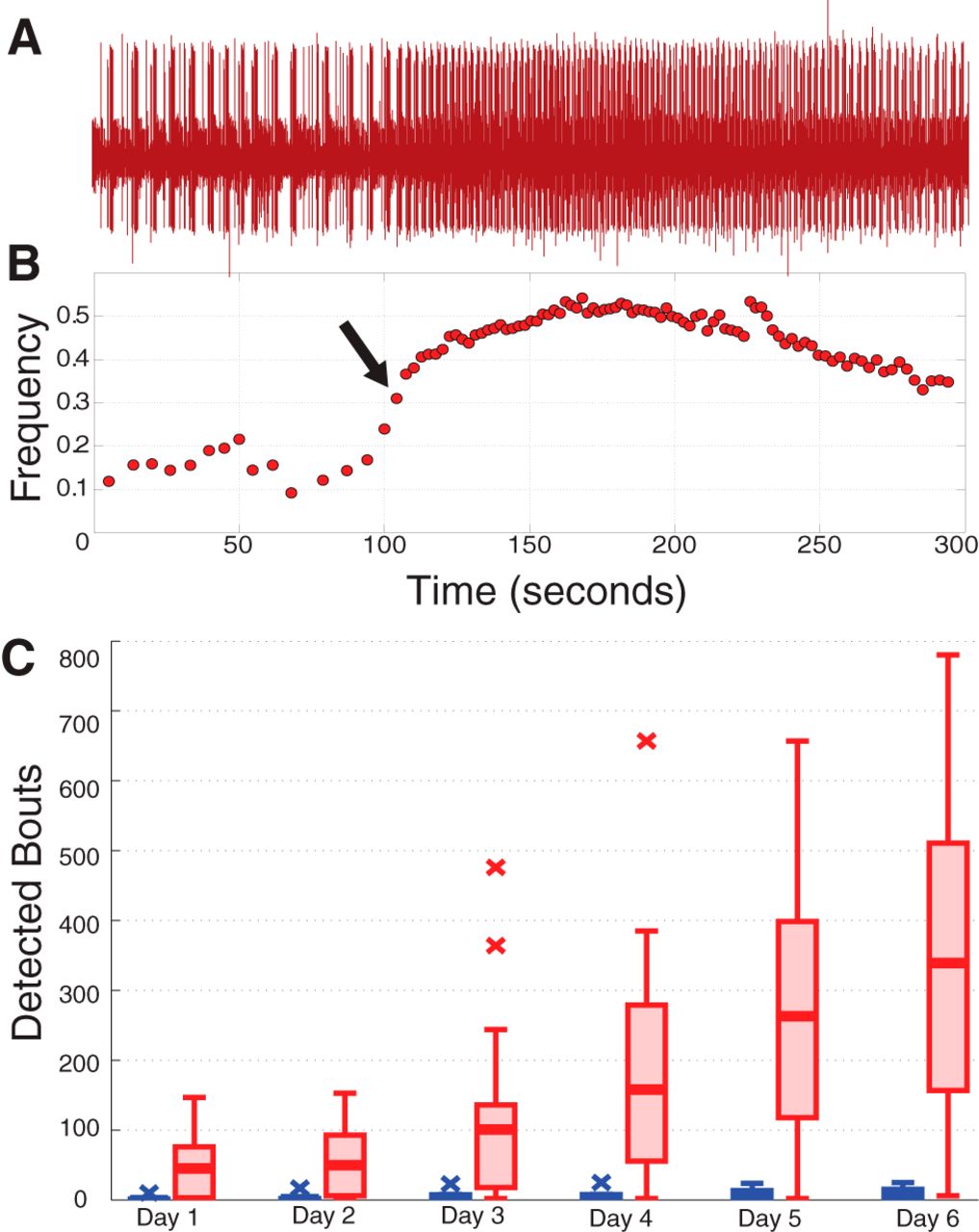
- Figure 5
Analysis of bouting. A, Example lvn recording featuring one detected bout, visible here in the noticeable increase in pyloric cycle frequency starting mid-trace. B, Frequency plot corresponding to the data in A. Arrow designates the point at which the bout was detected. C, Box plots show population data of the number of detected bouts by day for intact (blue) and decentralized (red) preparations.
- Figure 6
Analysis of previously used measures of recovery. A. Two previously described definitions of recovery (time to first bout, and 5 h strong and steady rhythm) fail to correlate. B−F, Box plots show population data of intact (blue), 30 min decentralized (dc; red), and the fastest 5 h post-decentralization period of data from each decentralized preparation within the first 120 h. n.s., not significant; ***p < 0.001.
- Figure 7
Daily saline exchanges transiently activate the pyloric rhythm. A, Example pyloric cycle frequency plot for an 8 h stretch surrounding a saline exchange that transiently activates the pyloric rhythm (done at arrow). B, Raw data from the same preparation reveals an increased frequency lasting several hours. C, Population data showing the average effect of exchanging saline in 10 long-term decentralized preparations. Left-most box shows time-matched data from continuously superfused preparations. ***p < 0.001.





Tables
Data n R p value Intact, Hz vs PD off 123 0.4113 1.19E-05 Intact, Hz vs LP on 123 0.0555 0.5759 Intact, Hz vs LP off 123 0.3422 3.77E-04 Decentralized, Hz vs PD off 115 0.8229 <1E-06 Decentralized, Hz vs LP on 115 0.8298 <1E-06 Decentralized, Hz vs LP off 115 0.8295 <1E-06 Intact, Hz vs 30 min decentralized Hz 115 0.4077 6.1049E-06 30 min decentralized Hz vs 2 d decentralized Hz 19 0.5234 0.0258 2 d decentralized Hz vs 3 d decentralized Hz 19 0.6046 7.90E-03 3 d decentralized Hz vs 4 d decentralized Hz 19 0.4249 0.0788 5 h recovery time vs time to first bout 19 0.151 0.537 Data n t p value Paired t tests Intact Pyloric Hz vs 30 min decentralized Hz 115 40.768 <1E-06 Hz, pre- vs post-saline exchanges (avg) 10 −5.2897 5.01E-04 Unpaired t tests Time-matched vs saline-exchange pre 13 matched; 10 exchanged −2.4662 0.028 Time-matched vs saline-exchange post 13 matched; 10 exchanged −5.2897 5.01E-04 Data n Between-subjects F Between-subjects p Within-subjects F Within-subjects p Interaction F Interaction p Pyloric Hz 19; 9 108.12 <1E-06 1.632 0.1578 1.775 0.1234 Cycle variation 19; 9 5.134 0.0347 0.426 0.82975 0.365 0.87119 PD off 19; 9 24.944 6.07E-05 0.254 0.93719 0.703 0.6226 LP on 19; 9 5.382 0.0489 0.687 0.63616 0.702 0.625 LP off 19; 9 9.234 0.016 2.175 0.076 0.55 0.7376 Bouts 19; 9 9.417 6.06E-03 7.337 6.8E-06 6.393 3.42E-06 Data n χ2 p value Fastest 5 h decentralized vs intact Pyloric Hz 19 5-h decentralized stretches; 123 intact 17.56 2.78E-05 Fastest 5 h decentralized vs intact cycle variation 19 5-h decentralized stretches; 123 intact 21.935 2.82E-06 Fastest 5 h decentralized vs intact PD off 19 5-h decentralized stretches; 123 intact 17.6187 2.70E-05 Fastest 5 h decentralized vs intact LP on 19 5-h decentralized stretches; 123 intact 8.1044 4.40E-03 Fastest 5 h decentralized vs intact LP off 19 5-h decentralized stretches; 123 intact 11.6703 6.35E-04 Fastest 5 h decentralized vs 30 min decentralized Hz 19 5-h decentralized stretches; 115 30-min decentralized 1.5332 0.2156 Fastest 5 h decentralized vs 30 min decentralized cycle variation 19 5-h decentralized stretches; 115 30-min decentralized 5.1989 0.0226 Fastest 5 h decentralized vs 30 min decentralized PD off 19 5-h decentralized stretches; 115 30-min decentralized 1.5585 0.2119 Fastest 5 h decentralized vs 30 min decentralized LP on 19 5-h decentralized stretches; 115 30-min decentralized 1.348 0.2456 Fastest 5 h decentralized vs 30 min decentralized LP off 19 5-h decentralized stretches; 115 30-min decentralized 1.348 0.2456
In this issue

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}



