

Abstract
Spatiotemporal properties of dopamine release play a major role both in striatal and nigral physiology because dopamine is released from nerve terminals and dendrites of nigrostriatal dopaminergic (DA) neurons. Pioneering work revealed gap junctional communication (assessed by dye-coupling experiments) between DA cells in the substantia nigra pars compacta (SNc). However, direct evidence of functional electrical synapses between DA neurons is still lacking. In this study, gap junctional communication between DA neurons was investigated in rat brain slices. Tracer coupling was observed in postnatal day 5 (P5) to P10 and P15-P25 rats. Dual whole-cell patch-clamp recordings revealed that 96% of DA neurons were coupled by electrical synapses in P7-P10 rats, and 20% were coupled in P17-P21 rats. These electrical synapses were mainly symmetrical and displayed strong low-pass filtering properties. When spontaneous firing activity was monitored, no significant synchronization was observed. Nevertheless, an efficient modulation of the spontaneous firing frequency of the postsynaptic cell occurred during injection of hyperpolarizing and depolarizing currents in the coupled presynaptic cell. Together, these observations demonstrate the existence of a fast communication between SNc DA neurons through electrical synapses.
- dopaminergic neuron
- substantia nigra pars compacta
- electrical synapse
- paired recordings
- gap junctions
- connexin
Introduction
The basal ganglia are a highly interconnected network of subcortical nuclei involved in adaptative control of behavior, in which the substantia nigra pars compacta (SNc) constitutes the main modulatory nucleus (Graybiel et al., 1990; Gerfen, 1992). The degeneration of nigrostriatal dopaminergic (DA) cells leads to Parkinson's disease (for review, see Obeso et al., 2000). DA neurons composing SNc mainly project to the dorsal striatum, a major input area of basal ganglia that selects relevant cortical information (Wilson, 1995). Dopamine potently modulates the processing of corticostriatal information (Reynolds and Wickens, 2002; Guzman et al., 2003).
Nigrostriatal DA neurons display two modes of discharge: a tonic background firing maintaining ambient dopamine levels in the striatum and a phasic firing (bursting activity) associated with brief high peaks of dopamine release, observed during appetitive learning and motivation-related behaviors (Gonon, 1988; Romo and Schultz, 1990; Ljungberg et al., 1992; Schultz et al., 1997).
Locally, dopamine released from dendrites regulates DA cell activity through DA autoreceptors (Skirboll et al., 1979; Cheramy et al., 1981). In addition, another mode of communication between DA neurons could be mediated by gap junctions. Indeed, an increasing body of evidence shows the existence of functional electrical synapses throughout the CNS not only between GABAergic interneurons (Gibson et al., 1999; Venance et al., 2000; Galarreta and Hestrin, 2001) but also between subsets of output neurons (Christie et al., 1989; Schmitz et al., 2001; Devor and Yarom, 2002; Venance et al., 2004). A pioneering study showed a dye coupling between DA neurons of the SNc in vivo in adult rats (Grace and Bunney, 1983b), suggesting the existence of an intercellular communication. Recent studies report the expression of connexins (Cx) in DA neurons in juvenile rats (Leung et al., 2002) but also an absence of dye coupling in these cells (Lin et al., 2003). Thus, gap junctional communication between DA neurons remains to be clarified. Furthermore, direct evidence of functional electrical synapses between DA neurons is still lacking.
Using tracer and electrical coupling experiments on rat brain slices, we directly measured electrical coupling for the first time between DA neurons of the SNc. DA neurons of SNc were found to be mainly interconnected by symmetrical electrical synapses that display low-pass filtering properties. Although no significant spontaneous synchronous activity was observed between pairs of DA neurons, electrical synapses were shown to modulate the firing frequency of electrically coupled DA neurons.
Materials and Methods
Electrophysiological recordings. Whole-cell recordings were done in coronal Sprague Dawley rat [postnatal day 5 (P5) to P25] brain slices (350 μm thick). All experiments were performed in accordance with the local ethical committee and European Union guidelines (directive 86/609/European Economic Community). Whole-cell recordings were made using borosilicate glass pipettes of 4-7 MΩ resistance containing the following (in mm): 105 K-gluconate, 30 KCl, 10 HEPES, 10 phosphocreatine, 4 ATP-Mg, 0.3 GTP-Na, 0.3 EGTA (adjusted to pH 7.35 with KOH). In experiments using Cs+-based intracellular solution, the composition was identical, except that K-gluconate and KCl were replaced by Cs-gluconate and CsCl, respectively (pH was adjusted to 7.35 with CsOH). The composition of the extracellular solution was as follows (in mm): 125 NaCl, 2.5 KCl, 25 glucose, 25 NaHCO3, 1.25 NaH2PO4, 2 CaCl2, 1 MgCl2, 10 μm pyruvic acid (sodium salt), and 10 μm 6,7-dinitroquinoxaline-2,3-dione (DNQX), 10 μm bicuculline methiodide, and 4 μm (+)-5-methyl-10,11-dihydro-5H-dibenzo [a,d] cyclohepten-5,10-imine maleate (MK-801) bubbled with 95% O2 and 5% CO2. All whole-cell recordings were performed at 34°C using a temperature control system (Bioptechs DTC3; Bioptechs, Butler, PA). Signals were amplified using an EPC9-2 amplifier (HEKA Elektronik, Lambrecht, Germany). Current-clamp recordings were filtered at 2.5 kHz and sampled at 5 kHz, whereas voltage-clamp recordings were filtered at 5 kHz and sampled at 10 kHz using the program Pulse-8.50 (HEKA Elektronik). Series resistance compensation was set to 75-90% in whole-cell configuration. Liquid junction potential error was calculated (-13.6 mV) according to the formula by Barry and Lynch (1991). The distance between two simultaneously recorded DA neurons was in the range of 10-50 μm. Except for DNQX (Tocris Cookson, Fischer Scientific, Illkirch, France), all chemicals (bicuculline methiodide, MK-801, carbenoxolone) were from Sigma (St. Quentin Fallavier, France).
Data analysis. Off-line analysis was performed using PulseFit-8.50 (HEKA Elektronik) and Igor Pro (Wavemetrics, Lake Oswego, OR). All results were expressed as mean ± SEM, and statistical significance was assessed using the Student's t test, or the nonparametric Mann-Whitney test when appropriate, at the significance level p < 0.05. Action potential (AP) threshold was measured as follows: DA neurons were first clamped to -60 mV, and successive depolarization steps (10 pA steps) were applied until the first AP, on which the threshold was then measured (see Fig. 1 A). Input resistance (Rinput) was calculated from voltage responses obtained after injecting a hyperpolarizing current (-10 pA; 1 sec duration). “Sag” amplitude was measured from voltage responses obtained after injecting a hyperpolarizing current (-90 pA; 1 sec duration; cell held previously at -60 mV) between the potential at sag peak (see Fig. 1, ○) and the potential at steady state (see Fig. 1, •). Spike duration was measured between the onset of the spike and the equipotential point during the repolarization phase. Fast afterhyperpolarization amplitude was taken between this last point and the minimum of the afterhyperpolarization. Coupling coefficient (e.g., k12) was calculated as the ratio of voltage responses of the postsynaptic (receiving) cell (here, cell 2) to the presynaptic (stimulated) cell (here, cell 1). For hyperpolarizing or depolarizing pulses, the amplitude of the presynaptic voltage response was measured between the holding membrane potential and the steady state after the sag (estimated 450 msec after stimulus onset), or the threshold of the first spike, respectively, for each stimulus. By convention, k12 was chosen as the higher k (i.e., the cell with the higher k to the other cell was chosen as cell 1). Using this convention, K ratio was defined as k12/k21 and Rinput ratio as R22/R11. Junctional conductance (Gj) was estimated as follows: Gj12 = k12/(R22 - R11k122). Rectification of electrical synapses was investigated by measuring Gj for depolarizing and hyperpolarizing stimuli in each direction of coupling (Gj for depolarizing stimuli, Gjd12 and Gjd21; Gj for hyperpolarizing stimuli, Gjh12 and Gjh21) and then comparing ratios of Gjd12/Gjd21 and Gjh12/Gjh21. When sinusoidal stimuli were used, 50-100 responses were averaged and subjected to a fast Fourier transform to confirm that the major frequency of the response was the frequency of the applied stimulus.

Electrophysiological characterization of DA neurons and tracer coupling between DA neurons. A, Responses of a DA neuron to current injections [-90, -60, -30, 0, +30 pA, left panel; +70 pA, right panel (same cell); holding potential was -60 mV, with -50.7 pA injected current]. The arrow represents where the AP threshold was estimated (at the point where the rate of rise accelerates sharply). Inset, Infrared image of an electrophysiologically identified DA neuron. Scale bar, 20 μm. B, Sag peak (○) and steady state (•) I-V relationship (mean of 21 DA neurons). C, Tracer coupling between SNc neurons in a coronal P17 rat brain slice. The electrophysiologically identified DA neuron (black arrow) was filled with a tracer (biocytin) that diffused into another neuron (white arrow) through gap junctions. Scale bar, 100 μm. D, Evolution of incidence of tracer coupling and number of coupled neurons during postnatal development. Fischer's exact test; *p < 0.05.
The relationship between the activity of each pair of cells was quantified using the following procedure. First, recordings were encoded into binary sequences: for each time point, subthreshold activity was encoded as 0 and spikes as 1. Second, mutual information between binary sequences X and Y of the cell pairs (i.e., the predictability of the activity of a cell from knowing the activity of the coupled cell) was computed as I(X:Y) = H(X) + H(Y) - H(X, Y), where H(X) is the entropy of the recording X, and H(X, Y) is the joint entropy of the recordings X and Y (Shannon, 1948). Entropy of the recording X is given by the following: 
where s(X) and p[s(X)] denote the state of recording X (i.e., 0 for subthreshold activity and 1 for spikes) and the probability of s(X), respectively. Similarly, joint entropy of simultaneous recordings X and Y is given by the following: 
Finally, statistical significance of the mutual information measurement was tested using a bootstrap method. Namely, one of the binary sequences from each pair was taken as a template, and the other was used to generate surrogate data by randomization of real interspike intervals. The correlation between the cells was considered significant when the mutual information computed between the “real” pair of cells was higher than that computed from the set of 500 surrogate pairs (statistical threshold, 5%).
Biocytin filling and histochemistry. Biocytin (5 mg/ml; Sigma) was dissolved in the pipette solution, and cells were filled during at least 45 min of recording (performed at room temperature). Subsequently, slices were fixed overnight in 2% paraformaldehyde at 4°C. Biocytin-filled cells were visualized using the avidin-HRP reaction (ABC Elite peroxidase kit; Vector Laboratories, Burlingame, CA) according to the instructions of the manufacturer.
Results
SNc DA neurons (n = 287) recorded from coronal rat brain slices were distinguished from fast-spiking neurons of the SNc and GABAergic output neurons of the substantia nigra pars reticulata by their specific electrophysiological features (Grace and Bunney, 1983a; Kita et al., 1986). Indeed, DA neurons display characteristic passive membrane properties and spiking pattern (Fig. 1A,B), including a pronounced sag (Ih current activation) (-21.1 ± 2.0 mV) during hyperpolarization steps (-90 pA) followed by a depolarizing rebound (11.3 ± 1.0 mV) at the stimulus offset, a depolarized action potential threshold (-36.4 ± 0.4 mV), a long spike duration (5.6 ± 0.1 msec), a large fast afterhyperpolarization amplitude (18.7 ± 0.8 mV), and finally a slow spontaneous regular spiking activity (3.7 ± 0.4 Hz) (as measured from 39 DA neurons).
Tracer coupling between dopaminergic neurons
Tracer coupling between DA neurons was assessed within the first postnatal month (P5-P25). Electrophysiologically identified DA neurons (n = 71) were successfully filled with biocytin contained in the patch pipette (Fig. 1C,D). From P5 to P10, coupling incidence was 40% (n = 20), with a mean of 2.8 ± 1.4 neurons to which tracer spreads per injected cell, but tracer coupling was no longer observed between P10 and P15 (n = 27). Surprisingly, between P15 and P25, 17% of DA cells (n = 24) were found coupled with one to three surrounding neurons (Fig. 1D). Thus, tracer coupling appears over two separate periods, suggesting a developmental regulation (variations of tracer coupling were found to be significant assessed by Fischer's exact test). Neither somato-somatic tracer coupling between DA neurons nor neuroglial coupling was detected.
Electrical coupling between dopaminergic neurons
The existence of electrical synapses between DA neurons was investigated by dual whole-cell patch-clamp recordings (at 32°C) from 66 DA neuron pairs during time periods when tracer coupling was observed.
In P7-P10 animals (n = 51 DA neuron pairs), most of DA neuron pairs were electrically coupled. Indeed, during injection of current or voltage in the presynaptic cell (stimulated cell), an electrotonic response was detected in the postsynaptic cell (receiving cell) in 96% of DA neuron pairs (Fig. 2). It is worth noting that membrane potential rebounds after hyperpolarizing or depolarizing steps were efficiently electrotonically transmitted (Fig. 2). The mean value of the coupling coefficient (k) of electrically coupled DA neuron pairs (n = 49) was 2.3 ± 0.2% (ranging from 1.4 to 4.3%) corresponding to a junctional conductance value (Gj) of 86.2 ± 7.0 pS (from 22.8 to 170.8 pS). Attempts were made to increase k by reducing K+ conductances (intracellular K+ replaced by Cs+) and Ca2+ buffering (10 mm intracellular EGTA added). These conditions (see Fig. 4, ○) did not significantly change the mean k (2.1 ± 0.3%; n = 4) and Gj (79.2 ± 25.1 pS; n = 4) when compared with control conditions (see Fig. 4, •). In addition, input resistance (Rinput) was not significantly different (307.6 ± 36.6 MΩ, n = 8, vs 298.6 ± 2.3 MΩ, n = 46, in control condition), denoting a weak contribution of potassium conductances in these cells.

Electrical coupling between dopaminergic neuron pairs. Simultaneous patch-clamp recordings from DA neuron pairs (V1 and I1 referring to cell 1; V2 and I2 referring to cell 2). Junctional currents in both directions were observed and estimated after injection of depolarizing or hyperpolarizing currents or applying voltage steps. Left, Current-clamp mode, -200 and +100 pA. Right, Voltage-clamp mode, -80 and +40 mV. k values, calculated as the ratio of the membrane potential variations in the two cells, were k12 = 4.4 ± 0.5% and k21 = 2.7 ± 0.3%. These values corresponded to a Gj of 154 and 149 pS, respectively. Traces were averaged from three to six sweeps in voltage-clamp mode and 8-19 sweeps in current-clamp mode. Note that the poststimulus rebound observed in the stimulated cell was efficiently transmitted in the receiving cell and that the spike is extremely attenuated. Holding membrane potentials were -60 mV (spontaneous membrane potentials, -40 mV).
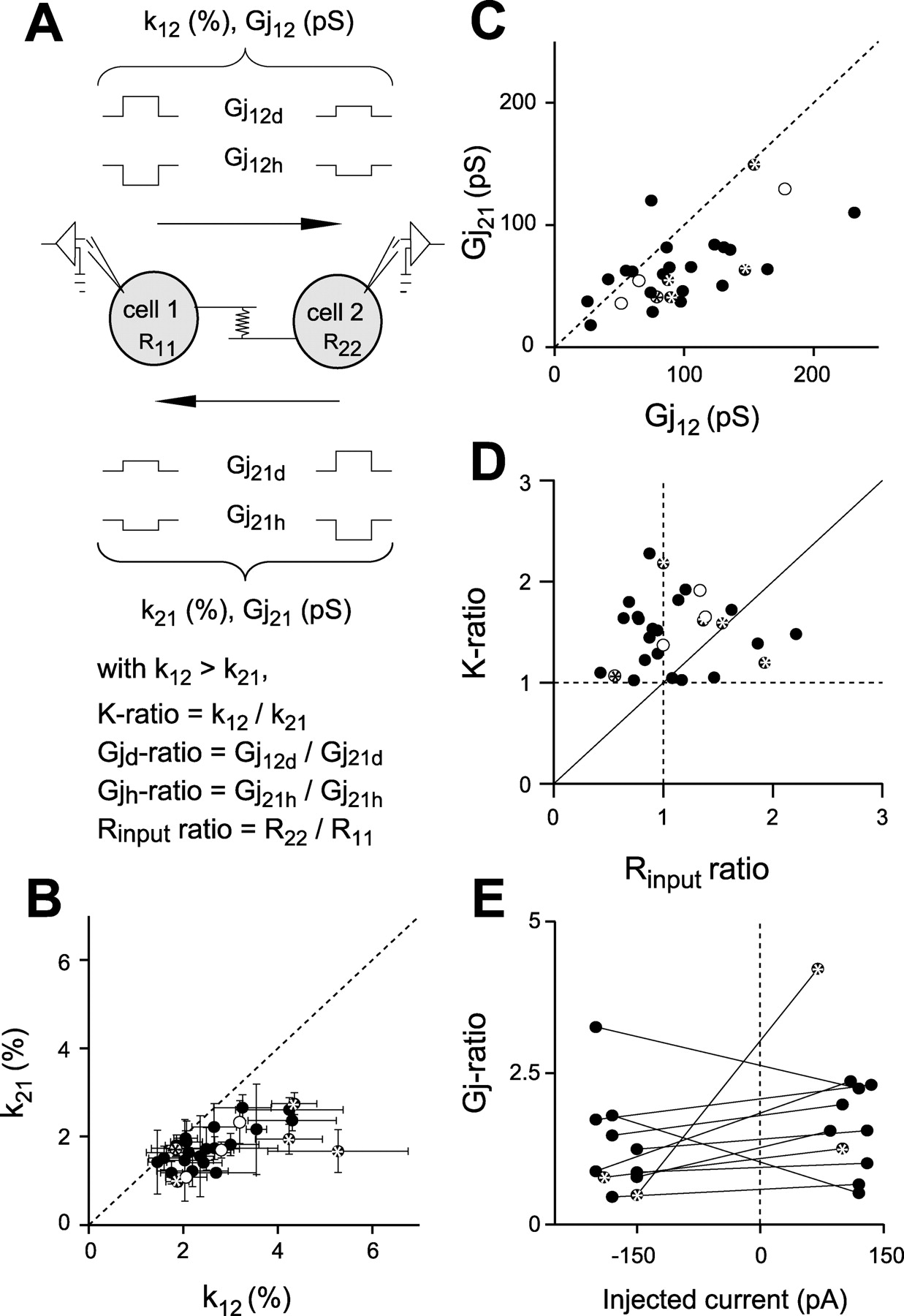
Characteristics of electrical synapses between dopaminergic neurons. A, Scheme of the protocol procedure with definition of parameters used in B-E. B, C, Comparison of k (B) and Gj (C) in both directions of coupling for each cell pair (n = 28) indicates, for most (82%) DA neuron pairs, a lack of significant rectification. k was plotted with k12 being the higher k, and Gj values were then plotted accordingly. Dashed lines indicate identical k or Gj in both directions, representing a symmetrical coupling. D, Relationship between apparent rectification (K ratio) and Rinput ratio. For each DA neuron pair, the K ratio is the ratio of the higher k to lower k (i.e., K ratio = k12/k21). The corresponding Rinput ratio is the ratio between the Rinput in the postsynaptic DA neuron versus the Rinput of the presynaptic cell in the direction of the larger k (i.e., Rinput ratio = R22/R11). The continuous diagonal line represents values expected for cell pairs connected by symmetrical Gj (with asymmetrical k and Rinput). Vertical and horizontal dashed lines represent values expected for cell pairs with Rinput and k symmetrical, respectively. The mean K ratio was 1.52 ± 0.07. Recordings with Cs+-based intracellular solution (n = 4; ○) did not differ significantly from those obtained in normal conditions (n = 24; •). E, Relationship between Gj ratio and injected current in 11 DA neuron pairs (Gjh ratios and Gjd ratios are plotted on the left and right sides of the vertical dashed line, respectively). Except for one cell pair (also displaying an asymmetrical coupling), DA electrical synapses appeared to be mainly nonrectifying. * indicates pairs presenting a significantly asymmetrical coupling.
When P17-P21 rats (n = 15 DA neuron pairs) were used, coupling incidence decreased to 20% of the recorded pairs. In these conditions, k (1.9 ± 0.3%; ranging from 1.4 to 2.2%) and Gj (60.3 ± 0.7 pS; ranging from 59.1 to 61.7 pS) values were not significantly different from those measured in P7-P10 rats.
Attempts were made to pharmacologically block gap junctional communication between DA neurons. Transjunctional currents were totally abolished in the presence of 2 mm octanol (n = 4; data not shown). However, unsaturated alcohols (heptanol and octanol) and volatile anesthetics (halothane) known to be efficient uncoupling agents also have several nonspecific effects (Venance et al., 1998; Rozental et al., 2001). Consequently, a rather specific uncoupling agent, carbenoxolone (Davidson et al., 1986) (but see Rouach et al., 2003, for inhibitory effect on spontaneous activity) was chosen. Carbenoxolone (200 μm) totally abolished junctional current in three of four cell pairs (Fig. 3). No significant change in Rinput between control and carbenoxolone conditions was observed. Indeed, the average value of input conductances (3723.2 ± 0.2 pS) was much larger than the junctional conductance values (86.2 ± 7.0 pS).

Inhibition of junctional currents by carbenoxolone (200 μm). Junctional currents were totally abolished after 5 min of bath application of carbenoxolone 200 μm (V2 is the average of 15 traces). Voltage traces were obtained from coupled DA neurons (P9), both held at -60 mV. The k value before carbenoxolone application was 1.0 ± 0.1%. Note that V2 baseline in the presence of carbenoxolone appears less noisy than V2 in control conditions. This could be attributable to an unspecific effect of carbenoxolone on spontaneous synaptic activity, as reported previously by Rouach et al. (2003).
Equality of k for both directions of electrical coupling was analyzed in 28 cell pairs (Fig. 4). Most DA neuron pairs (82%) displayed equal k, whereas five DA cell pairs showed significantly different k (ranging from 1.6- to 2.2-fold) (Fig. 4B). The relative values of the two Rinput of the cells were then investigated. First, Gj was estimated, and a distribution similar to the k plot was obtained (Fig. 4C). Second, the ratio of the higher to the lower k (K ratio) was plotted against the ratio of the Rinput of the postsynaptic to the presynaptic cell in the direction of the higher k (Rinput ratio) (Fig. 4D). In their great majority, cell pairs displayed a symmetric Gj.
No evidence of rectification was found when successive hyperpolarizing and depolarizing current steps were applied. Indeed, we measured Gj for depolarizing and hyperpolarizing stimuli in each direction of coupling (Gj for depolarizing stimuli, Gjd12 and Gjd21; Gj for hyperpolarizing stimuli, Gjh12 and Gjh21) and then compared ratios of Gjd12/Gjd21 and Gjh12/Gjh21 (n = 11 electrically coupled DA neuron pairs) (Fig. 4E). The weak difference between Gjd ratio and Gjh ratio indicated that the investigated electrical synapses were mainly nonrectifying.
Electrical synapses between dopaminergic neurons act as a low-pass filter
Brain electrical synapses described thus far display low-pass filter properties (Galarreta and Hestrin, 2001). To determine whether electrical synapses between DA neurons exhibit such a characteristic, sinusoidal stimuli of different frequencies (1-50 Hz) were applied to the presynaptic neuron (Fig. 5A-C). Electrically coupled DA neuron pairs (P7-P10; n = 30) exhibited low-pass filtering properties, namely a strong attenuation (3.0-fold between 2 and 10 Hz) in the k value and an increasing phase lag (3.1-fold between 2 and 10 Hz) for higher frequencies. Transmission of spikes through gap junctions appears to be a physiological illustration of these low-pass filtering properties. Indeed, in electrically coupled DA neuron pairs, presynaptic action potentials can generate electrotonic postsynaptic events, named spikelets (Bennett, 1997; Galarreta and Hestrin, 2001). Such events should be highly dependent on k values, electrotonic distance of the coupling site, and neuronal architecture. Among DA neuron pairs, spikelets were only observed in an electrically coupled pair presenting one of the highest k (3.5 ± 0.8%) values (Fig. 5D). In this cell pair, spikelets displayed a mean peak amplitude value of 0.31 ± 0.02 mV (n = 14) corresponding to a k (ratio of spikelet to presynaptic spike amplitudes) of 0.52 ± 0.03%. Spikelets showed a latency (basis to basis) of 0.6 ± 0.1 msec (peak-to-peak latency, 2.6 ± 0.3 msec), a rise time of 2.1 ± 0.4 msec, and a decay time of 14.1 ± 2.1 msec (n = 14).

Electrical synapses between dopaminergic neurons act as low-pass filters. A, Subthreshold sinusoidal current injections (I1) (200 pA peak-to-peak amplitude at 2 Hz, 260 pA at 5 Hz, 400 pA at 10 Hz) into cell 1 of an electrically coupled pair induced membrane potential oscillations in both cell 1 (V1) and cell 2 (V2) (averages of 50 traces). Mean k value of this DA neuron pair was 3.5 ± 0.8%. B, Magnitude of the fast Fourier transform computed from traces shown in A, V2 (bin, 1 Hz). The major frequency in the receiving cell is identical to the applied stimulating frequency (A; I1). C, Frequency dependency of k (○) and phase lag (•). Values were normalized for each DA neuron pair to the 10 Hz k and phase lag values (n = 19). Mean k at 10 Hz was 0.20 ± 0.05%. Mean phase lag at 10 Hz was 173 ± 7°. Note that coupling coefficient decreases with increasing frequency of the sinusoidal oscillations associated with an increased phase lag. D, Spike transmission through electrical synapses between a pair of dopaminergic neurons. The attenuated transmission of presynaptic spikes (spikelets) is illustrated by superimposing the presynaptic spike over an induced spikelet recorded in the postsynaptic cell of a coupled DA neuron pair. Holding membrane potentials were -60 mV.
Electrical coupling fails to synchronize spontaneous activity but efficiently modulates firing rate of coupled dopaminergic neurons
Because most DA neurons are tonically active, both in vitro and in vivo (Grace and Bunney, 1983a; Sanghera et al., 1984; Freeman et al., 1985; Kita et al., 1986), the impact of electrical coupling on spontaneous spiking activity was investigated. Among all recorded DA neurons, 94% were found to be tonically active in our conditions. First, to determine a possible synchronization in spontaneous firing between DA neurons (membrane potential, -44.9 ± 0.8 mV when no current was injected; n = 62 neurons), cross-correlation analysis was performed on 31 P7-P10 DA neuron pairs (Fig. 6A). Summation of the 31 normalized cross-correlograms did not reveal any correlation between pairs of DA neurons (Fig. 6Aa,Ab). To objectify this result, we calculated the mutual information value in the 20 msec range of the suprathreshold activities for each pair of simultaneously recorded coupled DA neurons. Mutual information value was weak and did not significantly segregate from values calculated from surrogate data (obtained by random shuffling of interspike intervals) (Fig. 6Ac). Thus, coupled DA neuron activities did not show any significant, linear or nonlinear, functional correlation.

Spontaneous firing activities of electrically coupled dopaminergic neurons are interdependent but not synchronous. Aa, Simultaneous recordings of spontaneous firing in an electrically coupled DA neuron pair. * indicates synchronous action potentials in a 10 msec range. Ab, Summation of the normalized cross-correlograms of 31 DA neuron pairs activities (bin, 20 msec). Spike count is normalized by the total number of reference spikes. Ac, Statistical analysis of the correlation between the activities of the DA neurons shown in a using a bootstrap method (see Materials and Methods). Position of the experimental mutual information value (arrowhead) calculated from original data did not segregate from the distribution of the mutual information values calculated from 500 surrogate recordings (built by randomization of the real interspike intervals of cell 2 in the same pair). This statistical analysis indicates that there was no significant correlation between spontaneous firing of DA neurons in this pair. Ba, Hyperpolarizing and depolarizing current injections in cell 1 (I1), while no current was injected in cell 2 (I2) allowing a spontaneous tonical activity, induces a decrease and an increase in the spontaneous firing rate of cell 2 (V2), respectively. Bb, Mean change of the spontaneous firing rate of the receiving cells for hyperpolarizing pulses (-200 pA; n = 7 DA neuron pairs) and depolarizing pulses (+70 pA; n = 8 DA neuron pairs) applied in the stimulated cells.
Finally, electrical synapses between DA neurons mediated an efficient modulation of spiking activity between electrically coupled neurons (Fig. 6B). To investigate the weight of electrical synapses on spiking activity, depolarizing or hyperpolarizing pulses were applied in one cell, and the effect on the action potential frequency of the postsynaptic cell was analyzed (Fig. 6Ba). In 64% of DA neuron pairs (P7-P10; n = 11), the firing rate of the postsynaptic neuron was significantly decreased (-6.6 ± 2.0%; n = 7) during the injection of a hyperpolarizing current into the presynaptic cell. Similarly, in 73% of the tested DA neuron pairs (n = 11), a significant increase in the spontaneous firing rate of the postsynaptic cell (+5.2 ± 1.7%; n = 8) was observed when a depolarizing current was applied to the presynaptic cell (Fig. 6Bb).
Discussion
Our study demonstrates that nigrostriatal DA neurons are connected by functional electrical synapses. Spontaneously active DA neurons in coupled pairs do not display significant synchronous activity. Nevertheless, membrane potential variations of one of these coupled DA neurons efficiently modulate the firing frequency of the coupled DA neuron.
Tracer coupling between dopaminergic neurons
A pioneering in vivo study by Grace and Bunney (1983b) first reported dye coupling between DA neurons in adult rats. Our results extend this observation because tracer coupling could be demonstrated in juvenile rat brain slices. Although tracer coupling was observed in P5-P10 and P15-P25 animals, surprisingly, tracer coupling was absent between P10 and P15. Two hypotheses could explain such observation: either an absence of junctional channels linking DA neurons or junctional channels are no longer permeable to biocytin. Such lack of tracer coupling at this time could explain the absence of dye coupling reported by Lin et al. (2003) in the same neurons (besides the difference of slice angle, horizontal vs coronal).
This temporary lack of tracer coupling takes place during a critical period of DA cell maturation. Indeed, an apoptotic peak occurs between P12 and P16, interpreted as a target-dependent selection of DA neurons (Marti et al., 1997; Burke, 2003). This temporary absence of intercellular communication may have a protective effect by preventing the propagation of death signals between preapoptotic and nonapoptotic DA neurons (Lin et al., 1998; Andrade-Rozental et al., 2000).
The tracer coupling between DA neurons suggests the diffusion and propagation of small molecules, such as energy metabolites, amino-acids, and second messengers between these cells (Harris, 2001). As shown in the peripheral neuroendocrine system, second messenger diffusion through gap junctions plays a key role in signaling and synchronization (Saez et al., 2003). Therefore, metabolic coupling between DA neurons may contribute to the formation of neuronal ensembles within the DA cell population.
The modalities of a metabolic coupling within and between subpopulations of DA neurons remain to be explored. Based on their electrophysiological characteristics (Gu et al., 1992; Hajos and Greenfield, 1993; Neuhoff et al., 2002), connectivity (Fallon and Moore, 1978; Grace and Onn, 1989; Gauthier et al., 1999), receptor subtypes (Chen et al., 2001), and peptide expression (Seroogy et al., 1988), DA neurons are heterogeneous. Depending on their functional states (open or closed, respectively), gap junction channels may attenuate or unveil the heterogeneity between subpopulations of DA cells. According to results from calcium-signaling experiments, such a role of gap junction channels has already been demonstrated in subpopulations of striatal astrocytes in culture (Venance et al., 1998).
Electrical coupling between dopaminergic neurons
DA neurons are connected by functional electrical synapses. Electrical coupling had a high incidence in P7-P10 rats, decreasing thereafter but still present in 20% of the recorded DA cells in P17-P21 rats. Electrical coupling between DA neurons was bidirectional and mainly symmetrical like most other electrical synapses. Electrical synapses between DA neurons act as low-pass filters, a functional property shared also by all electrical synapses described so far in the CNS of mammals (Galarreta and Hestrin, 2001; Bennett and Zukin, 2004). Indeed, fast events such as spikes were greatly attenuated or even not detected in the postsynaptic cell, whereas events with slow kinetics such as the sag and the posthyperpolarization rebound were efficiently transmitted. In DA neurons, these low-pass filtering properties of electrical synapses were more pronounced than those described for electrical synapses of neocortical, thalamic, or retinal GABAergic interneurons (Galarreta and Hestrin, 1999; Landisman et al., 2002; Meyer et al., 2002; Veruki and Hartveit, 2002).
Functional implications
The activity of DA neurons is regulated by the nigro-striato-nigral feedback loop (Bunney and Achajanian, 1976) and, locally, by dopamine released from dendrites (Cheramy et al., 1981) through its inhibitory effect via DA autoreceptors (Groves et al., 1975; Skirboll et al., 1979; Pucak and Grace, 1994). According to our results, the depolarization of a DA cell can increase the spontaneous firing frequency of the electrically coupled neuron despite the expected hyperpolarization resulting from the action of dendritically released dopamine on DA autoreceptors. Electrical synapses thus appear to have a greater impact than chemical interactions in fast communication between DA neurons. However, determination of the respective localizations (segregation or not) of electrical synapses and chemical interactions is necessary to understand their interplay and respective weight.
DA neurons display two modes of activity (tonic and phasic) resulting in different amounts of dopamine released in the striatum (Gonon, 1988). The tonic discharge, associated with a low but constant release of dopamine, supports a permanent tune-up of the striatal network, whereas phasic increases or decreases of discharge, resulting in important peaks or abrupt reductions in the transmitter release, correspond to a predictive reward value or to the lack of expected reinforcement, respectively (Schultz et al., 1997). In slices, DA neurons display only tonic activity without glutamatergic stimulation (Kita et al., 1986; Mereu et al., 1997). According to our results, this tonic activity was not significantly synchronized in electrically coupled DA neurons. However, the involvement of electrical coupling in synchronization processes is now well established for GABAergic interneuron networks (Fricker and Miles, 2001; Galarreta and Hestrin, 2001; LeBeau et al., 2003; Bennett and Zukin, 2004). In contrast to GABAergic interneurons that transmit fast information, DA neurons have wider action potentials (6 vs 0.5 msec), display a much lower firing frequency (3-4 vs 40-70 Hz), and have a neuromodulatory role. These important kinetic and functional differences between the two types of cells could explain the apparent differences in the role of electrical coupling in these cells. Indeed, the lack of synchronization between tonically active DA neurons could be essential for a constant and nonpulsatile striatal dopamine release.
Nevertheless, according to the mathematical models of DA neuron pairs performed by Komendantov and Canavier (2002), in which NMDA receptor-mediated excitatory inputs are taken into account (mimicking an in vivo context), electrical coupling should promote synchronous bursting activity. This prediction is particularly promising because synchronization under NMDA stimulation is expected especially for weak electrical coupling (in the range of Gj estimated from our study). Thus, investigation of synchronization under NMDA-mediated stimulation remains to be performed because the expected synchronization of subsequent dopamine peaks in distinct striatal areas could have important functional consequences such as the linkage of reward-related information concerning different behavioral aspects in which the nigrostriatal pathway is involved. Still, synchronization may involve more complex processes, because an in vivo study by Morris et al. (2004) reports that only 27% of DA neurons display a synchronous activity.
Footnotes
This work was supported by an Action Concertée Incitative “Jeune Chercheur” grant from the French Ministère de la Recherche, by Fondation de France Grant 20020111943, by the Institut National de la Santé et de la Recherche Médicale, and by the College de France. We thank Dr. Laurent Pezard for help with statistical analysis of nonlinear correlation; Anne-Marie Godeheu for technical assistance for histology; and Prof. Jean-Michel Deniau, Elodie Fino, Dr. Christian Giaume, Dr. Jasna Jerecic, and Prof. Istvan Mody for careful reading of this manuscript.
Correspondence should be addressed to Laurent Venance, Laboratoire de Neurobiologie Pharmacologique, Institut National de la Santé et de la Recherche Médicale U-114, Collège de France, 11 place Marcelin Berthelot, 75005 Paris, France. E-mail: laurent.venance{at}college-de-france.fr.
Copyright © 2005 Society for Neuroscience 0270-6474/05/250291-08$15.00/0





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}