

Abstract
Central vestibular neurons receive substantial inputs from the contralateral labyrinth through inhibitory and excitatory brainstem commissural pathways. The functional organization of these pathways was studied by a multi-methodological approach in isolated frog whole brains. Retrogradely labeled vestibular commissural neurons were primarily located in the superior vestibular nucleus in rhombomeres 2/3 and the medial and descending vestibular nucleus in rhombomeres 5–7. Restricted projections to contralateral vestibular areas, without collaterals to other classical vestibular targets, indicate that vestibular commissural neurons form a feedforward push–pull circuitry. Electrical stimulation of the contralateral coplanar semicircular canal nerve evoked in canal-related second-order vestibular neurons (2°VN) commissural IPSPs (∼70%) and EPSPs (∼30%) with mainly (∼70%) disynaptic onset latencies. The dynamics of commissural responses to electrical pulse trains suggests mediation predominantly by tonic vestibular neurons that activate in all tonic 2°VN large-amplitude IPSPs with a reversal potential of −74 mV. In contrast, phasic 2°VN exhibited either nonreversible, small-amplitude IPSPs (∼40%) of likely dendritic origin or large-amplitude commissural EPSPs (∼60%). IPSPs with disynaptic onset latencies were exclusively GABAergic (mainly GABAA receptor-mediated) but not glycinergic, compatible with the presence of GABA-immunopositive (∼20%) and the absence of glycine-immunopositive vestibular commissural neurons. In contrast, IPSPs with longer, oligosynaptic onset latencies were GABAergic and glycinergic, indicating that both pharmacological types of local inhibitory neurons were activated by excitatory commissural fibers. Conservation of major morpho-physiological and pharmacological features of the vestibular commissural pathway suggests that this phylogenetically old circuitry plays an essential role for the processing of bilateral angular head acceleration signals in vertebrates.
Introduction
A major functional principle in the vestibular as in the auditory system for improving sensory signal processing is an interconnection of central neurons by brainstem-crossing inhibitory pathways (Grothe, 2003; Straka and Dieringer, 2004). Second-order vestibular neurons (2°VN) are interconnected by canal-specific commissural pathways that maintain a balanced activity at rest (Graham and Dutia, 2001), increase the sensitivity for angular head acceleration-related signals (Shimazu and Precht, 1966; Shimazu, 1972; Markham et al., 1977), mediate inhibitory signals for the angular vestibulo-ocular reflex velocity storage (Wearne et al., 1997; Holstein et al., 1999), and recalibrate the asymmetric resting activity of 2°VN after a unilateral vestibular loss (Curthoys, 2000) through plastic changes of the GABAergic transmission (Yamanaka et al., 2000; Bergquist et al., 2008). The number of organizational similarities of this pathway across different vertebrate species suggests a rather conserved functional role during phylogeny (Straka and Dieringer, 2004; Highstein and Holstein, 2006). Common organizational principles include a plane specificity of inhibitory commissural connections from contralateral coplanar semicircular canals, a larger proportion of vertical than horizontal canal neurons that receive a commissural inhibition, as well as mediation by midline-crossing inhibitory type I neurons as well as by excitatory commissural neurons and inhibitory type II neurons on the other side (Shimazu and Precht, 1966; Kasahara and Uchino, 1974; Holler and Straka, 2001).
A GABAergic and glycinergic commissural inhibition (Precht et al., 1973a) is supported by the presence of both types of inhibitory commissural neurons (Bagnall et al., 2007), whereas an exclusive GABAergic vestibular commissural inhibition has been reported in brainstem slices (Furuya et al., 1991; Furuya and Koizumi, 1998). This apparent discrepancy in transmitter use as well as the additional presence of a canal commissural excitation in some 2°VN (Shimazu and Precht, 1966; Holler and Straka, 2001) might be related to a more specific functional organization of inhibitory/excitatory commissural signals that depends on the dynamic profile, (i.e., type A/B vestibular neurons in mammals) of both the commissural projections and the contralateral target neurons (Straka et al., 2005). The different organization of GABAergic and glycinergic inhibition in mouse type A and B vestibular neurons (Camp et al., 2006) as well as the presence of commissural excitatory responses in some 2°VN (Broussard and Lisberger, 1992; Holler and Straka, 2001) suggest rather specific functional implications of dynamically different inhibitory and excitatory vestibular neurons in commissural signaling and a major role of these connections in the adaptive plasticity of vestibulo-motor responses.
The goal of this study was thus to obtain a comprehensive knowledge of semicircular canal commissural functional organization, including cellular, developmental, morpho-physiological, and pharmacological principles in a single species. These aspects were studied in isolated frog whole brains with intact semicircular canal nerve branches to selectively activate commissural inputs in functionally characterized 2°VN. Selective lesions of fiber tracts and combined retrograde tracing and immunohistochemistry revealed the hindbrain segmental organization of commissural neurons and allowed distinguishing between an organization as a feedforward push–pull or feedback closed-loop mode.
Parts of this work have been published previously in abstract form (Straka and Gilland, 2002).
Materials and Methods
Electrophysiology and pharmacology.
In vitro electrophysiological experiments were performed on the isolated brains of 26 grass frogs (Rana esculenta) and complied with the Principles of Animal Care (Publication 86-23, revised 1985 by the National Institute of Health). As described in previous studies (Straka and Dieringer, 1993), the animals were deeply anesthetized with 0.1% 3-aminobenzoic acid ethyl ester (MS-222) and perfused transcardially with iced Ringer's solution (in mm: 75 NaCl, 25 NaHCO3, 2 CaCl2, 2 KCl, 0.5 MgCl2, and 11 glucose, pH 7.4). Thereafter, the skull and the bony labyrinths were opened by a ventral approach. After dissecting the three semicircular canals on each side, the brain was removed with all labyrinthine end organs attached to the eighth cranial nerve (N.VIII). Subsequently, the brain was submerged in iced Ringer's solution, and the dura, the labyrinthine end organs and the choroid plexus covering the IVth ventricle were removed. In all experiments, the forebrain was detached. Brains were used up to 4 d after their isolation and were stored overnight at 6°C in oxygenated Ringer's solution (carbogen: 95% O2, 5% CO2) with a pH of 7.5 ± 0.1. For the experiments, the brainstem was glued with cyanoacrylate to a plastic mesh, which was fixed with insect pins to the Sylgard floor of a chamber (volume of 2.4 ml). For the experiments in which a longitudinal incision of the internal and/or external arcuate fiber tracts was made, the brains were directly fixed with insect pins to the Sylgard floor to allow flexible manipulation of the brains before and after the lesion. The recording chamber was continuously perfused with oxygenated Ringer's solution at a rate of 1.3–2.1 ml/min. The temperature was electronically controlled and maintained at 14 ± 0.1°C.
For electrical activation of individual semicircular canal nerve branches, single constant current pulses (duration, 0.2 ms; threshold, 1.5–3.1 μA) were produced with a stimulus isolation unit (WPI A 360) and applied across individually adjusted suction electrodes (diameter, 120–150 μm). These suction electrodes allowed electrical isolation of the individual nerve branches inside the customized glass capillaries and prevented current spread to auditory nerve fibers, the facial nerve, or the other semicircular canal nerves as demonstrated previously in the isolated frog whole brain (Straka et al., 1997). In the few experiments in which the N.VIII was stimulated, bipolar concentric electrodes (tip diameter, 25 μm) were used. Glass microelectrodes used for extracellular and intracellular recordings were made with a horizontal puller (P-87 Brown/Flaming). Electrodes for extracellular field potential recordings were beveled (30°, 20 μm tip diameter) and filled with a 2 m solution of sodium chloride (∼1 MΩ final resistance). Electrodes for intracellular recordings were filled with a mixture of 2 m potassium acetate and 3 m potassium chloride (10:1), which gave a final resistance of ∼90–110 MΩ. Neuronal recordings were made in bridge mode (SEC-05L; NPI Electronic).
At the beginning of each experiment, presynaptic and postsynaptic field potentials (Precht et al., 1974) were evoked by separate stimulation of individual ipsilateral labyrinthine nerve branches or the entire N.VIII and recorded at a standard reference site in the vestibular nuclei (Straka et al., 1997). The amplitude of the field potentials served to optimize the position of the stimulus electrodes and to determine the stimulus threshold (T) for each branch. Stimulus intensities were indicated as multiples of the threshold intensity (×T) for the postsynaptic field potentials. Intracellular recordings were obtained from neurons in the vestibular nuclei after identification as 2°VN by monosynaptic EPSPs following separate electrical stimulation of the three ipsilateral semicircular canal nerve branches and/or of the N.VIII. A total of 354 vestibular neurons in different sets of experiments were recorded in all vestibular subnuclei [superior (SVN), medial (MVN), lateral (LVN), and descending (DVN) vestibular nucleus], except the most medial parts of the medial vestibular nucleus. After identification as 2°VN, the presence of commissural semicircular canal inputs was tested by stimulation of the contralateral N.VIII or by separate stimulation of the three contralateral semicircular canal nerve branches. Based on different discharge patterns during long positive current steps, frog 2°VN were distinguished as phasic or tonic vestibular neurons (Straka et al., 2004; Beraneck et al., 2007). As reported previously, most of the vestibular neurons in the isolated frog brain had no spontaneous discharge at resting membrane potential (Straka and Dieringer, 2004; Straka et al., 2005). Only neurons with a membrane potential more than −55 mV were included in this study.
The implication of GABAergic and/or glycinergic synapses in the commissural inhibition of 2°VN was studied by bath application of the glycine-receptor blocker strychnine hydrochloride (1 μm; Sigma), the GABAA-receptor blocker bicuculline methochloride (1–5 μm; Sigma), and the GABAB-receptor blocker CGP 35348 (p-3-aminopropyl-p-diethoxymethyl phosphoric acid) (100 μm; Tocris Bioscience). Antagonist-related changes of the evoked synaptic responses occurred after ∼5 min and reached steady state after 10–15 min (Biesdorf et al., 2008). After 30–40 min, the washout of the different antagonists was usually complete.
Single sweeps of the responses were digitized at 20 kHz (CED 1401; Cambridge Electronic Design), stored on computer, and analyzed offline (SIGNAL; Cambridge Electronic Design). Synaptic potentials were analyzed from averages of 10–30 single sweeps after electronic subtraction of the extracellular field potential recorded in the vicinity. Statistical differences in parameters were calculated according to the Mann–Whitney U test (unpaired parameters) and the Wilcoxon's signed rank test (paired parameters; Prism; Graphpad Software). All averaged results were expressed as mean ± SE. Graphical presentations were made with the aid of commercially available computer software [Origin (Microcal Software); Corel Draw (Corel Corporation)].
Lesions of commissural fiber tracts were made by bilateral longitudinal incisions from the level of the cerebellum to the obex with a small piece of a razor blade. These incisions interrupted on both sides either the internal arcuate fiber tract (IAF) (n = 3) or the external arcuate fiber tract (EAF) (n = 3), or the IAF on one side and the EAF on the other side (n = 3). Vestibular neurons were recorded on both sides of the brainstem before (n = 39) and after (n = 107) the lesion. The location and extension of the cuts was verified histologically after each experiment. For this purpose, the brains were fixed for 4 h in a phosphate buffer (PB) solution, pH 7.4, containing 2% paraformaldehyde and 1% glutaraldehyde and were subsequently transferred into 15% sucrose in PB for 2–4 h and into 30% sucrose in PB overnight. Brains were cut coronally in 80 μm sections, mounted on gelatin-coated slides, dried overnight, counterstained with cresyl violet, and coverslipped.
Modeling.
Simultaneous recordings of presynaptic commissural fibers and postsynaptic vestibular target neurons are almost impossible. To determine the dynamic profile of the midline-crossing 2°VN (i.e., tonic–phasic) that evoked commissural IPSPs in tonic 2°VN, a modeling approach was used. This method took into account the synaptic delay and time constants of commissural responses to single pulses, recorded in this study, as well as the known differential discharge dynamics of tonic and phasic 2°VN during sinusoidally modulated pulse trains (Pfanzelt et al., 2008) to calculate compound commissural inhibitory model responses. By comparison of the time courses of calculated model and recorded responses in the current study, it was possible to determine the dynamic profile of the presynaptic midline-crossing 2°VN that evoked the commissural inhibitory response in tonic 2°VN. Modeling of the commissural synaptic signal processing in tonic 2°VN was performed by extending the previously described intrinsic tonic 2°VN model (Pfanzelt et al., 2008) with dual-exponential conductance-based synapses at the soma. Synapses were realized using the following equation: II = gIF [exp(−t/τ2)−exp(−t/τ1)](V − EI), with τ1 as rise and τ2 as fall time constants (Johnston and Wu, 1994). The peak conductance for a single synaptic event was defined by gI. EI indicates the reversal potential and V the somatic membrane potential. The scaling factor F is defined such that the peak of the dual exponential is 1. Real-time compartmental models were simulated in NEURON (Carnevale and Hines, 2006), and the synapses were implemented by the function “Exp2Syn.” Synaptic parameters were estimated by fitting the model to the first seven IPSPs in a tonic 2°VN after activation of the contralateral coplanar canal nerve with a stimulus train that consisted of a sinusoidally modulated sequence of single electrical pulses (Pfanzelt et al., 2008). Using the parameters of a pulse train with a half-cycle length of 1 s and a peak frequency of 70 Hz, the fitting was performed with the Multiple Run Fitter Tool in NEURON. Each single stimulus triggered a synaptic IPSP with an average disynaptic onset latency of 8.3 ms (Holler and Straka, 2001). The fitting operation yielded the following parameters: τ1 = 6.1 ms, τ2 = 26.6 ms, gI = 50.4 nS, EI = −75.6 mV. The entire stimulus train was simulated using three hypothesized neuronal configurations for mediation of the commissural inhibition: (1) tonic commissural 2°VN, in which each single stimulus of the pulse train triggers a synaptic event; (2) phasic commissural 2°VN, in which only the first seven stimuli of the pulse train trigger a synaptic event based on the discharge dynamics of these neurons (Pfanzelt et al., 2008); (3) two synaptic processes via tonic and phasic commissural 2°VN, each contributing 50% to the commissural IPSPs with half of the conductance of gI.
Anatomy and immunohistochemistry.
The spatial distribution of vestibular commissural neurons was evaluated by retrograde tracing from three different contralateral vestibular target areas that were stereotactically defined with respect to the caudal end of N.VIII and coincided with different vestibular subnuclei (Reichenberger et al., 1997; Straka et al., 2000, 2003, 2006). Crystals of Alexa Fluor 488 or 546 dextran (Invitrogen) were inserted from the dorsal surface into the vestibular nuclei in isolated frog brains in vitro. Tracer applications to the rostral part, i.e., between −0.5 mm rostral and 0 mm to N.VIII (n = 6), primarily covered the SVN, whereas those to the intermediate part, i.e., between 0 and 0.5 mm caudal to N.VIII (n = 6), coincided with the LVN and DVN. Applications to the caudal part, i.e., between 0.5 and 1.0 mm caudal to N.VIII (n = 5), exclusively covered the DVN. Different parts of the MVN were included at all levels, respectively. In an additional set of experiments (n = 3), crystals of Alexa Fluor 488 dextran were inserted in the rostral part and of Alexa Fluor 546 dextran in the caudal part of the vestibular nuclei on the same side. With respect to the hindbrain segmental organization, the application sites coincided with rhombomeres (r) r2/r3, r4/r5, and r6/r7, respectively. This correlation is possible because a stereotactic reference frame of the rhombomeric organization in adult frogs was established on the basis of preserved hindbrain motoneuronal segmental positions relative to the visible rhombomeres in larval frogs (Straka et al., 2006). The cerebellum was split in the midline in all cases to exclude potential labeling of vestibular commissural neurons via cerebellar pathways. Because the auditory nucleus, located dorsal to the vestibular nuclei in r4–r6, was inevitably included during tracer applications at this level, the retrograde labeling of auditory commissural neurons served as a control after tracer application into the intermediate part of the vestibular nuclei (Grofová and Corvaja, 1972; Feng and Lin, 1996).
After 48 h of survival at 11°C in oxygenated Ringer's solution (Birinyi et al., 2001; Straka et al., 2006), the isolated brains were fixed in 4% paraformaldehyde in 0.1 m PB for 24 h and stored overnight in 15 and 30% sucrose in 0.1 m PB. All brains were cut coronally in 50 μm sections on a cryostat. Sections were mounted on slides, coverslipped with Vectashield (Vector Laboratories by Biovalley) and analyzed by laser scanning confocal microscopy (Carl Zeiss LSM 510) at a wavelength of 488 or 543 nm. Stacks of 10–20 confocal images were generated with a 10×/0.3 numerical aperture and 20×/0.5 numerical aperture objective and a plan Apochromat 40×/1.3 oil-immersion objective. Images for Figures 1 and 4 were produced by horizontal projection of the entire stack.
The number of retrogradely labeled commissural vestibular neurons was counted on each of the 50 μm section between 0.75 mm rostral and 1.5 mm caudal to the entry of N.VIII in the brainstem and plotted with respect to the rostrocaudal position of the section. The indicated area essentially covers the entire rostrocaudal extent of the vestibular nuclei (Reichenberger et al., 1997). The diameters of the labeled neurons were determined from confocal scanning images using NIH Image J software and were expressed as mean ± SE. A mapping of the commissural neurons relative to the four classical vestibular nuclei was obtained by superimposing the cellular location on the section on schematic outlines of the frog vestibular nuclei after adjustment of the calibration (Straka et al., 2000, 2003). Representative views of all four vestibular nuclei were obtained after reconstruction of frontal sections at five representative rostrocaudal levels (−0.5 mm rostral, 0 mm, 0.4 mm, 0.7 mm, and 1.0 mm caudal to the entry of the N.VIII) from published data (Matesz, 1979; Kuruvilla et al., 1985). The position of vestibular commissural neurons within the hindbrain segmental scaffold was determined by using previously published stereotactic coordinates of the individual segments in adult frog (Straka et al., 2006). This latter procedure allowed linking the spatial position of vestibular commissural neurons in the adult frog hindbrain with the structural elements for attaining segmentally specific identities of the neuronal populations during ontogeny.
Combined retrograde tracing and GABA/glycine immunohistochemistry was used to determine the contributions of GABAergic and/or glycinergic neurons to the vestibular commissural pathway. Vestibular commissural neurons were labeled after insertion of crystals of Alexa Fluor 488 dextran into rostral and intermediate parts of the vestibular nuclei in isolated frog whole-brain preparations in vitro (n = 10) as described above. After 48 h of survival at 11°C in continuously oxygenated Ringer's solution, brains were fixed overnight in 3% paraformaldehyde and 1% glutaraldehyde in 0.1 m PB and stored for 24 h in 15 and 30% sucrose in 0.1 m PB. Coronal sections (20 μm) were cut on a cryostat, mounted on slides, and kept at −80°C until use. After rinsing in 0.1 m PBS, sections were treated with 0.1 m sodium borohydride for 10 min, rinsed again, and preincubated in PBS supplemented with 10% normal goat serum and 0.1% Triton X-100 for 30 min. Primary antibodies were diluted in this solution and applied overnight at 4°C. The antibodies consisted of a mouse anti-GABA monoclonal antibody (GABA93 monoclonal; generated by the laboratory of Dr. G. R. Holstein, Departments of Neurology, Neuroscience, and Anatomy/Functional Morphology, Mt. Sinai School of Medicine, New York, NY) (Holstein et al., 2004) at 1:500 dilution and a rabbit anti-glycine antibody (Millipore Bioscience Research Reagents) at 1:2000 dilution. Antibody binding was detected with goat anti-rabbit or goat anti-mouse antibodies conjugated to cyanine 3 (Jackson ImmunoResearch by Interchim). Subsequently, sections were washed, air dried, and coverslipped with Vectashield. Sections were analyzed by laser scanning confocal microscopy (see above).
Results
Segmental organization of vestibular commissural neurons
The location of retrogradely labeled vestibular commissural neurons was revealed after insertion of crystals of Alexa Fluor 488 (green) and/or 546 (red) dextran into the vestibular nuclear areas (Fig. 1A,G) at a rostral (SVN/MVN; n = 6), an intermediate (LVN/DVN/MVN; n = 6), and a caudal (DVN; n = 5; see Material and Methods) level. The application sites usually extended for ∼0.25 mm in all directions and included at the intermediate level the auditory dorsal nucleus (DN), located dorsal to the vestibular nuclei at the level of the LVN (Fig. 1A,G). In contrast, tracer application never extended ventrally into the reticular formation located below an oblique mediolateral line, marked by the sulcus limitans on the ventricular side (Fig. 1A, white arrow). Mapping the retrogradely labeled vestibular commissural neurons onto a rostrocaudal coordinate system with reference to N.VIII facilitated correlation of the neuronal positions with the cytoarchitectonically defined vestibular subnuclei (Fig. 2) described in frogs (Matesz, 1979; Kuruvilla et al., 1985), as well as with the intrinsic, ontogenetically derived hindbrain segmental framework (rhombomeres r2–r7 in Figs. 2, 3) (see Material and Methods). This latter correlation allowed direct comparison of the spatial organization of vestibular commissural neurons in adult frogs with the developmental origin of the respective segmentally organized populations in young larvae (Straka et al., 2001) as well as with the embryonic patterns of other vertebrates (Cambronero and Puelles, 2000; Glover, 2000).

Photomicrographs of coronal sections showing confocal reconstructions of retrogradely labeled vestibular commissural, vestibulo-ocular, and vestibulo-spinal neurons. A–C, Application site of Alexa Fluor 488 dextran (green area) in the rostral region of the VN (A), crossing commissural axons in the midline (ml; inset in A) and retrogradely labeled cell bodies of commissural neurons in the contralateral VN (B, C); higher magnification (C) of labeled neurons in the outlined area in B. D, E, Separate populations of vestibular commissural neurons (green, red) after application of Alexa Fluor 488 dextran (green) into the rostral and of Alexa Fluor 546 dextran (red) into the caudal region of the vestibular nuclei. Only very few double-labeled cells (yellow) were labeled in the caudal region of the contralateral VN; higher magnification (E) of labeled neurons in the outlined area in D and in a section 100 μm more caudal (F). G–K, Target sites after application of Alexa Fluor 488 dextran (green) into the left VN (G) and of Alexa Fluor 546 dextran (red) into the right oculomotor nucleus (N.Oc; H). Separate populations of vestibular commissural (green) and vestibulo-ocular neurons (red) were retrogradely labeled in rostral regions of the VN (I–K); higher magnification (J) of labeled neurons in the outlined area in I and in a section 50 μm more caudal (K). L, M, Separate populations of retrogradely labeled vestibular commissural (green) and of vestibulo-spinal neurons (red) in caudal regions of the VN after application of Alexa Fluor 488 dextran into the VN and of Alexa Fluor 546 dextran bilaterally into the upper spinal cord; higher magnification (M) of labeled neurons in the outlined area in L. N, Commissural neurons in the DN after tracer application into the contralateral vestibular/auditory nuclei (see A) at the level of N.VIII; the inset shows DN neurons in N at higher magnification. White arrows in A, B, D, and L indicate the sulcus limitans. Scale bars: A, B, D, G, H, I, L, N, inset in A, 300 μm; C, E, J, M, inset in N, 100 μm; F, K, 50 μm. lat, Lateral; med, medial.

Schematic drawings of coronal sections through the dorsal hindbrain summarizing the location of vestibular/auditory commissural neurons. A–C, Location of retrogradely labeled neurons with respect to vestibular and auditory nuclei at five rostrocaudal levels after tracer applications into rostral (n = 6; A), intermediate (n = 6; B), and caudal (n = 5; C) areas of the contralateral vestibular nuclei. The different symbols in each row represent labeled cells from different experiments, respectively. The rostrocaudal level of the sections with respect to the caudal end of the entry of N.VIII in the brainstem (0 mm) is indicated in A; the rostrocaudal position of the sections with respect to the hindbrain segmental scaffold (r2–r7) was obtained from Straka et al. (2006) and is indicated in B. The outline of the vestibular subnuclei was adopted from Kuruvilla et al. (1985) and Matesz (1979). The arrow at each level in C indicates the sulcus limitans (s.l.).

Distribution of vestibular commissural, GABAergic, and glycinergic vestibular neurons along the rostrocaudal extent of the vestibular nuclei. A–C, Number of labeled vestibular commissural neurons per section after application of Alexa Fluor 546 dextran (red bars) to rostral (A; n = 6), intermediate (B; n = 6), or caudal (C; n = 5) areas of the contralateral vestibular nuclei; different symbols in A–C represent individual experiments and the red line the average of these data, respectively. D, Number of double-labeled vestibular commissural neurons per section after application of Alexa Fluor 546 dextran to rostral (red bar) and Alexa Fluor 488 dextran (green bar) to caudal areas of the contralateral vestibular nuclei (n = 3); different symbols represent individual experiments and the orange line the average of these data. E, Average number of retrogradely labeled vestibular commissural neurons per section after rostral, intermediate, and caudal vestibular tracer applications (data from A–C). F–H, Average number of GABAergic (F) and glycinergic (G) vestibular neurons per section (data adopted from Reichenberger et al., 1997) and comparison with the normalized rostrocaudal distribution of retrogradely labeled vestibular commissural neurons (Com; H); different symbols in F and G represent individual experiments and the yellow (F) and blue line (G) the average of these data, respectively. In H, the number of cells per section was normalized to the maximal number of each type. Vertical stripes in the background of each plot indicate the hindbrain segmental scaffold (r2–r7) obtained from Straka et al. (2006). Note that GABAergic and vestibular commissural neurons have a similar rostrocaudal distribution between r2 and r6.
Application of Alexa Fluor 488 or 546 dextran into the vestibular nuclei at any rostrocaudal level labeled neurons on the contralateral side in particular vestibular subnuclei, the adjacent reticular formation, and the auditory DN (Fig. 1B–F,I–K). The axons of these retrogradely labeled neurons crossed the midline in several broad bundles (Fig. 1A, inset) at the level of the parent cell body. Commissural neurons in the vestibular nuclei comprised a relatively homogeneous population of mostly small, seldom medium-sized neurons (mean ± SE, 15.3 ± 3.3 μm; n = 310) with an oval or round cell body (Fig. 1C,E,F,J,K,M). Independent of the location of the tracer application site, labeled neurons clustered in two subgroups with respect to the rostrocaudal extent of the vestibular nuclei that coincided with hindbrain segments r2/r3 (rostral subgroup) and r5–r7 (caudal subgroup) (Figs. 2A–C, 3A–C,E). The location of the r2/r3 cluster of vestibular commissural neurons primarily correlated with the SVN/MVN (Figs. 2A–C, 3A–C), with the majority of commissural neurons in the r2 and fewer neurons in the r3 portion of the SVN (Fig. 3E). Within the SVN, labeled neurons tended to be located mainly in medial and dorsal parts of the nucleus (Figs. 1I,J, green neurons; 2A–C). The location of the more dispersed r5–r7 cluster overlapped with the DVN/MVN with a predominance of commissural neurons in r6/r7 throughout the dorsoventral extent of the DVN (Figs. 2A–C, 3A–C).
The number of labeled neurons in the rostral subgroup was considerably larger after tracer applications to the contralateral vestibular nuclear area that included the SVN and rostral parts of the MVN (∼5–15 cells per section) (Figs. 2A,B, 3A,B) compared with more caudal applications that primarily covered the DVN and caudal parts of the MVN (approximately three to five cells per section) (Figs. 2C, 3C). In contrast, the caudal subgroup of vestibular commissural neurons always contained similar numbers of neurons (approximately four cells per section), independent of the rostrocaudal location of the application site (Fig. 3A–C). The two distinct clusters of vestibular commissural neurons were separated by a zone with very few or no labeled cells that corresponded to the LVN located in r4 (Figs. 2A–C, 3A–C,E). The virtual absence of labeled neurons indicates that this vestibular subnucleus does not give rise to commissural projections (Fig. 2A–C). However, many vestibular commissural neurons were labeled after tracer application that included this nucleus (Fig. 2B; injections in the intermediate part in Fig. 3B,E), suggesting that LVN neurons in turn receive substantial commissural inputs from the contralateral SVN/MVN and DVN/MVN in r2/r3 and r5–r7, respectively. In addition to the vestibular commissural neurons, a considerable number of retrogradely labeled small- to medium-sized oval neurons (10–25 μm) were located just ventral to the DVN in the adjacent reticular formation (Fig. 2A–C) in r6/r7. Because these neurons form a population that is primarily contiguous with the DVN at its caudal aspect, it is possible that this area is also part of the vestibular nucleus that receives labyrinthine afferent inputs, although it is classically not included in the description of the vestibular nuclear region that was defined in frog by cell cytoarchitectonics (Gregory, 1972; Matesz, 1979).
A significant number of labeled commissural neurons were observed in the auditory DN in r4–r6 (Fig. 1N) mainly after tracer application to intermediate vestibular areas, which included the contralateral auditory nucleus (Fig. 2A–C). This result complies with earlier descriptions of a prominent commissural pathway that interconnects the bilateral DN (Feng and Lin, 1996). Labeled DN neurons were usually small (10–15 μm) with a round or oval cell body (Fig. 1N, inset) and mostly located in the rostral part of the DN (Fig. 2B, 0 mm), according to the histological outline of the nuclear boundaries (Kuruvilla et al., 1985; Feng and Lin, 1996).
Combined application of Alexa Fluor 546 dextran to rostral (i.e., the SVN/MVN) and of Alexa Fluor 488 dextran to caudal (i.e., the DVN) vestibular areas labeled separate, but spatially overlapping, populations of vestibular commissural neurons (Fig. 1D–F, red and green neurons). Only very few neurons were double labeled (Fig. 1D–F, yellow neurons), indicating that the vast majority of vestibular commissural neurons have rather circumscribed, spatially limited terminal regions in the contralateral vestibular nuclei. Almost all double-labeled neurons were encountered in the caudal part of the vestibular nuclear area, i.e., the DVN mainly in r6 and essentially none in the SVN in r2/r3 (Fig. 3D). The absence of larger numbers of double-labeled vestibular commissural neurons further suggests that the axons of these neurons, after crossing the brainstem at the level of the parent cell body, enter the contralateral target area only at the rostrocaudal level of the respective termination site. Axonal trajectories through larger portions of the contralateral vestibular nuclei are thus unlikely, because many more axons and parent cell bodies would have been double labeled after combined tracer application to different vestibular areas.
Vestibular commissural connections as feedforward push–pull or feedback closed-loop circuitry
Vestibular commissural connections might be either organized as feedforward or as feedback circuitry with different implications for the bilateral signal processing, respectively (Galiana and Outerbridge, 1984; Galiana et al., 1984). The two possibilities differ from each other by the presence of additional projections of individual vestibular commissural neurons to targets other than the contralateral vestibular nuclei. Because vestibular commissural neurons in the SVN, DVN, and MVN in frog overlap in part with vestibulo-ocular (Straka and Dieringer, 1991), vestibulo-spinal (Ten Donkelaar, 1982; Matesz et al., 2002), and cerebellar-projecting vestibular neurons (Grover and Grüsser-Cornehls, 1984), the possibility exists that some vestibular commissural neurons have axon collaterals that project to one or the other additional targets. This was tested by combined application of Alexa Fluor 488 and 546 dextran to the vestibular nuclear area on one side and unilaterally or bilaterally to the oculomotor nucleus (Fig. 1G,H), the upper spinal cord, or the cerebellum. Combined tracer application to the vestibular nuclei (Fig. 1H) and to the ipsilateral or contralateral oculomotor nucleus (Fig. 1G) labeled distinct but spatially more or less overlapping populations of vestibular commissural (Fig. 1I–K, green neurons) and of vestibulo-ocular neurons (Fig. 1I–K, red neurons), mainly in the SVN. Cell bodies of vestibulo-ocular neurons (mean ± SE, 16.4 ± 3.5 μm; n = 108) were on average somewhat larger than those of vestibular commissural neurons (p ≤ 0.01, Mann–Whitney U test). Importantly, no double-labeled neurons, indicating a dual projection of individual neurons to both target areas, were encountered.
A similar result, i.e., the absence of double-labeled neurons, was obtained after combined tracer application to the vestibular nucleus and to the upper spinal cord (Fig. 1L,M) or the cerebellum (data not shown). Thus, vestibular commissural and vestibulo-spinal or vestibulo-cerebellar neurons also form separate functional subgroups of vestibular neurons (Fig. 1L,M). Because vestibular commissural neurons were located mainly in the SVN and DVN and vestibulo-spinal neurons in the LVN (Matesz et al., 2002) and DVN, a limited spatial overlap of the two distinct subpopulations was encountered in the latter vestibular nucleus (Fig. 1L,M). The separation into distinct populations of vestibular commissural and vestibulo-spinal neurons was also mirrored by the clear difference in cell size. Cell bodies of vestibulo-spinal neurons (Fig. 1L,M, red neurons) were significantly larger (mean ± SE, 23.7 ± 5.3 μm; n = 104; p ≤ 0.0001, Mann–Whitney U test) than those of vestibular commissural neurons (Fig. 1L,M, green neurons). In contrast, cell bodies of cerebellar-projecting vestibular neurons were rather small (mean ± SE, 15.1 ± 2.8 μm; n = 59) and similar in size to those of vestibular commissural neurons. The latter two neuronal subtypes spatially overlapped in the DVN and to a smaller extent in the SVN and MVN, consistent with previous reports on the location of cerebellar-projecting vestibular neurons (Grover and Grüsser-Cornehls, 1984). The general absence of double-labeled neurons after combined tracer application to the vestibular nuclei and other classical vestibular efferent projection sites indicates that vestibular commissural neurons form a particular functional subgroup of vestibular neurons with a single target, the contralateral vestibular nuclei.
GABA and glycine immunohistochemistry of vestibular commissural neurons
The ultrastructure of a particular population of GABAergic velocity storage-related commissural neurons in the rostral medial vestibular nucleus along with the synaptic terminations on different postsynaptic cellular sites has been shown previously (Holstein et al., 1999). Although the subcellular structure of these neurons including the morphology of axo-dendritic and axo-axonic contacts were described in detail, the anatomical distribution of the entire population of GABAergic vestibular commissural neurons within the different vestibular nuclei and the hindbrain segmental scaffold as well as the role of potential inhibitory glycinergic commissural vestibular neurons remains unknown. Therefore, the organization and spatial arrangement of the entire population of GABAergic and glycinergic vestibular commissural neurons was determined to correlate the respective hindbrain positions with the different vestibular nuclei and the underlying rhombomeric framework. Retrograde tracing of commissural neurons after injection of Alexa Fluor 488 dextran into the contralateral VN combined with GABA (n = 5) and glycine (n = 5) immunohistochemistry revealed the pattern and profile of inhibitory vestibular commissural projections. Among the population of retrogradely labeled commissural vestibular neurons (Fig. 4A,B, green neurons), a subgroup of ∼20% was also GABA immunopositive (Fig. 4B, * yellow neurons) as indicated by the red immunostaining of these cells (Fig. 4B, inset). These double-labeled cells were not clustered but intermingled with the larger group of GABA-immunonegative commissural neurons and almost entirely located in the SVN/MVN in r2/r3, in which vestibular commissural neurons overlap to a large extent with GABAergic vestibular neurons (Fig. 3H) (Reichenberger et al., 1997). Because of limited antibody penetration in the tissue that impairs complete staining of all GABAergic neurons, the percentage of GABA-immunopositive vestibular commissural neurons might however be an underestimation. Nonetheless, it clearly shows that a noticeable population of commissural vestibular neurons that project across the midline is putatively GABAergic.

Photomicrographs of coronal sections showing confocal reconstructions of retrogradely labeled GABAergic or glycinergic commissural neurons. A–C, Combined GABA immunohistochemistry (red) and retrograde tracing with Alexa Fluor 488 dextran (green) revealed double-labeled GABA-immunopositive vestibular commissural neurons (yellow) in rostral regions of the VN (A, B) but not in the DN (C); higher magnification (B) of double-labeled vestibular commissural neurons (*) in the outlined area in A. Inset in B shows GABA-immunopositive but retrogradely unlabeled neurons in the VN from the adjacent section; retrogradely labeled commissural neurons in the DN (green) were GABA immunonegative but were surrounded by dense GABA-immunopositive terminal-like structures (red). Insets I and II show higher magnifications of DN neurons in the outlined area (II) and from the adjacent section (I). D–F, Combined glycine immunohistochemistry (red) and retrograde tracing with Alexa Fluor 488 dextran (green) revealed the absence of double-labeled glycine-immunopositive commissural neurons (yellow) in the VN (D, E) but moderate numbers in the DN (F). Higher magnification (E) of vestibular commissural (green) and of glycine-immunopositive neurons (red) in the outlined area in D; the inset in E shows glycine-immunopositive neurons in the VN from the adjacent section. Several retrogradely labeled (green) commissural neurons in the DN (F) were also glycine immunopositive (red) and thus appear yellow; the inset in F shows a higher magnification of the DN neurons in the outlined area. White arrows in A, C, D, and F indicate the sulcus limitans. Scale bars: A, C, D, F, 250 μm; B, E, insets in E, F, and inset II in C, 50 μm; inset in B and inset I in C, 20 μm.
In contrast to the distinct subgroup of GABAergic vestibular commissural neurons, glycine-immunopositive commissural vestibular neurons were absent (Fig. 4D,E). This absence of glycinergic vestibular commissural neurons, however, is not related to a general absence of glycine-immunopositive neurons in the VN, because moderate numbers of glycine-immunopositive neurons (Fig. 4E, inset) were encountered mainly in the r4–r6 part of the DVN, compatible with previous results (Reichenberger et al., 1997). The absence of glycinergic vestibular commissural neurons was also not too surprising because glycine-immunopositive cells and commissural neurons in the VN comprise spatially mostly nonoverlapping subgroups of functional phenotypes, respectively (Fig. 3H). Because glycinergic neurons apparently do not contribute to vestibular commissural projections, the majority of the retrogradely labeled commissural neurons (those that are GABA-immunonegative) are likely excitatory, compatible with previous results from electrophysiological studies (Ozawa et al., 1974; Holler and Straka, 2001).
Interestingly, a complementary transmitter organization occurs for brainstem commissural neurons in the auditory nucleus. Commissural neurons in the DN include a considerable proportion of up to ∼35% glycine-immunopositive (Fig. 4F, yellow neurons) but no GABA-immunopositive (Fig. 4C) neurons. The apparent absence of the latter cell type and the generally small number of GABA-immunopositive neurons in this nucleus is contrasted by the presence of a dense arrangement of GABA-immunopositive terminal-like structures around the majority of DN neurons (Fig. 4C, insets I, II). This causes some DN cells to appear as ghost-like neurons, given the dense surrounding by GABA-immunopositive terminal-like structures (Fig. 4C, inset II).
Trajectory of vestibular commissural connections
Retrograde labeling of vestibular commissural connections indicated that the axons of the commissural neurons cross the midline of the brainstem in more or less distinct bundles (Fig. 5A) along the entire rostrocaudal extent of the vestibular nuclei. The axons of both vestibular and auditory commissural neurons form common, rather broad fiber tracts that cross the midline as an IAF or an EAF tract (Fig. 5B) (Grofová and Corvaja, 1972; Feng and Lin, 1996). The majority of fibers change their trajectory from an IAF to an EAF position and vice versa at the midline, whereas only few fibers remain in their respective bundle of origin (Fig. 5A,B). The course of the axonal trajectory by which commissural signals are mediated to contralateral vestibular neurons was determined by recording N.VIII-evoked synaptic inputs in identified 2°VN throughout the rostrocaudal extent of the vestibular nuclei in isolated frog whole brains in vitro before and after different combinations of bilateral longitudinal sections of the IAF and EAF tracts (Fig. 5C).

Organization of brainstem commissural fiber pathways. A, Coronal section showing labeled vestibular and auditory fibers crossing the midline of the brainstem after application of Alexa Fluor 546 dextran to the right VN; the inset shows a coronal section with the application site in the VN at the level of N.VIII. B, Schematic drawing illustrating the organization of brainstem crossing commissural pathways between the bilateral vestibular and auditory nuclei (modified from Grofová and Corvaja, 1972); axons of commissural vestibular and auditory neurons cross the midline as an IAF and an EAF tract. C, Histological reconstructions of coronal sections at the level of N.VIII and iN.VIII-evoked afferent (black traces) and cN.VIII-evoked commissural responses (red traces) in 2°VN recorded after the lesion; depth and extension of longitudinal cuts of the bilateral EAF (C1), IAF (C2), and alternate EAF/IAF (C3) is indicated by the red areas. Stimulus intensities for the afferent responses ranged from 1.4 to 2.3 × T and for the commissural responses from 3.0 to 4.8 × T; records are the average of 20 responses, respectively, except for the orthodromic action potentials in C1 and C2, which are single sweeps. Arrowheads in C1–C3 indicate stimulus onset and dashed lines the resting membrane potential of −65 mV in C1, −66 mV in C2, and −64 mV in C3 on the left side and −67 mV in C3 on the right side. Calibration of responses to left iN.VIII and left cN.VIII in C3 apply also to responses to right iN.VIII and right cN.VIII. c, Contralateral; i, ipsilateral; s.l., sulcus limitans.
Electrical stimulation of the ipsilateral N.VIII (Fig. 6A) evoked in control neurons (n = 39) monosynaptic EPSPs with a latency of 2.3 ± 0.3 ms, a time-to-peak of 3.8 ± 1.5 ms, and an amplitude of 4.1 ± 1.0 mV at a stimulus intensity of 1.5 × T (values are mean ± SE). At stimulus intensities >2.5 × T action potentials were superimposed on the EPSPs (Fig. 5C1,C2, iN.VIII). Commissural responses in these 2°VN were elicited by electrical stimulation of the contralateral N.VIII at 4.5 × T (Fig. 6A) and consisted of an excitatory response with a latency of 8.2 ± 1.1 ms and a rather variable time-to-peak (35.8 ± 23.1 ms) and amplitude (4.3 ± 2.4 ms). The onset latency of 8–9 ms is compatible with a disynaptic connection between electrically activated fibers in the N.VIII and contralateral 2°VN (Holler and Straka, 2001). Because an N.VIII stimulation recruits fibers from all labyrinthine end organs, the canal-specific commissural IPSPs in frog 2° semicircular canal neurons from the contralateral coplanar canal nerve (Fig. 6B4) (Holler and Straka, 2001) are masked by excitatory inputs from other end organs (Fig. 6B5,B6). Although the stimulation of the entire contralateral N.VIII is unspecific for canal commissural projections, it facilitated the study of particular contributions of the different arcuate fiber tracts to the mediation of vestibular commissural signals.

Synaptic organization of commissural inputs in phasic and tonic 2°VN. A, Schematic drawing illustrating the recording site in the right VN and the stimulation of individual semicircular canal nerve branches and the N.VIII on both sides. B, Commissural inputs in an identified phasic 2° AC neuron. B1, An initial subthreshold notch (black trace) and a single spike (gray trace) evoked by intracellular injection of a positive current step at two intensities (bottom traces) characteristic for phasic 2°VN; the inset in B1 shows the response onset at an extended timescale. B2, B3, iAC nerve-evoked monosynaptic afferent EPSP (B2) and cN.VIII-evoked disynaptic commissural EPSPs (B3); stimulus intensities in B2 (1.3, 2.0, 2.8 × T) and in B3 (2.1, 3.0, 4.0 × T). B4–B6, Separate stimulation of the three contralateral semicircular canal nerves evoked an oligosynaptic commissural IPSP from the coplanar cPC (B4) and disynaptic and oligosynaptic EPSPs from the two non-coplanar cAC (B5) and cHC (B6), respectively. Stimulus intensities are 2.5, 3.0, and 4.5 × T in B4, 2.2, 3.3, and 4.8 × T in B5, and 2.5, 3.8, and 5.0 × T in B6; note that all responses in B are from the same neuron. C, Coplanar semicircular canal commissural IPSPs in a tonic 2° PC neuron. C1, A continuous discharge in response to a positive current step (bottom trace) characteristic for tonic 2°VN. C2, Monosynaptic afferent EPSPs after electrical stimulation of the iPC nerve at 1.3 and 2.2 × T. C3, Disynaptic commissural IPSPs after stimulation of the coplanar cAC nerve at 2.2, 3.0, and 4.6 × T. D, Coplanar semicircular canal commissural EPSPs in a phasic 2° HC neuron. D1, A single spike in response to a positive current step (bottom trace) characteristic for phasic 2°VN. D2, Monosynaptic afferent EPSPs after electrical stimulation of the iHC nerve at 1.3 and 2.1 × T. D3, Commissural EPSPs after stimulation of the coplanar cHC nerve at 2.2, 3.0, and 4.6 × T with a disynaptic onset. E, Latency distribution of coplanar semicircular canal nerve-evoked commissural IPSPs (n = 138) and EPSPs (n = 51) in all recorded 2°VN; response latencies were distinguished into disynaptic (black bars) and oligosynaptic (>disynaptic, gray bars) onsets. Records in B1, C1, and D1 are single sweeps and in B2–B6, C2, C3, D2, and D3 the average of 20 responses. Arrowheads in B2–B6, C2, C3, D2, and D3 mark stimulus onset and dashed lines the resting membrane potential of the neurons, respectively; vertical gray bars in B3–B6, C3, and D3 indicate the mean ± SE of the disynaptic onset of the vestibular commissural field potential. Calibration in B2, and C2 apply to B3–B6 and C3, respectively. AC, Anterior canal; PC, posterior canal; HC, horizontal canal; c, contralateral; I, ipsilateral; CB, cerebellum; OT, optic tectum.
The longitudinal incisions that interrupted the IAF (n = 2) or EAF tracts bilaterally (n = 2) or the IAF tract on one and the EAF tract on the other side (n = 3) were histologically verified at the end of the experiments. Typical examples of reconstructed schematic coronal sections at the level of N.VIII are shown in the left column of Figure 5C1–C3. The longitudinal extent of the lesion (from ∼0.8 mm rostral to ∼2.2 mm caudal to the entry of the N.VIII) exceeded the rostrocaudal extent of the vestibular nuclei and all incisions were long and deep enough (Fig. 5C1–C3, outlined in red in the sections) to interrupt the IAF and EAF tracts, respectively (Grofová and Corvaja, 1972). After the longitudinal incisions, 107 neurons were recorded on both sides of brains with a bilateral IAF (26 cells) or EAF (37 cells) tract lesion or with an alternate IAF/EAF lesion on the two sides (44 cells). Ipsilateral N.VIII-evoked monosynaptic EPSPs (Fig. 5C1–C3, iN.VIII) in 2°VN in brains after the different types of lesions exhibited mean latencies, times-to-peak, and amplitudes at 1.5 × T that were similar to the control values, respectively.
After the different tract sections, electrical stimulation of the contralateral N.VIII was still able to evoke commissural responses in all 2°VN (Fig. 5C1–C3, cN.VIII). Independent of the type of the preceding lesion, the mean latency (8.5 ± 0.8 ms; n = 107) and time-to-peak (32.1 ± 16.1 ms; n = 107) of the commissural excitatory responses were similar to those of the corresponding control responses before the lesion. However, the amplitude of the commissural excitation was significantly smaller (p ≤ 0.0001, Mann–Whitney U test) compared with controls. At comparable stimulus intensities (4.5 × T), the amplitude (mean ± SE) was 2.1 ± 1.0 mV (n = 26) after bilateral IAF, 1.8 ± 0.8 mV (n = 37) after bilateral EAF, and 1.9 ± 0.7 mV (n = 44) after alternate IAF/EAF tract section on the two sides. The ∼50% reduction in the amplitude of the commissural EPSPs after the longitudinal sections suggests an additive effect of the crossing fibers in mediating commissural responses, because the lesions disrupted ∼50% of the crossing fibers in each case. This allows the conclusion that brainstem crossing vestibular commissural fibers are not organized into a distinct, single bundle but are more or less equally distributed among the entire midline-crossing IAF and EAF tracts.
Synaptic organization of the canal commissural inhibition
The organization of commissural semicircular canal inputs in 2°VN was studied in isolated frog whole brains in vitro in which the nerve branches to all six semicircular canals were stimulated separately with suction electrodes (Fig. 6A). All 2°VN were identified as semicircular canal neurons by a monosynaptic EPSP from an ipsilateral canal nerve (Fig. 6B2,C2,D2). Commissural inputs in these identified 2° canal neurons (Fig. 6B2) consisted of a disynaptic slow EPSP after electrical stimulation of the entire contralateral N.VIII (see above) (Fig. 6B3), compatible with previous in vivo (Ozawa et al., 1974) and in vitro (Cochran et al., 1987) studies. However, electrical activation of individual contralateral semicircular canal nerve branches revealed a more specific organization. The observed pattern consisted of a plane-specific commissural inhibition from the coplanar contralateral canal (Fig. 6B4) and an unspecific excitation from the two other non-coplanar canals (Fig. 6B5,B6). These latter commissural excitatory inputs from the other two canals thus mask the canal-specific commissural coplanar inhibition after electrical stimulation of the entire contralateral N.VIII (Holler and Straka, 2001).
A coplanar semicircular canal commissural inhibition (Fig. 6B4,C3) was encountered in the majority of identified 2° canal neurons (138 of 189; ∼70%), whereas the other 2° canal neurons (51 of 189; ∼30%) exhibited a commissural excitation that usually did not evoke action potentials even at higher stimulus intensities (Fig. 6D3). The activated commissural coplanar IPSPs and EPSPs after electrical stimulation of the contralateral coplanar canal nerve could be subdivided with respect to onset latency. Using previous calculations based on the relative timing of commissural field potentials, presynaptic action potentials (Fig. 7A,B1) and synaptic delay (Holler and Straka, 2001) commissural responses had either disynaptic (Fig. 6B5,C3,D3) or oligosynaptic (Fig. 6B4,B6) onset latencies. Plotting the onset distribution of coplanar commissural IPSPs and EPSPs in the recorded neurons revealed a predominance for disynaptic latencies (Fig. 6E, ■). This preponderance suggests that the major vestibular commissural inhibitory (103 of 138 neurons; ∼75%) and excitatory (35 of 51 neurons; ∼70%) connections are direct and involve brainstem-crossing projection neurons such as those that were retrogradely labeled and illustrated in Figure 1. Oligosynaptic pathways that require interneurons on the side of the recorded commissural responses (e.g., type II vestibular neurons) (Shimazu and Precht, 1966) are less prominent and supplement preceding disynaptic components.

Parameters of coplanar semicircular canal commissural inputs in phasic and tonic 2°VN. A, All-or-nothing axonal action potentials recorded in the vestibular nuclei after electrical stimulation of the cAC nerve at threshold intensity; the inset shows the onset at an extended timescale. The onset latency is compatible with presynaptic action potentials that evoke IPSPs in 2°VN with a disynaptic onset (dotted line, arrow). B, Coplanar semicircular canal commissural IPSPs in a tonic 2° PC neuron. B1, cAC nerve-evoked disynaptic commissural IPSPs at stimulus intensities of 2.1, 2.5, 3.2, 3.9, and 4.5 × T; stronger stimuli recruited longer-latency components (arrows). B2, Commissural cAC nerve-evoked (3.2 × T) IPSPs at different membrane potentials; changes in IPSP amplitude with membrane polarization were used to determine the reversal potential. B3, Monosynaptic EPSPs after iPC nerve stimulation at 1.2, 1.7, and 2.3 × T. B4, Continuous discharge in response to a positive current step (bottom trace) characteristic for tonic 2°VN; note that all responses in B are from the same neuron. C, Coplanar semicircular canal commissural IPSPs in a phasic 2° PC neuron. C1, Monosynaptic EPSP after iPC nerve stimulation at 1.6 × T. C2, Single spike discharge in response to a positive current step (bottom trace) characteristic for phasic 2°VN. C3, Commissural cAC nerve-evoked (4.5 × T) IPSPs at different membrane potentials; note that IPSPs did not reverse with membrane hyperpolarization. D, Relative proportion of commissural IPSPs and EPSPs in tonic and phasic 2°VN. E, Relative proportion of commissural IPSPs with disynaptic and oligosynaptic (>di) onsets in tonic and phasic 2°VN. F, Amplitude distribution of commissural IPSPs evoked with stimulus intensities of >4.5 × T in phasic and tonic 2°VN at resting membrane potential. G, Dependency of commissural IPSP amplitudes on membrane polarization in tonic and phasic 2°VN; IPSPs in tonic 2°VN reverse at approximately −74 mV (black dotted line, vertical arrow), whereas IPSPs in phasic 2°VN remain small and independent of the membrane potential (gray dashed line). Records in A, B4, and C2 are single sweeps and in B1–B3, C1, and C3 the average of 20 responses. Arrowheads in A, B1–B3, C1, and C3 mark stimulus onset and dashed lines the membrane potential, respectively; gray bars in B1, B2, and C3 indicate the mean ± SE of the disynaptic onset of the vestibular commissural field potential. Calibration in B1 applies to B2.
Differential organization of coplanar semicircular canal commissural IPSPs in phasic and tonic 2°VN
Frog 2°VN can be distinctly separated into a larger subgroup of phasic (Figs. 6B1,D1, 7C2) and a smaller subgroup of tonic (Figs. 6C1, 7B4) neurons based on their intrinsic membrane properties and discharge dynamics (Straka et al., 2004; Beraneck et al., 2007). Independent of these intrinsic response characteristics, commissural IPSPs from the contralateral coplanar semicircular canal were encountered in both types of 2° canal neurons (Fig. 6B,C), although in different proportions. Coplanar canal-specific commissural IPSPs were present in all tonic 2°VN (23 of 23 neurons; 100%) but only in a moderate number of phasic 2°VN (16 of 39 neurons; ∼40%) (Fig. 7D, ■). Moreover, the mainly small IPSPs in the latter phasic 2°VN were often followed by a pronounced excitatory component. The remaining phasic 2°VN (23 of 39 neurons; ∼60%) received coplanar canal-specific commissural EPSPs (Fig. 6D3), a pattern that was never encountered in tonic 2°VN (Fig. 7D). This suggests that the classical push–pull organization of coplanar semicircular canal signals is implemented for all tonic 2°VN but only in a subpopulation of phasic 2°VN. Such an arrangement, in which a contralateral excitation that forms part of a push–push organization complements the classical push–pull pattern in a subgroup of central vestibular neurons seems to be also present in the turtle vestibular commissural system (Ariel et al., 2004).
The onset latencies of the coplanar semicircular canal commissural IPSPs were mostly disynaptic (Fig. 7E, ■) in both tonic and phasic 2° canal neurons compatible with the results from nonclassified 2°VN (Fig. 6E). However, the maximal amplitude and its dependency from membrane polarization differed considerably in the two types of 2°VN. Maximal amplitudes of commissural IPSPs (at 4.5 × T) in tonic 2°VN at resting membrane potential (mean ± SE, 68.2 ± 2.1 mV; n = 23) were variable and usually >2–3 mV (Fig. 7B1). In contrast, commissural IPSPs (at 4.5 × T) in all phasic 2°VN at resting membrane potential (mean ± SE, 67.3 ± 1.7 mV; n = 16) were rather small (Fig. 7C3) and ranged between 0.3 and 1.2 mV. This difference became more obvious after plotting the distribution of the commissural IPSP amplitudes in phasic and tonic 2°VN (Fig. 7F). In addition, the amplitude of commissural IPSPs in tonic 2°VN depended on the actual membrane potential and changed with polarization. In the latter type of neurons, the IPSPs usually reversed after hyperpolarization of the membrane potential beyond −74 mV and became depolarizing (Fig. 7B2, black dotted line in G). In contrast, in phasic 2°VN, IPSP amplitudes remained rather small and were independent from the membrane potential (Fig. 7C3, gray dashed line in G), suggesting a more remote, dendritic origin of the commissural inhibitory input.
Intrinsic response characteristics of semicircular canal vestibular commissural neurons
Semicircular canal commissural IPSPs could be mediated by tonic, phasic, or both types of brainstem crossing commissural 2°VN (Fig. 8A, 1–3). The different options were studied in an electrophysiological and modeling approach that made use of the distinctly different discharge dynamics of phasic and tonic 2°VN after semicircular canal nerve stimulation with sinusoidally modulated pulse trains (Pfanzelt et al., 2008). In tonic 2°VN, the repetitively applied single pulses activated action potentials throughout the entire length of the stimulus train (Fig. 8B1). In contrast, phasic 2°VN fired only few action potentials during the first single pulses in the train (Fig. 8B2). Thus, depending on the distinctly different timing of the discharge in the two types of 2°VN (Fig. 8B3) that potentially could form commissural connections, contralateral semicircular canal nerve-evoked inhibitory responses in 2°VN will exhibit different time courses.

Characterization of the intrinsic response dynamics of vestibular commissural neurons. A, Schematic drawing illustrating possible commissural connections with respect to the intrinsic response dynamics of tonic (gray) and phasic (black) 2°VN; disynaptic commissural responses could be mediated by tonic (1), phasic (3), or both types of 2°VN (2). B, Typical responses of a tonic (B1) and a phasic (B2) 2°VN to stimulation of the iAC (B1) and iHC (B2) nerve with a train of single electrical pulses (at 2.9 × T in B1 and at 3.2 × T in B2) that were sinusoidally modulated in frequency between 0 and 70 Hz (bottom traces); normalized rate of evoked spike discharge (B3) during pulse trains with peak frequencies of 70 Hz in tonic (dashed line) and phasic (solid line) 2°VN (adopted from Pfanzelt et al., 2008). C, Commissural compound IPSP of a tonic 2° AC neuron after electrical stimulation of the coplanar cPC nerve by a sinusoidally modulated pulse train (at 4.0 × T) with a peak frequency of 70 Hz (bottom traces); the inset shows the IPSP to the first single pulse (gray area) of the train (open arrowhead) at an extended timescale. Double arrows indicate the activation of individual IPSPs by the last two single pulses of the train. D, Model compound commissural inhibition in a tonic 2°VN after sinusoidal pulse train stimulation at a peak frequency of 70 Hz (bottom traces) with the assumption that the response was mediated only by phasic (D1), only by tonic (D3), or by equal proportions of both types of 2°VN (D2); the parameters to generate the model responses were obtained from the commissural IPSPs of the neuron shown in C at a resting membrane potential of −67 mV (see also Materials and Methods); the arrowhead in D1 indicates the truncation of the model response by the termination of the presynaptic discharge of putative phasic commissural 2°VN. E, Commissural compound EPSP of a phasic 2° PC neuron after electrical stimulation of the coplanar cAC nerve by a sinusoidally modulated pulse train (at 3.8 × T) with a peak frequency of 40 Hz (bottom traces); double arrows indicate the activation of individual EPSPs by the last two single pulses. Records in B1 and B2 are single sweeps and in C the average of 10 responses; dashed lines indicate the resting membrane potential, respectively; Calibration in B1 and D3 apply to B2 and D1, D2, respectively. aff, Afferent; com, commissural; s.c., semicircular canal.
The responses after electrical stimulation of the contralateral coplanar semicircular canal nerve with a sinusoidally modulated pulse train consisted of a temporal summation of individual disynaptic commissural IPSPs (Fig. 8C, inset) evoked by each single pulse of the train (Fig. 8C). Because it is almost impossible to simultaneously record presynaptic commissural fibers and their respective postsynaptic vestibular target neurons, a modeling approach was used (Pfanzelt et al., 2008). This method took into account the synaptic delay and time constants of the recorded commissural responses to single pulses (see Materials and Methods) for a calculation of compound commissural inhibitory responses based on the differential discharge dynamics of presynaptic tonic or phasic 2°VN during a sinusoidally modulated pulse train (Fig. 8B3). The minimal cellular models for tonic and phasic 2°VN consisted of a soma and a three compartmental dendrite with uniformly distributed conductances that were generated from frequency domain data of the respective type of 2°VN (Pfanzelt et al., 2008; Rössert et al., 2009a,b). Based on this approach, a model compound semicircular canal commissural inhibition was generated assuming its mediation by phasic (Fig. 8D1), tonic (Fig. 8D3), or in equal proportions by both types of brainstem crossing commissural vestibular neurons (Fig. 8D2). Comparison of the time courses of the calculated model responses (Fig. 8D1–D3) with the recorded commissural compound IPSP (Fig. 8C) suggests that the latter was primarily evoked by the presynaptic activity of tonic commissural 2°VN, although a somewhat minor contribution of phasic commissural 2°VN during the initial part of the pulse train-evoked compound response cannot be entirely ruled out. A major contribution of tonic 2°VN to the vestibular commissure also complies with the finding that the last single electrical pulses of the stimulus train always elicited noticeable IPSPs (n = 6) (Fig. 8C, double arrows). This would not have been the case if exclusively phasic 2°VN had mediated the response because their spike discharge dynamics is temporally restricted to the first few pulses of the train (Fig. 8B2,B3). A corresponding activation of commissural EPSPs by the last pulses of the train was also observed for the coplanar canal commissural compound excitation in phasic 2°VN (n = 7) (Fig. 8E, double arrows). This suggests that brainstem-crossing inhibitory as well as excitatory commissural pathways, linking bilateral coplanar semicircular canal neurons, are established predominantly by tonic 2°VN.
Pharmacological profile of semicircular canal commissural inhibition
The vestibular commissural excitation is most likely mediated by glutamate that activates contralateral vestibular neurons through both AMPA and NMDA receptors as shown previously in the isolated frog brain (Cochran et al., 1987; Knöpfel, 1987). The relative contributions of AMPA and NMDA components to the commissural EPSPs varied between the different studies and depended on the Mg2+ concentration in the bath. The substantial number of GABAergic and the virtual absence of glycinergic vestibular commissural neurons (Fig. 4) suggest that the dominating transmitter for eliciting commissural IPSPs is GABA. Compatible with the immunohistochemical results, bath application of the glycine-receptor blocker strychnine (2 μm) (Fig. 9A1) and of the GABAA-receptor blocker bicuculline (5 μm) (Fig. 9A2) revealed a differential effect of the two antagonists on contralateral semicircular canal-evoked commissural field potentials. Strychnine only marginally increased longer-latency excitatory field potential components, whereas bicuculline distinctly augmented the excitatory components at the earliest possible onset (Fig. 9A1,A2, red traces). Because extracellularly recorded vestibular commissural field potentials are a composite of excitatory and inhibitory postsynaptic response components with different onset latencies, this result suggests that GABA activates disynaptic and oligosynaptic IPSPs, whereas glycine is implicated in the mediation of longer-latency commissural inhibitory components.

Pharmacological profiles of semicircular canal commissural responses in phasic and tonic 2°VN. A, cAC nerve-evoked commissural field potentials in the vestibular nuclei before (black traces; control), during subsequent application of 2 μm strychnine (red trace in A1) and 5 μm bicuculline (red trace in A2), and after washout (gray traces; wash). B–E, Effect of glycinergic and GABAergic antagonists on coplanar semicircular canal nerve-evoked commissural responses in four different 2°VN; neurons were identified as 2° semicircular canal neurons by monosynaptic EPSPs from the iHC (B1), iPC (C1), or iAC (D1, E1) nerves and characterized as tonic 2°VN by a continuous discharge (B2, C2, E2) or as phasic 2°VN by a single spike (D2) in response to intracellular injected positive current pulses (indicated by bottom gray traces: 0.5 nA in B3, E3; 0.4 nA in C3; 0.7 nA in D3). B3, C3, D3, Effect of 2 μm strychnine on semicircular canal nerve-evoked commissural inhibition (gray traces, respectively); strychnine leaves disynaptic responses unaffected but reduces longer-latency components (black arrows in B3, C3, D3). C3, D3, Effect of 5 μm bicuculline and 2 μm strychnine on semicircular canal nerve-evoked commissural responses (red traces, respectively); bicuculline blocks all IPSP components including those with a disynaptic onset (red arrow in C3) and unmasks or increases EPSPs with a disynaptic onset in phasic 2°VN (red arrow in D3). E3, Effect of 100 μm CGP 35348 (gray trace) and combined application of 100 μm CGP 35348 and 5 μm bicuculline (red trace) on semicircular canal nerve-evoked commissural responses; CGP 35348 slightly reduces the IPSP amplitude, whereas bicuculline in the presence of CGP 35348 blocks the disynaptic IPSP completely and unmasks an EPSP with a disynaptic onset (red arrow). F, Proportion of coplanar canal commissural IPSPs with a disynaptic (F1) or oligosynaptic onset (>di; F2) that were reduced by bicuculline (Bic), CGP 35348 (CGP), or strychnine (Stry), respectively. Numbers in F indicate recorded neurons. Records in B2, C2, D2, and E2 are single sweeps and in A1, A2, B1, B3, C1, C3, D1, D3, E1, E3 the average of 20 responses. Arrowheads in A, B1, B3, C1, C3, D1, D3, E1, E3 mark stimulus onset and dashed lines in A the baseline and in B–E the resting membrane potential. AC, Anterior canal; PC, posterior canal; HC, horizontal canal; c, contralateral; i, ipsilateral.
The differential impact of GABA and glycine blockers on semicircular canal commissural IPSPs in individual neurons was evaluated by intracellular recordings (n = 27) of monosynaptically identified (Fig. 9B1,C1,D1,E1) and dynamically characterized (Fig. 9B2,C2,D2,E2) tonic and phasic 2°VN. Bath application of 2 μm strychnine decreased the amplitude of semicircular canal commissural IPSPs in tonic 2°VN (Fig. 9B3,C3, gray traces) and increased the amplitude of excitatory components in phasic 2°VN (Fig. 9D3, gray trace). However, the onset of the reduced glycinergic inhibitory component (n = 8) and of the increased excitatory component (n = 3) was always delayed with respect to the disynaptic onset of the commissural responses (Fig. 9B3,C3,D3, black arrows; B3, inset). This indicates that glycine does not account for the disynaptic component of the canal commissural inhibition (Fig. 9F1), compatible with the absence of glycinergic vestibular commissural neurons (Fig. 4D,E). However, glycine mediates, at least in part, delayed inhibitory commissural components through oligosynaptic pathways (4 of 5 neurons, 80%) (Fig. 9F2) that are mediated by excitatory brainstem-crossing vestibular commissural neurons and local type II inhibitory vestibular interneurons (Holler and Straka, 2001).
In contrast, bath application of 5 μm bicuculline blocked the semicircular canal commissural IPSPs in all tonic and phasic 2°VN (Fig. 9C3,D3, red traces), including the initial components with a disynaptic onset (Fig. 9C3,D3, red arrows; D3, inset) that were not affected by strychnine in the same neurons. The almost complete block of the disynaptic canal commissural IPSP components by the GABAA blocker (Fig. 9F1) complies with the presence of GABAergic vestibular commissural neurons (Fig. 4A,B) and suggests a major role for GABA as inhibitory transmitter of semicircular canal commissural signals in 2°VN. The block of IPSPs by combined application of strychnine and bicuculline (Fig. 9D3) unmasked in the majority of the recorded 2°VN (6 of 10 neurons) an excitatory component with a disynaptic onset latency. Because bicuculline also reduced all canal commissural IPSPs with an oligosynaptic onset (data not shown), it is likely that these longer-latency inhibitory components, which require excitatory vestibular commissural neurons and local inhibitory interneurons, are mediated by GABA as well as by glycine (Fig. 9F2).
Bath application of the GABAB receptor antagonist CGP 35348 (100 μm) was used to reveal a possible contribution of GABAB receptors to the canal commissural GABAergic inhibition (Fig. 9E3). This GABA receptor subtype is implicated in the postlesional plasticity of the vestibular commissural circuitry after unilateral labyrinthectomy (Johnston et al., 2001; Bergquist et al., 2008). Application of CGP 35348 reduced the amplitude of the canal commissural IPSPs with a disynaptic onset (Fig. 9E3, gray trace) in part of the tonic and phasic 2°VN by ∼10–15% (3 of 5 neurons, 60%) (Fig. 9F1). Combined application of CGP 35348 and bicuculline blocked the disynaptic IPSPs completely (Fig. 9E3, red trace) and again unmasked an excitatory canal commissural response component with the same onset latency (Fig. 9E3, red arrow). Thus, based on the pharmacological study, the semicircular canal commissural inhibition is mainly mediated disynaptically by GABAergic inhibitory commissural neurons through an activation of GABAA and GABAB receptors and supplemented by an oligosynaptic pathway that requires excitatory commissural neurons that activate local glycinergic and GABAergic type II vestibular interneurons.
Discussion
Vestibular brainstem commissural projections originate as distinct cell groups from specific hindbrain segments that coincide with the SVN, MVN, and DVN. The mostly disynaptic coplanar semicircular canal commissural inhibition is mediated by midline-crossing GABAergic tonic 2°VN and supplemented by oligosynaptic GABAergic and glycinergic IPSPs. Large-amplitude GABAA receptor-mediated commissural IPSPs are activated in all tonic 2° canal neurons, whereas pronounced coplanar canal commissural EPSPs predominate in phasic 2° canal neurons. This suggests that the canal commissural inhibition is dynamically specific and essentially involves 2°VN with rather linear response properties.
Conserved ontogenetic origin of vestibular commissural pathways in vertebrates
Comparison of origin and trajectory of vestibular commissural neurons across different vertebrate groups indicates a highly conserved organizational pattern (Grofová and Corvaja, 1972; Gacek, 1978; Epema et al., 1988; Newlands et al., 1989; Büttner-Ennever, 1992; Highstein and Holstein, 2006). The majority of brainstem-crossing vestibular commissural neurons is located in the SVN, MVN, and DVN and coincides with an ontogenetic hindbrain segmental origin from r2/r3 and r5–r7, respectively (Fig. 10A) (Cambronero and Puelles, 2000; Straka et al., 2001). The localized projections of vestibular commissural neurons to contralateral vestibular targets (Figs. 1D–F, 3D) provide the morphological basis for specific reciprocal connections that link bilateral vestibular subpopulations with spatially corresponding head motion signals. The virtual absence of commissural neurons from the magnocellular part of the LVN (Gacek, 1978; Straka et al., 2001) primarily excludes r4 as an origin of the vestibular commissure. Because vestibular commissural neurons are generally rather small with low-dynamic membrane properties, as shown here, the absence of commissural neurons in the LVN complies with the fact that this latter nucleus is dominated by large to giant neurons with particularly phasic response dynamics (Uno et al., 2003).

Summary diagram depicting the major organizational principles of the vestibular commissure. A, Top view of a schematic hindbrain (left side) illustrating the rostrocaudal arrangement of the vestibular nuclei along the rhombomeric scaffold (A1) and of distinct vestibular subgroups with unique projections to major vestibulo-motor-related targets (A2), suggesting a feedforward push–pull organization of the vestibular commissure. B, Physio-pharmacological organization of pathways mediating a coplanar semicircular canal commissural inhibition in tonic (T) 2°VN (B1) and a coplanar semicircular canal commissural excitation in phasic (P) 2°VN (B2). VC, Vestibulo-cerebellar neurons; VCom, vestibular commissural neurons; VN, vestibular nucleus; VO, vestibulo-ocular neurons; VS, vestibulo-spinal neurons. Green neurons are excitatory, red neurons are inhibitory, and orange neurons are excitatory or inhibitory.
The SVN, MVN, and DVN not only give rise to vestibular commissural neurons but also to vestibulo-ocular, vestibulo-spinal, and cerebellar-projecting neurons (Fig. 10A). In particular, the large SVN subgroup of commissural neurons (Figs. 2, 3) (Gacek, 1978; Büttner-Ennever, 1992) overlaps with oculomotor and trochlear nucleus-projecting anterior and posterior vertical semicircular canal neurons (Straka et al., 2005). However, all commissural neurons projected to single targets without collaterals to other known vestibular projection sites (Fig. 10A). This is particularly noticeable because inhibitory 2°VN that terminate on vertical and oblique extraocular motoneurons in the oculomotor and trochlear nucleus are GABAergic (Precht et al., 1973b; Cochran, 1992), as are vestibular commissural neurons located in the same portion of the SVN (Figs. 1I–K, 3A–C). This finding, however illustrates that the signals for ipsilateral oculomotor/trochlear motoneurons and for contralateral coplanar semicircular canal neurons are mediated by separate subsets of inhibitory GABAergic neurons (Fig. 10A). Thus, canal commissural inhibitory connections, at least those that interconnect bilateral vertical coplanar semicircular canal neurons, are organized in a feedforward push–pull and not in a feedback closed-loop mode as assumed previously for the evaluation of the role of commissural pathways in vestibular compensation (Galiana and Outerbridge, 1984; Galiana et al., 1984).
Pharmacological profile of vestibular commissural pathways
The initial in vivo study in cat suggested that GABA and glycine mediate a horizontal canal commissural inhibition (Precht et al., 1973a), whereas an exclusive GABAergic commissural effect was reported in rat slice preparation in vitro (Furuya et al., 1991). A purely disynaptic GABAergic commissural inhibition that is supplemented by oligosynaptic GABAergic and glycinergic commissural inhibitory responses, mediated through type II vestibular neurons, as found in the present study (Fig. 10B1) complies with both results given that longer-latency pathways in slice preparations are potentially impaired. Moreover, all available pharmacological data in intact and unilateral labyrinthectomized animals suggest GABA as the major transmitter of the vestibular commissural inhibition, which is predominantly mediated by GABAA and to a minor extent by GABAB receptors, compatible with the presence of both receptor subtypes on central vestibular neurons (Yamanaka et al., 2000; Graham and Dutia, 2001; Bergquist et al., 2008). In view of the dominant role of the GABAergic vestibular commissural inhibitory pathway, the presumed newly born ipsilesional GABAergic neurons after unilateral neurectomy (Tighilet et al., 2007) could theoretically contribute to the recalibration of a balanced resting activity between the bilateral vestibular nuclei, given that the new cells are not local type II vestibular neurons as assumed previously but GABAergic commissural neurons.
At variance with the absence of glycinergic vestibular commissural neurons in frog (Fig. 4D,E), such a population has been demonstrated in mouse (Bagnall et al., 2007). Given otherwise large conserved similarities of basic organizational principles of the vertebrate vestibular system, this difference might be related to behavioral differences such as the presence of a velocity storage mechanism in mammals (Holstein et al., 1999) and its clear absence in frog (Straka and Dieringer, 2004). Based on fundamental differences in locomotor performance of frog and mouse, an additional set of glycinergic vestibular commissural neurons in the latter might also be an expression of the necessity for a more efficient commissural push–pull modulation of vestibular activity during the more or less continuous mammalian-type locomotion. Alternatively, GlyT2 transporter-expressing vestibular neurons in the YFP-16 mouse (Bagnall et al., 2007) might be a specific population of excitatory vestibular commissural neurons that colocalize glutamate and glycine as a subgroup of frog and rat vestibular nerve afferents (Reichenberger and Dieringer, 1994), although mouse GlyT2-positive vestibular neurons were VGluT2 negative.
Functional organization and physiological profile of vestibular commissural neurons
The plane-specific inhibitory commissural projection that interconnects bilateral coplanar semicircular canal neurons in cat (Kasahara and Uchino, 1971) and frog (Holler and Straka, 2001) increases the sensitivity of angular head acceleration-related responses in central vestibular neurons (Shimazu and Precht, 1966; Markham et al., 1977). The anatomical pathway that links the bilateral vestibular nuclei is classically illustrated as trisynaptic with a local type II inhibitory interneuron (Shimazu and Precht, 1966). Some of these local type II inhibitory neurons might also be involved in mediating the canal-specific ipsilateral feedforward inhibition (Fig. 10B1) (Minor and Goldberg, 1991; Straka et al., 1997) because ∼50% of these neurons were excited by the ipsilateral N.VIII (Shimazu and Precht, 1966). Although such a trisynaptic commissural connectivity is clearly implemented, in particular for horizontal semicircular canal neurons, the commissural inhibition is dominated by a direct midline-crossing disynaptic pathway in cat (Uchino et al., 1986) as in frog (Holler and Straka, 2001).
The coplanar canal-specific commissural inhibition is supplemented by a commissural excitation in a considerable number of vestibular neurons in frog (Holler and Straka, 2001), turtle (Ariel et al., 2004), guinea pig (Babalian et al., 1997), mouse (Broussard, 2009), and monkey (Broussard and Lisberger, 1992). In the latter species, the relative proportion of afferent and commissural vestibular excitation in horizontal canal-related floccular target neurons (FTNs) has been suggested to be important for vestibulo-ocular motor plasticity (Broussard and Lisberger, 1992). In mammals, these FTNs form a specific subgroup of type B vestibular neurons (Babalian and Vidal, 2000; Sekirnjak et al., 2003) and are thus functionally equivalent to frog phasic 2°VN (Straka et al., 2005). The majority of the latter frog vestibular neurons also receive, as do monkey FTNs, a bilateral vestibular excitation (Figs. 7D, 10B2), suggesting a conserved basic organizational vertebrate pattern.
The present results indicate that the inhibitory GABAergic vestibular commissure, which forms the major component of the commissural inhibition, is mediated in frog by tonic 2°VN (Figs. 8, 10B) and in mouse by neurons with low response dynamics (GIN neurons; Bagnall et al., 2007). Based on physiological parameters, these latter neurons coincide with mammalian type A vestibular neurons (Takazawa et al., 2004; Bagnall et al., 2007; Gittis and du Lac, 2007), which are functionally equivalent to tonic 2°VN in frog (Straka et al., 2005). Thus, the GABAergic commissural inhibition in vertebrates appears to be generally mediated by cells with rather linear response dynamics that are ideally suited for a gradational control of the sensitivity of central vestibular neurons over a larger range. In mammals, this pathway is complemented by commissural projections that involve type B-like excitatory and likely also inhibitory vestibular neurons (Bagnall et al., 2007) with response dynamics like those of oculomotor-projecting vestibular neurons (Sekirnjak and du Lac, 2006). Although it is possible that frog phasic 2°VN also contribute to vestibular commissural projections, this latter neuronal subtype persistently receives a pronounced commissural excitation through tonic 2°VN (Fig. 10B2). Commissural connections that consist of low-dynamic inhibitory and of higher-dynamic excitatory vestibular neurons have been suggested for the turtle to explain changes in head rotation-evoked discharge modulation in vestibular neurons after blocking labyrinthine afferent activity (Ariel et al., 2004). The presence of afferent and commissural excitatory inputs in central semicircular canal neurons might be rather common given their occurrence in several vertebrate species. Thus, the traditional scheme of the semicircular canal commissure as a push–pull system as initially proposed by Shimazu and Precht (1966) needs to be complemented by a pronounced canal commissural excitation. Functionally, the relative synaptic weight of excitatory feedforward circuits in combination with a disynaptic ipsilateral afferent (Straka et al., 1997) and a commissural inhibition is likely to play the essential role in the control and fine-tuning of sensitivity and dynamics of central vestibular activity.
Footnotes
-
This research was supported by the French Centre National de la Recherche Scientifique and the Centre National d'Etudes Spatiales. We greatly appreciate the opportunity to use a GABA antibody generated by the laboratory of Dr. G. R. Holstein. Thanks to Dr. Mathieu Beraneck for critically reading this manuscript.
- Correspondence should be addressed to Dr. H. Straka, Division of Neurobiology, Department Biology II, Ludwig-Maximilians-University München, Grosshadernerstrasse 2, 82152 Planegg, Germany. straka{at}lmu.de





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
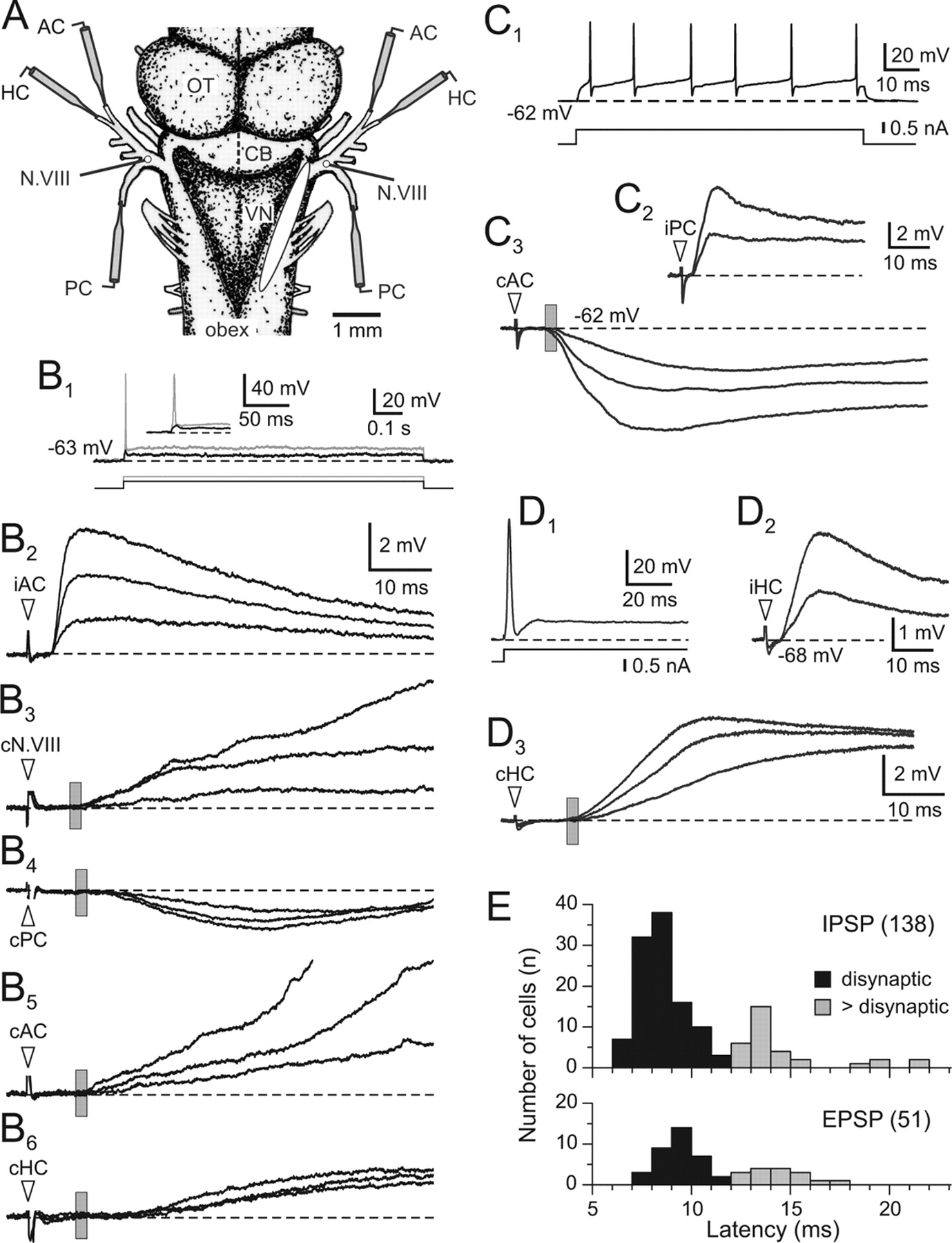{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}