
Abstract
Research on avoidance conditioning began in the late 1930s as a way to use laboratory experiments to better understand uncontrollable fear and anxiety. Avoidance was initially conceived of as a two-factor learning process in which fear is first acquired through Pavlovian aversive conditioning (so-called fear conditioning), and then behaviors that reduce the fear aroused by the Pavlovian conditioned stimulus are reinforced through instrumental conditioning. Over the years, criticisms of both the avoidance paradigm and the two-factor fear theory arose. By the mid-1980s, avoidance had fallen out of favor as an experimental model relevant to fear and anxiety. However, recent progress in understanding the neural basis of Pavlovian conditioning has stimulated a new wave of research on avoidance. This new work has fostered new insights into contributions of not only Pavlovian and instrumental learning but also habit learning, to avoidance, and has suggested that the reinforcing event underlying the instrumental phase should be conceived in terms of cellular and molecular events in specific circuits rather than in terms of vague notions of fear reduction. In our approach, defensive reactions (freezing), actions (avoidance) and habits (habitual avoidance) are viewed as being controlled by unique circuits that operate nonconsciously in the control of behavior, and that are distinct from the circuits that give rise to conscious feelings of fear and anxiety. These refinements, we suggest, overcome older criticisms, justifying the value of the new wave of research on avoidance, and offering a fresh perspective on the clinical implications of this work.
Similar content being viewed by others

Introduction
Avoidance is a natural and adaptive response to danger. Animals, including humans, cannot survive without the ability to avoid harm. Nevertheless, avoidance can have detrimental consequences—excessive and/or unnecessary avoidance is a hallmark of anxiety disorders.1, 2, 3, 4, 5, 6, 7, 8 In order to understand this nuanced behavioral phenomenon, researchers in the late 1930s began studying avoidance conditioning in animals.9, 10, 11 The goal of this work was to illuminate the psychological processes underlying human and animal avoidance, as well as to inform ways to ameliorate the troubling consequences of pathological avoidance in people suffering from debilitating anxiety. But by the 1980s, unresolved conceptual debates and inconsistent findings about the underlying neural circuitry prompted researchers to abandon avoidance and turn to simpler Pavlovian conditioning paradigms.12, 13, 14 The result was a wealth of data about the circuits, cells, synapses and molecules underlying so-called Pavlovian fear conditioning.15, 16, 17, 18, 19, 20, 21 But many important questions have gone unanswered about the clinically important topic of avoidance. Fortunately, after several decades of neglect, there are signs of growing interest in avoidance conditioning and its neural underpinnings.22, 23, 24, 25, 26, 27, 28, 29, 30
In this review we consider the nature of avoidance conditioning and discuss why it fell out of favor as a behavioral paradigm. We argue that progress in understanding the circuitry underlying Pavlovian aversive conditioning has made it possible to revisit the neural basis of avoidance from a fresh perspective. We also argue that key criticisms that plagued the avoidance paradigm were conceptually misguided. The hypothesis that emerges is that avoidance involves three forms of learning—Pavlovian conditioning, action-outcome learning and habit learning, each mediated by a unique neural circuit. We end with a consideration of the clinical implications of the new wave of avoidance research.
What is avoidance conditioning?
Avoidance refers to both a behavioral conditioning procedure used in laboratory studies and a coping strategy used by anxious people. Most of this review will focus on the laboratory research on avoidance, with the clinical implications of that research saved for the end.
In a laboratory context, avoidance is defined as a class of conditioning procedures in which subjects learn to minimize or prevent contact with aversive events (typically electric shocks or stimuli associated with them). Under this broad heading, different forms of avoidance are recognized.25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35 The broadest distinction is between passive and active avoidance. In the passive avoidance procedure (also called inhibitory avoidance), harm is avoided by withholding responses. A rat that has been shocked when it steps off a platform or enters a certain location can avoid shock by withholding those behaviors. The primary focus of this review is active avoidance, where harm is prevented by taking action. There are several forms of active avoidance conditioning. Some involve warning signals (signaled active avoidance) and others do not (unsignaled or Sidman avoidance). Many studies have used the shuttle box signaled active avoidance procedure, in which a warning signal indicates that the subject can avoid harm by crossing a divided chamber (Figure 1). Other signaled active avoidance procedures use responses such as lever pressing or stepping onto a platform to avoid shock. In this review, the term avoidance will refer to active avoidance, except when noted.

Active avoidance: the shuttlebox learning paradigm. Top panel: initially, subjects undergo Pavlovian threat conditioning, in which a conditioned stimulus (CS; tone) is paired with an aversive unconditioned stimulus (US; shock). Middle panel: once the CS–US association is acquired, subjects learn that the US can be inactivated by shuttling—this is an escape response. On subsequent trials, subjects learn that shuttling during the CS causes the inactivation of the CS and the omission of the US—this is an avoidance response. Bottom panel: once behavior becomes well-trained, the behavior is preformed in the presence of the CS, even though the US does not result. With continued training the behavior persists habitually in spite of the fact that US is no longer predicted by the CS.
The birth of the active avoidance paradigm
John Watson36 built on Ivan Pavlov’s conditioning paradigm37 in founding the behaviorist school of psychology. In one of the earliest and most famous studies of Pavlovian conditioning, Watson presented a young boy with a neutral stimulus (a white rat) in connection with a loud noise.38 Thereafter, the presence of the rat induced crying and other signs of distress in the boy. This came to be described as ‘fear learning,’ and paved the way for future studies of so-called Pavlovian ‘fear conditioning’ in animals and humans. Despite the initial impact of Watson’s experiment, much of the research done under the banner of behaviorism focused on the other major form of behavioral learning, instrumental or operant conditioning. In the instrumental conditioning procedure, complex responses are acquired (reinforced) by the outcomes they produce.39, 40, 41 This form of learning was viewed as more relevant to complex human behaviors than Pavlovian conditioning, which involves simpler behavioral and physiological reactions. Coming out of this behaviorist tradition in the late 1930s, O.H. Mowrer9, 10, 11 merged Pavlovian and instrumental approaches by choosing conditioned avoidance for his animal studies of aversion, which he pursued to elucidate the mechanisms of fear and anxiety in humans.
Mowrer accepted on Sigmund Freud’s42 premise that fear and anxiety are learned states, but he recast Freud’s ideas in terms of behaviorist stimulus–response principles. According to Mowrer,9 ‘anxiety (fear) is the conditioned form of the pain reaction, which has the highly useful function of motivating and reinforcing behavior that tends to avoid or prevent the recurrence of the pain-producing (unconditioned) stimulus.’ Mowrer thought that the Pavlovian conditioned stimulus (CS) served to trigger a fearful state because of its learned association with the aversive unconditioned stimulus (US). If a behavior allowed the subject to remove or minimize exposure to the CS and/or to prevent contact with the US, it was thought to alleviate CS-elicited fear. Thus, Pavlovian conditioning established a CS–fear link, and then through instrumental conditioning a fear-avoidance response link occurs. Behaviors that successfully reduce fear were said to be reinforced and learned for use in the future.43, 44, 45, 46, 47, 48, 49
Conditioned fear reduction was considered the essential psychological mechanism that reinforced instrumental avoidance behavior.10, 11, 45, 46, 50, 51 In this two-factor account of the common avoidance experiment depicted in Figure 1, the rat learns to cross the divided chamber when doing so causes cessation of a tone CS and omission of a shock US. This action, according to two-factor theory, allows the subject to mitigate the fear state triggered by CS. The two-factor theory of avoidance is thus often referred to as two-factor ‘fear theory’.47, 52
Mowrer’s ideas and research,10, 11, 46, 51, 53 together with the work of his colleague Neal Miller,45, 50, 54 defined laboratory studies of fear and anxiety for the next several decades. A large number of studies were conducted on avoidance behavior6, 31, 43, 44, 49, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66 and the underlying brain mechanisms.67, 68, 69, 70, 71 Because avoidance was also recognized as a major symptom of pathological fear and anxiety in humans,72 the Mowrer–Miller approach greatly influenced subsequent ideas about the nature and treatment of these disorders.1, 5, 6, 52, 63, 72, 73, 74, 75 However, despite the broad impact of the avoidance paradigm, with few exceptions,76 laboratory research on active avoidance had largely ceased by the mid-1980s.
The death of avoidance
The demise of avoidance research was the result of a number of issues with the paradigm that accumulated between the 1940s and 1980s.22, 23, 24 Particularly troubling were unresolved conceptual debates about the psychological processes underlying avoidance behavior. The arguments carried on for years without coming to a satisfying resolution,43, 44, 47, 49, 52, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66 causing researchers to question the fundamental value of the paradigm.60, 62 The two most contentious questions were the following: (1) whether fear reduction reinforces the avoidance response; and (2) whether learned avoidance responses qualify as instrumental behaviors under the criteria of learning theory. These questions will be addressed in detail in the next section. For now we consider other issues that had a negative impact on avoidance research.
Interest in the paradigm was also diminished by the fact that no clear picture of the essential neural circuits had emerged, despite years of research.67, 68, 69, 70, 71, 77 This was in part due to the poor understanding of neuroanatomical connections in the period between the birth and demise of avoidance. In addition, there was less awareness of how different avoidance tasks might engage different brain circuits. Later research on learning and memory revealed the strong impact of task structure on the underlying neural circuitry.78
Another contributing factor was the success of simple learning procedures used to identify the cellular and molecular substrates of aversive memory in invertebrates.79, 80 This, together with the failure to discover clear neural substrates of avoidance, inspired vertebrate researchers to streamline their approach by adopting simpler experimental procedures, such as Pavlovian conditioning.13, 81, 82, 83, 84 A bevy of easy-to-use paradigms for the neuroscientific study of aversive learning and memory were available from behavioral research on Pavlovian conditioning in rodent work that had proceeded in parallel with avoidance research.48, 85, 86, 87 Because Pavlovian conditioning is simpler, and thus easier to relate to its brain substrates, a disincentive arose for the use of the more complex and conceptually troubled avoidance paradigm. The net effect was that the field turned to Pavlovian procedures to study aversive conditioning. Over the subsequent decades, Pavlovian aversive conditioning, so-called ‘fear conditioning,’ emerged as one of the most successful behavioral paradigms for understanding the brain mechanisms of behavior (see discussion of this research below).
Parenthetically, the success of Pavlovian approaches was greatly aided by newly emerging tools that allowed researchers to accurately trace anatomical connections, as well as to uncover the neurochemical constituents of neurons and their synapses.88, 89 Although these methodological developments would surely have aided avoidance research to some extent as well, it is unlikely that avoidance could have succeeded without resolution of the conceptual issues that plagued the paradigm. But, as we will illustrate below, these issues may not be as troubling as they once seemed, especially when they are reconsidered in the light of contemporary ideas about learning and behavior.
Digging deeper into the conceptual stumbling blocks
Of the issues mentioned above, most damaging to avoidance research were conceptual problems surrounding the psychological processes underlying the avoidance paradigm. In particular, questions about whether fear reduction reinforces avoidance, and whether avoidance responses are instrumental (that is, learned by their consequences), remained contentious and unresolved, muting interest in avoidance conditioning as a behavioral tool.
Fear reduction as the reinforcer in avoidance
A reinforcer is a stimulus that strengthens behavior, increasing the likelihood that a given behavior will be repeated in similar situations. Negative reinforcement occurs when the elimination of a stimulus makes a behavior more likely to occur again. In the Mowrer–Miller theory, avoidance behavior was said to be negatively reinforced by CS termination because this was hypothesized to reduce the fear state elicited by the CS.45, 46, 47, 50, 51, 53, 54
Despite the early prominence of two-factor fear theory, subsequent empirical work failed to support a role for fear reduction in the avoidance paradigm.49, 57, 59, 60 One particularly damaging argument against two-factor theory had to do with extinction. Because the avoidance response prevents the US from occurring, presentations of the CS are no longer followed by the US if avoidance behavior occurs on every trial. Put differently, every trial is an extinction trial once avoidance behavior is fully acquired. This should extinguish fear elicited by the CS in and of itself, diminishing the motivation to perform the avoidance response. Yet avoidance persists and is quite resistant to extinction,6, 57, 85, 90 even though other conditioned responses thought to indicate learned fear (freezing and heart rate)83, 86, 91 can be effectively extinguished by withholding an aversive US.6, 85, 92, 93 Similarly, the behavioral and physiological reactions thought to indicate conditioned fear do not co-vary with the acquisition or performance of the avoidance response.6, 60 Such observations created a paradox for two-factor fear theory. Authors such as Bolles43, 60 and Seligman62 viewed these observations as particularly damaging to the avoidance paradigm. Two-factor theory fell out of favor in large part due to this disconnect between avoidance behavior and other observable phenomena thought to indicate conditioned fear.
Mowrer's two-factor theory was doomed to controversy by the use of a subjective state (fear) to explain learned changes in animal behavior. Because the relationship between a conscious feeling and a behavior is ultimately unverifiable in rodent subjects, the central tenant of two-factor fear theory cannot be tested in a satisfactory way in animal studies. Even if objective responses thought to indicate conditioned fear in rats did vary with avoidance, the connection between those indicators and a conscious feeling of fear could not be conclusively established. In fact, in humans, behavioral and physiological responses are poorly correlated with subjective states.7, 94, 95 If feelings of fear do not correlate with other indicators in humans, why should they in rats? Fear theory was predestined to wind up in an intractable debate, and its early prominence painted avoidance research into a corner.
To solve this problem, some offered an alternative two-factor model, called aversion theory, that did not posit fear reduction as the reinforcer.55, 56 Instead, it simply gave a behaviorist explanation—the Pavlovian CS is aversive, defined by the observable fact that animals will work to remove it. Thus, CS inactivation negatively reinforces avoidance behavior. But the field was committed to some kind of fear-based explanation. The result was an effort to operationally redefine fear as something other than a subjective state, focusing instead on observable stimuli and responses.39, 40, 41 Fear came to be a psychological or physiological intervening variable that accounted for relation between external and defensive behavior.40, 43, 47, 48, 61, 63, 91, 96, 97, 98, 99, 100, 101, 102, 103, 104, 105
Despite such attempts at reformulation, these operational definitions of fear were not used consistently. For some, fear remained a conscious feeling. Mowrer,10 for example, called for the return of consciousness to studies of animal behavior, arguing that rats freeze ‘by-cause of fear.’ Contemporary researchers such as Panksepp106, 107 argue that the conscious feeling of fear arises from the same circuits that control defensive behavior in both rats and humans. In this view, as in Mowrer’s, freezing in rats should tell us about fearful feelings in people. However, even those who tried to define fear without reference to subjective states would use the term in more than one way. Bolles,97 for example, called for care in the use of subjective state terms like fear because of the surplus meaning they possess, but also wrote about ‘frightened rats.’ Bolles’ student, and prominent ‘fear’ researcher, Michael Fanselow says that a goal of science should be to ‘replace inaccurate subjective explanations… with more scientifically grounded explanations’.104 Yet, he and other "fear" reserachers claim that freezing rats can help understand pathological fear (presumably subjectively expereinced fear) in humans.20, 21, 102, 103, 105, 108, 109 Clearly, confusingly mixed meanings of the term fear were, and still are, in common use.
In an insightful commentary on operational definitions and their monikers, Marx notes that it is important to distinguish operational validity, which concerns the empirical relation between observable variables, and semantic validity, which refers to the relation of the name applied to the operational definition.110 He points out that there is a ‘semantic danger’ that results when ‘names chosen to represent the intervening variable have …vague and varied meanings.’ When common language terms are redefined in novel ways, the result, according to Marx, is that it is not always clear, which meaning is in play because ‘each reader tends to read into the word…his own meanings and biases’. Attempts to redefine fear in nonsubjective terms suffer from this problem. For instance, the common meaning of the word fear (for example, a subjective sense of being in danger) is often conflated with the empirical basis of the operational definition (defensive responses elicited by threatening stimuli). As a result, for some, empirically observed behavioral outcomes are treated as an index of subjective feelings of fear, while for others fear is nothing more than activity in the circuit that connects threats to defense responses. As Marx’s analysis predicted, the situation has lead to confusion.
Our position is that subjective states of fear should not be invoked to describe the defensive behavior of species in which such states cannot be verified by verbal report.30, 111, 112 While we do not deny the possibility of such states in nonhuman animals, we argue that they cannot be directly assessed in nonverbal species.30 At the same time, just as problems arise from calling upon subjective fear in animals, so do they arise from failing to acknowledge the role of such states in humans,104 a species in which they can be verified and studied. Subjective experiences of fear and anxiety are a leading factor that causes people to seek clinical help, and therapies are judged successful largely based on their capacity to change these subjective experiences. This is probably why many researchers, even those who deny that subjective states have value as a scientific topic,104 feel compelled to refer to nonsubjective meditational states as states of fear or anxiety, and claim relevance to human fear and anxiety.20, 21, 102, 103, 105, 108, 109
In order to discuss the processes underlying avoidance without falling prey to the conceptual issues associated with two-factor fear theory, a change in terminology is required. As Bolles once noted, subjective state terms from human experience will always carry surplus meaning.97 To minimize subjective surplus meaning, we propose that what was once considered a fear stimulus can be referred to as a threat, which we conceive of as a potential source of danger or a danger-predictive cue. Fear conditioning, by the same token, simply becomes Pavlovian threat conditioning (PTC).112 A fear response becomes a defensive response, which we define as the behavioral outputs that protect against danger and physiological responses that support these responses. Given that numerous investigators use fear response and defense response interchangeably, this should be non-controversial. We can study how threats control these defensive responses (including avoidance) in animals and humans alike, without conflating defensive processes with the mechanisms of conscious fear experiences.30, 111, 112, 113 These terminological changes allow us to consider the underlying mechanisms in humans and animals on a level conceptual playing field. If subjective fear does not account for expression of CS-elicited Pavlovian defense responses, reduction of CS-elicited subjective fear cannot be the explanation for the why avoidance is reinforced and learned, or why, once learned, the avoidance responses are performed.
From a neuroscientific perspective, behavioral learning can be accounted for in terms of cellular and molecular events occurring in functional circuits. These events strengthen connections between stimuli in Pavlovian conditioning, and between actions and outcomes in instrumental learning. There is no need to introduce a fear construct. This point is highlighted by the observation that human feelings of fear are poorly correlated with physiological responses that are thought to measure fearful feelings,7, 94, 95 and by findings in healthy participants114, 115, 116, 117, 118, 119, 120 and blindsight124, 125, 126, 127 patients showing that threats elicit defensive responses without the person knowing the stimulus is present and without feeling fear. Moreover, amygdala damage in humans disrupts physiological and behavioral responses but not conscious feelings.121, 122, 123 These and related observations lead to the conclusion that different circuits underlie fearful feelings and defensive responses in humans.30, 128
The term fear will always carry extra meaning when used to name a nonsubjective state that functions as mediator between threats defensive responses. This is true whether we are discussing humans or other animals. Nonconscious fear, in short, is a cumbersome and misleading concept. The mediator between threats and defensive responses is a defensive circuit that involves the amydgala and related brain regions. Although the activity in this circuit does not itself give rise to a conscious fear state, it nevertheless contributes indirectly to the conscious experience of fearful feelings, which, we propose, are products of cortical systems.30 We therefore prefer terms such as ‘defensive motivational circuit’ or ‘defensive system’30, 49, 111, 112, 129, 130 that are less intrinsically biased toward subjective interpretations by their common language meaning.
In summary, conceptual problems with fear reduction can be circumvented by acknowledging that important defensive responses can be accounted for without making reference to conscious experience. In lieu of fear reduction, we take a brain-based approach to defensive behaviors evoked by conditioned threats. Though we have been critical of two-factor fear theory, we do agree that avoidance behavior involves distinct learning processes that occur in sequence. As described below, we propse three learning processes. First, the subject acquires an association between a CS and an aversive US through Pavlovian threat conditioning. Next, we believe that a negative reinforcement process guides the instrumental acquisition of the avoidance response. Third, we argue that the avoidance behavior becomes habitual with significant training, allowing it to persist even when it becomes disconnected from its reinforcing consequences.
Instrumentality of avoidance
An instrumental action is shaped by the outcomes it produces.131 As noted above, early researchers assumed that avoidance was acquired through instrumental conditioning, but this idea was strongly criticized. Here we re-evaluate the arguments against the role of instrumental learning in the avoidance paradigm. We contend that many of these critiques were flawed in unappreciated ways and should no longer impede contemporary research on avoidance learning.
One criticism of the instrumentality of avoidance was actually a problem with two-factor fear theory itself. The influence of the Mowrer–Miller approach caused fear reduction to become conflated with an instrumental interpretation of avoidance behavior. Much of the field concluded that avoidance could not be considered instrumental without a fear-reduction reinforcement mechanism. Once two-factor theory fell out of favor, the instrumentality of avoidance was also dismissed. An obvious alternative was not given substantial consideration—that the avoidance response is instrumental even though fear alleviation is not the relevant reinforcer. The failure of two-factor fear theory says more about the field’s view of fear than about the role of instrumental processes in avoidance behavior.
Another issue revolved around a key control condition, the yoked control, in avoidance research. In this procedure, subjects are paired such that one animal determines the delivery of stimuli for both, creating an arrangement in which both subjects receive the same temporal pattern of experimental events even though only one has control. This design was a major tool used to establish the instrumentality of a behavior. However, the yoked control has been forcefully criticized. In particular, Church argued that within and between subject variation can systematically bias the paradigm, leading to differences in group behavior that are not necessarily due to instrumental control.132, 133 The ambiguities of the yoked design, which had been in common use, made it methodologically difficult to establish the instrumentality of avoidance. This methodological problem added impediments to the question of how to study instrumentality, but this is not the same as evidence against instrumentality.
There were other criticisms, as well. Bolles, for example, argued that avoidance behavior was a CS-controlled, species-specific defensive response.59, 60, 91 Specifically, avoidance was defined as CS-elicited ‘flight’, acquired through Pavlovian learning. Because flight responses are not modifiable by the consequences they produce, Bolles concluded that avoidance was not instrumental. However, there are fundamental differences between the classic flight response and avoidance. Flight is typically a stereotyped, innate burst of activity in reaction to imminent danger. An avoidance behavior is a complex action that can take a number of forms, which vary substantially between individuals and situations. Often the avoidance responses in a shuttle box more resembles locomotion134 or ‘active wandering’135 than the rapid burst of activity characteristic of innate flight that Bolles seems to refer to. Moreover, avoidance responses are acquired and maintained in ways that make them distinct from Pavlovian responses. Relative to Pavlovian freezing, avoidance responses are more slowly acquired, more difficult to extinguish, more variable and are mediated by different brain circuits (see below).
Another important point that is not widely recognized is that key findings in the classic literature on avoidance involved data collected from animals that had undergone extensive training.57, 85, 90 Modern learning theory recognizes that well-trained responses can become habits, which are actions that continue despite a weakened connection to the reinforcing outcome by which they were first acquired.131, 136, 137, 138, 139, 140 This is critical, because a behavior’s instrumentality (that is, outcome dependency) cannot be assessed once that behavior has become habitual (that is, outcome independent). The focus on well-learned habitual responses may account for at least part of why past research was unable to establish a role for instrumental conditioning in avoidance.43, 48, 49, 59, 60
The concept of habit also resolves the extinction paradox described in the previous section. Briefly, if a subject avoids on every trial, it no longer has contact with the aversive US, and the avoidance response should extinguish. Critics wondered why the response persisted in such a situation. Habit provides a conceptual resolution to this problem. Contact with the reinforcing stimulus is not required once a behavior has become habitual.
Habit also sheds light on the inconsistent results of early neuroscientific studies of avoidance.68, 69, 70, 71, 76 Appetitive studies have shown that the transition from instrumental action to habit involves a shift in the underlying neural circuitry.136, 139, 140 Consistent with the appetitive work, the amygdala is required for the acquisition and expression of the avoidance response,141 but not for avoidance after extensive training.142 Amygdala manipulations carried out at different time points thus produce inconsistent results, which were difficult to interpret without acknowledging an important third factor in avoidance learning.
In short, the idea of habit is an important solution to more than one issue with the avoidance paradigm. But why does avoidance become habitual? Habit is a form of ‘automatic’ behavior that can be acquired with substantial experience. The advantage of automaticity is that it allows the brain to bypass the extensive neural circuitry needed to process environmental contingencies that have already been established. Instead, a streamlined circuit directly connects a stimulus to a response. While this fosters efficient processing, the cost is a reduced sensitivity to changes in outcome. Future research on avoidance should explore the hypothesis that habit is a crucial third factor in avoidance learning.
The arguments against instrumentality have long stigmatized avoidance as being too problematic to pursue. While flaws in key critiques do not establish the instrumentality of the avoidance response, we believe that our re-evaluation of the literature justifies a fresh look at the avoidance paradigm. Moving forward, studies designed to assess the psychological structure of avoidance conditioning will benefit from advances in learning concepts that arose after avoidance work fell out of favor. For example, the notion that actions can become habits is an insight from learning theory that has already paid off. Further, nuanced criteria have emerged to determine if a behavior is instrumental,136, 137, 143, 144, 145, 146, 147, 148, 149, 150 providing clearer guidelines for pursing the instrumentality of avoidance as the new wave of research proceeds.
Reinforcement of avoidance
While more work is needed to establish the instrumentality of avoidance, we can identify possible reinforcers of avoidance behavior. Some potential negative reinforcers include omission of the US (for example, prevention of a shock), escape from the CS (for example, termination of a tone) or escape from the US (for example, omission of a shock). It can be difficult to distinguish between these possibilities using an active avoidance task.60, 151
The escape from threat (EFT) paradigm, often called escape from fear,48, 151, 152 was designed to shed light on potential mechanisms of negative reinforcement. In EFT, rats first undergo Pavlovian conditioning to a tone CS and a shock US. The next day they are placed in a novel chamber where the tone CS is presented. Over trials, subjects will learn to make a specific response, such as shuttling in a runway, in order to inactivate the CS. This response is reinforced completely by tone offset, as the US never occurs in the novel chamber. EFT demonstrates that CS escape is sufficient to reinforce instrumental learning. Although early EFT paradigms were subject to a number of criticisms, many of these were addressed in more recent work151 and it seems clear that CS termination can act as a negative reinforcer.
Because EFT learning is somewhat weak, avoidance may depend on both escape from the CS and prevention of the US. Indeed, evidence suggests that both CS inactivation and US omission act synergistically to reinforce avoidance responses.58 Thus, active avoidance behavior is likely negatively reinforced by multiple salient outcomes, each of which contribute to the acquisition and performance of the response. Studies of the brain mechanisms of negative reinforcement should consider these possibilities.
Another possibility is that avoidance is reinforced, not by the removal of danger, but by the addition of safety cues. Stimuli associated with successful avoidance, such as the offset of the CS, may function as conditioned inhibitors—signals that are associated with the absence of shock.33, 153, 154 In this sense, avoidance is positively reinforced by the presence of safety cues. An elaboration of this argument suggests that the avoidance response itself functions as a conditioned inhibitor.155 Because the shock is absent when the CS and the avoidance response occur together, the response becomes a signal that discriminates between trials on which the US will and will not occur. Thus, the action itself becomes associated with the absence of shock via Pavlovian learning, and this association inhibits the usual responses (for example, freezing) evoked by the CS. While this is an intriguing idea, we note that freezing and other Pavlovian responses remain suppressed when the CS is presented in an alternate environment that does not allow the avoidance response.85, 156 In other words, avoidance training can attenuate conditioned aversive behaviors elicited by the CS even when the response is not present.
There is an important sense in which avoidance of danger and approach to safety are necessarily entwined—if safety is the absence of danger, avoidance is a source of safety. Thus, we believe that negative reinforcement resulting from removing threat and positive reinforcement from achieving safety are complimentary processes that may both contribute to avoidance learning, though perhaps in differing degrees.
Mowrer10 talked about negative reinforcement in terms of fear reduction, leading to ‘relief,’ and he conceived of approaching safety as the experience of ‘hope.’ In our view, reinforcement in avoidance, whether negative reinforcement from the reduction or removal of threats or positive reinforcement from the presence of safety signals, involves cellular and molecular processes in the functional circuitry of avoidance. These events provide the necessary and sufficient reinforcement mechanism to account for behavioral learning. Subjective experiences of fear, hope, relief or other emotions may occur but are not the cause of learning.
The resurrection: revisiting avoidance action circuitry through the lens of the Pavlovian reaction circuitry
In the previous two sections, we argued for the flaws in the major conceptual stumbling blocks that have hampered avoidance research. As a result, the renewed interest in the avoidance paradigm and its neural underpinnings is justified. In pursuing the neural basis of avoidance, we build on the argument mounted above—that avoidance involves three distinct types of learning, each supporting different behaviors that are acquired sequentially. Initially, Pavlovian conditioning results in defensive reactions, which are stereotyped species-specific behaviors (freezing, for example). These are supplanted by defensive actions (avoidance responses), which are a more flexible class of behavior that, we hypothesize, are learned by their outcomes. Finally, with additional training actions transition into defensive habits, which are highly persistent and outcome insensitive actions (habitual avoidance responses). Below we will demonstrate that these three classes of behavioral learning depend on dissociable neural substrates.
Because Pavlovian conditioning is the first of the three phases, we argue that the wealth of data about the neural circuitry of PTC can be leveraged to more effectively pursue the neural basis of avoidance learning, especially if the avoidance paradigm chosen uses CSs and USs that have been used in the Pavlovian work. For this reason, we focus on a signaled active avoidance paradigm that uses a tone CS and a footshock US. This allows us to ask whether the amygdala-based circuitry underlying Pavlovian defensive reactions overlaps with or diverges from the circuitry required for avoidance. If the circuitry of reaction and action overlap entirely, avoidance is likely to be a wholly Pavlovian process. If the action circuitry diverges from the reaction circuitry, it would suggest that reactions and actions are acquired through at least partially distinct processes. In addition, the substrates of defensive actions can be compared with the well-characterized substrates of appetitive instrumental action, which may inform the form of learning that supports active avoidance behavior. The relationship between action and habit can be evaluated similarly.
Neural circuits underlying Pavlovian threat conditioning
As noted above, PTC occurs when a neutral CS (for example, tone) is paired with an aversive US (for example, footshock) (Figure 2). The amygdala is a critical substrate of PTC in both humans and animals.16, 157, 158, 159, 160 Rodent studies demonstrate that the CS and US converge in the lateral nucleus of the amygdala (LA).161, 162, 163 During conditioning, LA neurons exhibit increases in neuronal activity evoked by the CS.164, 165, 166, 167, 168, 169 This activity is necessary to support learning—destruction or inactivation of LA disrupts both the acquisition and expression of Pavlovian reactions.170, 171, 172 Within LA a variety of cellular and molecular events transform a neutral stimulus it into an aversive CS.16, 18, 160, 173, 174, 175

Auditory pavlovian threat conditioning. Top panel: an auditory conditioned stimulus (CS) is paired with a foot shock unconditioned stimulus (US). Middle panel: when the CS is presented in a novel context, it elicits a conditioned reaction, freezing. Bottom panel: if the CS is not paired with the US it does not elicit freezing during the test.
LA projects to a variety of other nuclei within the amygdala.176, 177, 178 Of critical importance for PTC is the progression of information from LA to the central nucleus of the amygdala (CeA). Similar to LA, CeA contributes to the acquisition of PTC,179, 180, 181, 182, 183 which depends in part on the potentiation of LA synapses in the lateral subdivision of CeA.184 However, CeA is best known as a major amygdalar output nucleus that controls the behavioral, autonomic and endocrine reactions elicited by the CS.83, 185, 186 Projections from CeA to the periaqueductal gray (PAG) are necessary for CS-evoked freezing,185, 187 while projections to other hypothalamic and brainstem targets control autonomic and neuroendocrine reactions to the CS.83, 185
It is important to note that the connections between LA and CeA are both direct and indirect. The indirect connections involve LA projections to basal amygdala (BA), medial amygdala and the intercalated nuclei—each of which projects to CeA.176, 177, 178 Information about conditioned threats may travel through one or more of these pathways in order to engage the appropriate conditioned defensive reaction.
While the human amygdala cannot be explored in such detail, studies of patient populations, as well as brain imagining studies of healthy participants, confirm the basic findings of animal research. Damage to the human amygdala disrupts PTC,188, 189 and PTC elicits BOLD activity in the amygdala of healthy participants.190, 191, 192 Recent studies using depth electrodes support a role for the human LA in the rapid processing of aversive stimuli.118, 193 The human amygdala supports implicit or nonconscious forms of threat processing,30, 100, 112, 113, 114, 115, 116, 117, 118, 119, 120, 123 which can be assessed explored similarly in humans and other mammals. The rodent amygdala is an apt model for nonconscious amygdala-dependent threat processing in humans.
Neural circuits of avoidance: actions and habits
For reasons articulated above, we emphasize signaled active avoidance studies using a two-way shuttlebox task in which animals learn to avoid a US by crossing a divided chamber when a warning signal (the Pavlovian CS) is present (Figure 1). Much of the historical work on avoidance has used this approach, and the underlying circuits that have been discovered in recent work are illustrated in Figure 3. Where appropriate, we also describe studies that have used avoidance responses such as lever-pressing27, 32 or stepping onto a platform29, 35 to avoid shock in the presence of a CS.

Neural circuits underlying defensive reactions (freezing) and actions (avoidance). The behavioral illustrations show the performance of previously acquired reactions (freezing) and actions (avoidance). (a) Reactive freezing is underpinned by a progression of information through the amygdala. Information about the auditory conditioned stimulus (CS) arrives in the lateral amygdala (LA) from auditory thalamus and/or cortex. CS information then proceeds to the central amygdala (CeA), either directly through LA projections to the central lateral CeA (CL), or indirectly via the basal amygdala (BA) and/or the intercalated cell masses (ITC). Medial CeA (M) projections to the brainstem coordinate CS-evoked reactions, such as freezing. (b) Active avoidance is underpinned by a different amygdalar output pathway. CS information is processed through LA and BA, before progressing to the nucleus accumbens (NAcc), which supports CS-prompted actions, such as shuttling to avoid. This behavior is regulated by the infralimbic prefrontal cortex (PFCIL), which suppresses CeA-mediated freezing.
Early neuroscientific studies of avoidance produced confusing results.68, 69, 70, 71, 76, 77 Damage to a given brain area, such as the amygdala, sometimes disrupted avoidance, sometimes facilitated avoidance, and sometimes had no effect at all. Much of this work involved ‘whole amygdala’ lesions that spared significant tissue in some cases, while there was significant damage to extra-amygdala regions in others. Contemporary research methods allow for the targeted manipulation of distinct amygdala subnuclei, such as LA, BA and CeA.
Lesion experiments have implicated LA in the acquisition and expression of avoidance responses.141, 142, 152 The involvement of this is area is especially significant because it suggests that a CS–US association encoded within LA circuits, is required for both reactions and actions evoked by the CS.28, 152, 194 However, reactive freezing and active avoidance depend on different intra-amygdala circuits that emanate from LA.
While connections from LA to CeA provide a key substrate of freezing, lesions of CeA enhance, rather than impair, the acquisition and expression of avoidance.141, 142, 195 CeA lesions also rescue performance in animals that fail to express avoidance behavior due to excessive freezing, allowing these ‘poor performers’ to avoid normally.141, 142 The defensive actions opposed by CeA are underpinned by interactions between LA and BA.32, 141, 152, 196, 197, 198 BA projects robustly to the nucleus accumbens (NAcc).199, 200, 201 NAcc has been implicated in appetitive instrumental behavior,136, 202, 203, 204, 205, 206, 207 as have BA neurons that project to NAcc.205, 208, 209, 210 Recent findings demonstrate that active avoidance requires NAcc,29, 33, 35, 211, 212, 213, 214, 215 as well as the flow of information from BA to NAcc.211 Thus, while LA is common to the circuits of reaction and action, distinct outputs of LA give rise to different amygdala output pathways underlying freezing reactions (CeA–PAG) and avoidance actions (BA–NAcc). In the variant of active avoidance conditioning called escape from threat, rats learn to perform actions reinforced solely by CS termination. The amygdala circuits underlying escape from threat mirror those of signaled active avoidance—LA and BA are required, but CeA is not.152
Voltammetry studies of dopamine release in NAcc demonstrate interesting similarities between appetitive instrumental behavior and active avoidance. On the appetitive side, it has been shown that dopamine levels begin to ramp up when the subject is presented with a cue that predicts the availability of sucrose, peaking when a lever-press is emitted to obtain that reinforcer.216 A comparable result has been reported using a lever-press signaled active avoidance paradigm. Presentation of a warning signal elicited an increase in NAcc dopamine, preceding a successful avoidance response. However, if no such dopamine increase was observed, subjects failed to avoid.27 In a follow-up experiment, presentation of an aversive Pavlovian CS caused a decrease in NAcc dopamine release,27 suggesting that defensive actions and reactions have a distinct neurochemistry in NAcc. Combined with evidence that BA-NAcc projections have a role in appetitive instrumental action,205, 208, 209, 210 these data are consistent with the idea that avoidance is acquired through instrumental learning of an action–outcome relationship.
Lesion and inactivation experiments demonstrate that the infralimbic region of the medial prefrontal cortex (PFCIL) has a key role in the transition from freezing to avoidance.156 Pre-training lesion or inactivation of PFCIL cause prolonged freezing during training, delaying acquisition of the avoidance response. Once avoidance has been acquired, inactivation of PFCIL impairs the transition from reaction to action, causing a return of freezing, which occludes the avoidance response. Intriguingly, the expression of c-Fos (a marker of neuronal activation) in PFCIL distinguishes good and poor performers in an unsignaled active avoidance paradigm, in which strong PFCIL recruitment correlates with high avoidance and low freezing.217 These data suggest that PFCIL functions to suppress defensive reactions and facilitate defensive actions, toggling between behaviors controlled by distinct amygdala output pathways. Thus, PFCIL inhibits behavior driven by the CeA-PAG projection (that is, freezing), facilitating behavior driven by the BA–NAcc projection (that is, avoidance).
But what recruits PFCIL to this task? In the initial phases of training, freezing is high but not total, and many animals will eventually emit the avoidance response randomly. We speculate that these early instances of the response allow subjects to detect the avoidance contingency, engaging PFCIL to initiate the transition from reaction to action. This idea is consistent with previous work implicating PFC in the detection of aversive contingencies.218 It also explains why poor performers tend to be high freezers141, 142, 217 –because of their strong Pavlovian reactions to the CS, these subjects are never able to recruit PFCIL.
The results described so far support the idea that at least two distinct types of learning are at work in the avoidance paradigm. An LA–CeA–PAG pathway mediates the acquisition and expression of CS-evoked freezing. This circuit directly opposes active avoidance, which requires the recruitment of an LA–BA–NAcc pathway later in training. Because defensive reactions and actions are acquired at different points directly conflict with one another, and depend on dissociable circuits, we argue that they are conditioned through two distinct processes. These data provide empirical support for our model, in which the acquisition of defensive reactions is the first factor in avoidance learning, while the acquisition of defensive actions is the second. The first factor is clearly Pavlovian; the second factor may well be instrumental, as suggested by the shared substrates of appetitive instrumental behavior and active avoidance.
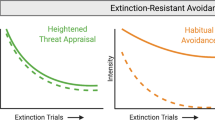
In addition to the sequential acquisition of defensive reactions and actions, we argue for a third factor, defensive habit, in avoidance learning. With prolonged training, the avoidance response becomes independent of the amygdala.142, 219 Because both defensive reactions and actions require amygdala circuits, we believe that the transition to amygdala-independent behavior demarcates a habitual phase of avoidance training. Habits are maintained despite a weakened connection with the reinforcing event that initially supported learning.136, 137, 138 The persistence of habit in the absence of reinforcement accounts for the observation that well-trained (habitual) avoidance responses tend to resist extinction, solving what has been considered a conceptual problem in the field.49, 60, 62
Habit learning has been explored extensively using appetitive reinforcers. These studies demonstrate that the initial substrates of instrumental learning give way to a new habit circuitry involving the dorsal striatum.140, 220 Extensive avoidance training may involve a similar transition, moving from outcome-dependent defensive action that involves NAcc to an outcome-independent defensive habit that involves dorsal striatum. Future work should explore the role of the striatal habit circuit in well-trained avoidance behaviors. In addition, activity in the prelimbic region of the prefrontal cortex correlates with avoidance behavior that persists under extinction conditions.29 We interpret these data to suggest a role for prelimbic prefrontal cortex in defensive habits that continue despite a reduced connection with reinforcing stimuli. This may be relevant to the persistence of avoidance in anxiety disorders, which we will discuss below.
Relatively few studies have examined the brain mechanisms of active avoidance in humans. However, the results from these are broadly consistent with the animal literature. Thus, studies using functional imaging have implicated the amygdala, nucleus accumbens, medial prefrontal cortex and other areas, including habit circuits of the dorsal striatum, in active avoidance in humans.221, 222, 223
In summary, we argue that avoidance learning proceeds through three distinct phases, each associated with its own neural circuitry. The first phase is Pavlovian, involving defensive reactions underpinned by an LA–CeA–PAG pathway. The second is instrumental and involves defensive actions that require an LA–BA–NAcc pathway. In order to transition from reaction to action, PFCIL is recruited to suppress freezing and facilitate avoidance. The third and final phase involves defensive habits, which are independent of the amygdala and may depend on the dorsal striatum.
Therapeutic implications
Anxious people often avoid situations in which threats may appear. Avoidance is a very effective way of reducing symptoms associated with fear and anxiety, and avoidant behaviors are negatively reinforced by their success in doing so. But excessive avoidance can also prevent one from learning which situations are actually dangerous. A person with social anxiety who avoids parties loses the opportunity to learn who is friendly and who is not. For good reason, then, avoidance is viewed negatively in the clinical literature.1, 2, 3, 4, 5, 6, 7, 8 The distinction between adaptive and maladaptive avoidance is thus very important.
Discussions of maladaptive avoidance in humans often start with Mower’s two-factor fear theory and ways to improve upon it.52, 74, 75, 224, 225 However, we argue that defensive actions like avoidance are reinforced and motivated by nonconscious processes rather than reduction of conscious fear. Evidence from studies of brain-lesioned patients demonstrates that subcortical regions such as the amygdala mediate conditioned defensive responses to threat, but not the conscious experience of fear.121, 122 In place of fear reduction, we reconceive of the reinforcer in avoidance learning as cellular and molecular events in the circuits underpinning defensive action rather than conscious feelings (see above). These changes establish a learned relationship between stimuli and strengthen active responses. This should not be taken to mean that subjective fear has no role in human anxiety and avoidance, but instead that subjective fear reduction is not what causes avoidance to be acquired and sustained. We argue that this view is likely to open more fruitful paths in the effort to understand avoidance.
To pursue the distinction between adaptive and maladaptive avoidance we build on the clinical notion of active and passive coping strategies. With passive coping harm is avoided or postponed by withholding actions to the threat, while active coping avoids harm by performing actions that engage with and control the threat. Active coping strategies help humans adapt and get back to routine life in the aftermath of trauma,226 and research in patients with anxiety disorders shows the virtues of active engagement as part of the therapeutic process.4, 227 Laboratory analogs of passive coping include freezing and passive avoidance, while active avoidance and escape from threat mirror active coping strategies.
Research on animal behavior has yielded important insights into the brain substrates of active coping. For example, the escape from threat variant of active avoidance conditioning shows how rats learn to perform responses that are negatively reinforced by CS termination. To do this, the passive coping response, freezing, must be inhibited to allow the active coping response to emerge (action cannot be taken while freezing). Through active coping, the animals gain control over the threatening circumstances. This should not mean that active coping is always adaptive and passive coping is always maladaptive. Freezing and passive avoidance are adaptive except when used excessively and begin to interfere with daily life, and active avoidance is adaptive unless avoidance becomes excessive and habitual.
Concepts such as coping and control clearly imply cognitive processes. But the involvement of cognitive processing should not be confused with the involvement of conscious experience.30 As we have argued, the learning underlying reactions, actions and habits is implicit; though conscious experiences may occur, they are not the basis of the learning.
Active avoidance paradigms in animals allow us to make two distinctions relevant to clinical problems. First, as noted above, most animals that undergo active avoidance training learn to actively cope with a dangerous situation. A small percentage, though, fail to express active avoidance during and after training. Instead, they exhibit a maladaptive form of coping in which excessive CeA-mediated freezing prevents them from expressing an active, NAcc-mediated avoidance response.142 By contrast, animals that are able to express an active strategy recruit medial prefrontal cortex, especially PFCIL, to toggle amygdala output pathways—control is shifted from CeA outputs that mediate freezing to the BA outputs that mediate active avoidance. Connections between medial PFC and the amygdala in humans have been implicated in other aspects of emotion regulation in the face of stress.228, 229 A treatment that is able to suppress excessive CeA-mediated reactions might be very useful in facilitating the acquisition of active coping skills in those predisposed toward passive coping. Treatments that enhance medial PFC activity may have a comparable effect. Finally, because NAcc dopamine responses occur on successful avoidance trials, but not when the subject fails to avoid,27 dopaminergic systems may be another target for therapies designed to facilitate active coping skills.
Second, while overcoming excessive freezing makes active coping possible, active coping itself can be adaptive or pathological. If avoidance, once learned, then becomes excessive, and comes to interfere with daily life, then it loses it adaptive qualities (just as freezing becomes maladaptive when excessive). This typically happens when the avoidance response becomes habitual. When this occurs, another branch point is reached, and there are additional opportunities for both adaptive and maladaptive coping. For example, during flu season, it can be wise to engage in more preventative hand washing than usual in order to avoid infection, particularly if one interacts with a large number of people. In this example, normal life activity is facilitated by an adaptive avoidance behavior. If that same behavior becomes habitual, it serves an adaptive function by not requiring constant intentional control over the behavior. But when it continues even in the absence of any threat because the individual does not take the opportunity to check if circumstances have changed, the habit has become maladaptive. In the case of obsessive-compulsive disorder, hand washing can become excessive and even injurious, persisting regardless of whether there exists any substantial risk of illness. Extinction-resistant forms of habitual avoidance may be particularly relevant to the compulsive behavior observed in many anxiety and addictive disorders.
Studies of active avoidance offer the opportunity to unravel both beneficial and pathological aspects of avoidance. It is the element of controllability, which puts the brakes on reactive defensive behaviors, that makes active avoidance useful. Animals and people who are able to engage in active avoidance may constitute the population of resilient individuals. Understanding the neurological differences underlying adaptive and maladaptive forms of coping may help reveal pharmaceutical or behavioral treatments that facilitate beneficial therapeutic coping strategies.
It is of interest that maladaptive avoidance is not simply a feature of anxiety disorders. It also occurs in people with obsessive-compulsive disorder, depression, suicidal tendencies and autism.26, 230 This is consistent with the idea that pathological avoidance is a domain or dimension231 spanning several diagnostic categories, rather than a symptom that identifies people with a particular diagnosis.
A final point to consider is whether maladaptive avoidance in humans is based on prior Pavlovian learning. Many have commented on the failure of patients to recount some previous experience that is at the root of their problems.7, 232, 233 This is viewed as evidence against a learning account of phobias, for exmaple. But using self-report in this way may underestimate the contribution of prior conditioning. The circuits involved in Pavlovian conditioning, as noted above, operate implicitly and may undergo learning independent of what the conscious mind notices and remembers. Moreover, intensely stressful events can impair memory formation and lead to amnesia for the event. Further, non-associative accounts of phobic acquisition have also been used to dismiss the role of learning.232 But non-associative accounts are still learning accounts (in other words, non-associative influences on behavior also involve learning). Regardless, the basic neuroscience of how over-responsivity to threats prevents active avoidance, and how the shift from instrumental to habitual behavior maintains maladaptive avoidance, is relevant to these clinical problems.
Conclusion
It is safe to say the research on avoidance has finally begun to extract itself from the damaging clutches of criticisms of the past. The new wave of research is showing vibrant signs of life, and beginning to reveal not only interactions between Pavlovian and action learning processes but also the previously unappreciated role of habit learning.
In some sense, the demise of avoidance research several decades ago has resulted in lost time and missed opportunities. But in another sense, it also allowed the field to focus on the neural basis of Pavlovian processes and thereby build up an impressive body of knowledge that is now also aiding the quest to understand active avoidance. The information obtained is allowing active avoidance to be approached with new concepts and new methods, and with new hope for a deeper understanding what avoidance is, how it works in the brain, and how and why it helps some but impairs others in daily life. Better understanding of avoidance circuits will hopefully also lead to new ideas about treatment for maladaptive avoidance, including ways to shift the neural control of behavior in ways typical of resilient individuals.
References
Marks I . Fears, Phobias, and Rituals: Panic, Anxiety and Their Disorders. Oxford University Press: New York, NY, USA, 1987.
Barlow DH . Anxiety and Its Disorders: The Nature and Treatment of Anxiety and Panic. Guilford Press: New York, NY, USA, 2002.
Lazarus RS . Psychological Stress and the Coping Process. McGraw Hill: New York, NY, USA, 1966.
Holahan CJ, Moos RH . Risk, resistance, and psychological distress: a longitudinal analysis with adults and children. J Abnorm Psychol 1987; 96: 3–13.
Foa EB, Kozak MJ . Emotional processing of fear: exposure to corrective information. Psychol Bull 1986; 99: 20–35.
Mineka S . The role of fear in theories of avoidance learning, flooding, and extinction. Psychol Bull 1979; 86: 985–1010.
Rachman S . Anxiety 2nd edn Psychology Press: Hove, England, 2004.
Hofmann SG . Emotion in Therapy: From Science to Practice. Guilford Press: New York, NY, USA, 2016.
Mowrer OH . A stimulus-response analysis of anxiety and its role as a reinforcing agent. Psychol Rev 1939; 46: 553–565.
Mowrer OH . Learning Theory and Behavior. Wiley: New York, NY, USA, 1960.
Mowrer OH . Anxiety-reduction and learning. J Exp Psychol 1940; 27: 497–516.
Kapp BS, Pascoe JP, Bixler MA The amygdala: a neuroanatomical systems approach to its contributions to aversive conditioning. In: Buttlers N, Squire LR (eds). Neuropsychology of Memory. Guilford: New York, NY, USA, 1984 pp 473–488.
LeDoux JE, Sakaguchi A, Reis DJ . Subcortical efferent projections of the medial geniculate nucleus mediate emotional responses conditioned to acoustic stimuli. J Neurosci 1984; 4: 683–698.
Hitchcock J, Davis M . Lesions of the amygdala but not of the cerebellum or red nucleus block conditioned fear as measured with the potentiated startle paradigm. Behav Neurosci 1986; 100: 11–22.
LeDoux JE . Emotion circuits in the brain. Annu Rev Neurosci 2000; 23: 155–184.
Johansen JP, Cain CK, Ostroff LE, LeDoux JE . Molecular mechanisms of fear learning and memory. Cell 2011; 147: 509–524.
Duvarci S, Pare D . Amygdala microcircuits controlling learned fear. Neuron 2014; 82: 966–980.
Pape HC, Pare D . Plastic synaptic networks of the amygdala for the acquisition, expression, and extinction of conditioned fear. Physiol Rev 2010; 90: 419–463.
Fanselow MS, Poulos AM . The neuroscience of mammalian associative learning. Annu Rev Psychol 2005; 56: 207–234.
Tovote P, Fadok JP, Luthi A . Neuronal circuits for fear and anxiety. Nat Rev Neurosci 2015; 16: 317–331.
Rosen JB, Schulkin J . From normal fear to pathological anxiety. Psychol Rev 1998; 105: 325–350.
LeDoux JE . Synaptic Self: How Our Brains Become Who We Are. Viking: New York, NY, USA, 2002.
Cain C, LeDoux JE Emotional processing and motivation: in search of brain mechanisms. In: Elliot AJ (ed). Handbook of Approach and Avoidance Motivation. Psychology Press, Taylor and Francis Group, LLC: New York, NY, USA, 2008 pp 17–34.
LeDoux JE, Schiller D, Cain C Emotional reaction and action: from threat processing to goal-directed behavior. In: Gazzaniga MS (ed). The Cognitive Neurosciences, 4th edn. MIT Press: Cambridge, MA, USA, 2009 pp 905–924.
Krypotos AM, Effting M, Kindt M, Beckers T . Avoidance learning: a review of theoretical models and recent developments. Front Behav Neurosci 2015; 9: 189.
Servatius RJ . Editorial: avoidance: from basic science to psychopathology. Front Behav Neurosci 2016; 10: 15.
Oleson EB, Gentry RN, Chioma VC, Cheer JF . Subsecond dopamine release in the nucleus accumbens predicts conditioned punishment and its successful avoidance. J Neurosci 2012; 32: 14804–14808.
Campese VD, Sears RM, Moscarello JM, Diaz-Mataix L, Cain CK, LeDoux JE . The neural foundations of reaction and action in aversive motivation. Curr Top Behav Neurosci 2016; 27: 171–195.
Bravo-Rivera C, Roman-Ortiz C, Montesinos-Cartagena M, Quirk GJ . Persistent active avoidance correlates with activity in prelimbic cortex and ventral striatum. Front Behav Neurosci 2015; 9: 184.
LeDoux JE . Anxious: Using the Brain to Understand and Treat Fear and Anxiety. Viking: New York, NY, USA, 2015.
Herrnstein RJ . Method and theory in the study of avoidance. Psychol Rev 1969; 76: 49–69.
Killcross S, Robbins TW, Everitt BJ . Different types of fear-conditioned behaviour mediated by separate nuclei within amygdala. Nature 1997; 388: 377–380.
Fernando AB, Urcelay GP, Mar AC, Dickinson A, Robbins TW . Safety signals as instrumental reinforcers during free-operant avoidance. Learn Mem 2014; 21: 488–497.
Jiao X, Beck KD, Myers CE, Servatius RJ, Pang KC . Altered activity of the medial prefrontal cortex and amygdala during acquisition and extinction of an active avoidance task. Front Behav Neurosci 2015; 9: 249.
Bravo-Rivera C, Roman-Ortiz C, Brignoni-Perez E, Sotres-Bayon F, Quirk GJ . Neural structures mediating expression and extinction of platform-mediated avoidance. J Neurosci 2014; 34: 9736–9742.
Watson JB . Behaviorism. W.W. Norton: New York, NY, USA, 1925.
Pavlov IP . Conditioned Reflexes. Dover: New York, NY, USA, 1927.
Watson JB, Rayner R . Conditioned emotional reactions. J Exp Psychol 1920; 3: 1–14.
Skinner BF . The Behavior of Organisms: An Experimental Analysis. Appleton-Century-Crofts: New York, NY, USA, 1938.
Hull CL . Principles of Behavior. Appleton-Century-Crofts: New York, NY, USA, 1943.
Tolman EC . Purposive Behavior in Animals and Men. Century: New York, NY, USA, 1932.
Freud S . The Problem of Anxiety. W. W. Norton: New York, NY, USA, 1936.
Rescorla RA, Solomon RL . Two process learning theory: relationships between Pavlovian conditioning and instrumental learning. Psychol Rev 1967; 74: 151–182.
Overmier JB, Brackbill RM . On the independence of stimulus evocation of fear and fear evocation of responses. Behav Res Ther 1977; 15: 51–56.
Miller NE Learnable drives and rewards. In: Stevens SS (ed). Handbook of Experimental Psychology. Wiley: New York, NY, USA, 1951 pp 435–472.
Mowrer OH . Two-factor learning theory: summary and comment. Psychol Rev 1951; 58: 350–354.
McAllister DE, McAllister WR Fear theory and aversively motivated behavior: Some controversial issues. In: Denny MR (ed). Fear, Avoidance, and Phobias: A Fundamental Analysis. Erlbaum: Hillsdale, NJ, USA, 1991.
McAllister WR, McAllister DE Behavioral measurement of conditioned fear. In: Brush FR (ed). Aversive Conditioning and Learning. Academic Press: New York, 1971 pp 105–179.
Masterson FA, Crawford M . The defense motivation system: a theory of avoidance behavior. Behav Brain Sci 1982; 5: 661–696.
Miller NE . Studies of fear as an acquirable drive: I. Fear as motivation and fear reduction as reinforcement in the learning of new responses. J Exp Psychol 1948; 38: 89–101.
Mowrer OH . On the dual nature of learning: a reinterpretation of “conditioning” and “problem solving”. Harvard Educ Rev 1947; 17: 102–148.
Levis DJ The case for a return to a two-factor theory of avoidance: the failure of non-fear interpretations. In: Klein SB, Mowrer RR (eds). Contemporary Learning Theories: Pavlovian Conditioning and the Status of Traditional Learning Theory. Lawrence Erlbaum Assn: Hillsdale, NJ, USA, 1989 pp 227–277.
Mowrer OH, Lamoreaux RR . Fear as an intervening variable in avoidance conditioning. J Comp Psych 1946; 39: 29–50.
Miller NE . An experimental investigation of acquired drives. Psychol Bull 1941; 38: 534–535.
Dinsmoor JA . Punishment. I. The avoidance hypothesis. Psychol Rev 1954; 61: 34–46.
Schoenfeld WN An experimental approach to anxiety, escape, and avoidance behavior. In: Hoch PH, Zubin J (eds). Anxiety. Grune & Stratton: New York, NY, USA, 1950 pp 70–99.
Solomon RL, Wynne LC . Traumatic avoidance learning: the principles of anxiety conservation and partial irreversibility. Psychol Rev 1954; 61: 353–385.
Kamin LJ . The effects of termination of the CS and avoidance of the US on avoidance learning. J Comp Physiol Psychol 1956; 49: 420–424.
Bolles RC . Species-specific defense reactions and avoidance learning. Psychol Rev 1970; 77: 32–48.
Bolles RC The avoidance learning problem. In: Bower GH (ed). The Psychology of Learning and Motivation vol. 6. Academic Press: New York, NY, USA, 1972 pp 97–145.
Trapold MA, Overmier JB The second learning process in instrumental learning. In: Black AA, Prokasy WF (eds). Classical Conditioning 2: Current Research and Theory. Appleton-Century-Crofts: New York, NY, USA, 1972 pp 427–452.
Seligman ME, Johnston JC A cognitive theory of avoidance learning. In: McGuigan FJ, Lumsden DB (eds). Contemporary Approaches to Conditioning and Learning. V. H. Winston & Sons: Oxford, UK, 1973 pp 69–110.
Gray JA . The Psychology of Fear and Stress vol. 2. Cambridge University Press: New York, NY, USA, 1971.
Overmier JB, Bull JA, Pack K . On instrumental response interaction as explaining the influences of pavlovian cs's upon avoidance behavior. Learn Motiv 1971; 2: 103–112.
Riccio DC, Silvestri R . Extinction of avoidance behavior and the problem of residual fear. Behav Res Ther 1973; 11: 1–9.
Rescorla RA, LoLordo VM . Inhibition of avoidance behavior. J Comp Physiol Psychol 1965; 59: 406–412.
Weiskrantz L . Behavioral changes associated with ablation of the amygdaloid complex in monkeys. J Comp Physiol Psychol 1956; 49: 381–391.
Goddard G . Functions of the amygdala. Psychol Rev 1964; 62: 89–109.
Grossman SP . A Textbook of Physiological Psychology. John Wiley & Sons, 1967.
Thompson R . A Behavioral Atlas of the Rat Brain. Oxford University Press: New York, NY, USA, 1978.
Sarter MF, Markowitsch HJ . Involvement of the amygdala in learning and memory: a critical review, with emphasis on anatomical relations. Behav Neurosci 1985; 99: 342–380.
Dollard J, Miller NE . Personality and Psychotherapy: An Analysis in Terms of Learning, Thinking, and Culture. McGraw-Hill: New York, NY, USA, 1950.
Stampfl TG, Levis DJ . Essentials of implosive therapy: a learning-theory-based psychodynamic behavioral therapy. J Abnorm Psychol 1967; 72: 496–503.
Hofmann SG . Cognitive processes during fear acquisition and extinction in animals and humans: implications for exposure therapy of anxiety disorders. Clin Psychol Rev 2008; 28: 199–210.
Dymond S, Roche B . A contemporary behavior analysis of anxiety and avoidance. Behav Anal 2009; 32: 7–27.
Gabriel M, Foster K, Orona E, Saltwick SE, Stanton M Neuronal activity of cingulate cortex, anteroventral thalamus, and hippocampal formation in discriminative conditioning: encoding and extraction of the significance of conditional stimuli. In: Sprague JM, Epstein AN (eds). Progress in Psychobiology and Physiological Psychology vol. 9. Academic Press: New York, NY, USA, 1980 pp 126–232.
Fonberg E . The role of the amygdaloid nucleus in animal behaviour. Prog Brain Res 1968; 22: 273–281.
Squire L . Memory and Brain. Oxford University Press: New York, NY, USA, 1987.
Kandel ER . Cellular Basis of Behavior: An Introduction to Behavioral Neurobiology. W.H. Freeman and Company: San Fransisco, CA, USA, 1976.
Carew TJ, Walters ET, Kandel ER . Associative learning in Aplysia: cellular correlates supporting a conditioned fear hypothesis. Science 1981; 211: 501–504.
Cohen DH . Involvement of the avian amygdalar homologue (archistriatum posterior and mediale) in defensively conditioned heart rate change. J Comp Neurol 1975; 160: 13–35.
Thompson RF . The neurobiology of learning and memory. Science 1986; 233: 941–947.
Kapp BS, Frysinger RC, Gallagher M, Haselton JR . Amygdala central nucleus lesions: effect on heart rate conditioning in the rabbit. Physiol Behav 1979; 23: 1109–1117.
Davis M . Sensitization of the acoustic startle reflex by footshock. Behav Neurosci 1989; 103: 495–503.
Kamin LJ, Brimer CJ, Black AH . Conditioned suppression as a monitor of fear ofthe CS in the course of avoidance training. J Comp Physiol Psychol 1963; 56: 497–501.
Blanchard RJ, Blanchard DC . Crouching as an index of fear. J Comp Physiol Psychol 1969; 67: 370–375.
Stebbins WC, Smith OA . Cardiovascular concomitants of the conditioned emotional response in monkey. Science 1964; 144: 881–883.
Heimer L, Robards MJ . Neuroanatomical Tract-Tracing Methods. Plenum Press: New York, NY, USA, 1981.
Emson PC . Chemical Neuroanatomy. Raven Press, 1983.
Solomon RL, Kamin LJ, Wynne LC . Traumatic avoidance learning: the outcomes of several extinction procedures with dogs. J Abnorm Psychol 1953; 48: 291–302.
Bolles RC, Fanselow MS . A perceptual-defensive-recuperative model of fear and pain. Behav Brain Sci 1980; 3: 291–323.
Annau Z, Kamin LJ . The conditioned emotional response as a function of the intensity of the US. J Comp Physiol Psychol 1961; 54: 428–432.
Starr MD, Mineka S . Determinants of fear over the course of avoidance learning. Learn Motiv 1977; 8: 331–350.
Lang PJ Fear reduction and fear behavior: problems in treating a construct. In: Schlien JM (ed). Research in Psychotherapy vol. 3. American Psychological Association: Washington, DC, USA, 1968 pp 90–103.
Zinbarg RE . Concordance and synchrony in measures of anxiety and panic reconsidered: a hierarchical model of anxiety and panic. Behav Ther 1998; 29: 301–323.
Bindra D . A unified interpretation of emotion and motivation. Ann N Y Acad Sci 1969; 159: 1071–1083.
Bolles RC . Theory of Motivation. Harper and Row: New York, NY, USA, 1967.
Logan FA, Wagner AR . Reward and Punishment. Allyn and Bacon: Boston, MA, USA, 1965.
Davis M, Hitchcock JM, Rosen JB Anxiety and the amygdala: pharmacological and anatomical analysis of the fear-potentiated startle paradigm. In: Bower GH (ed). The Psychology of Learning and Motivation vol 21. Academic Press: San Diego, CA, USA, 1987 pp 263–305.
LeDoux JE . The Emotional Brain. Simon and Schuster: New York, NY, USA, 1996.
Lang PJ, Bradley MM, Cuthbert BN . Emotion, attention, and the startle reflex. Psychol Rev 1990; 97: 377–395.
Lang PJ, Davis M . Emotion, motivation, and the brain: reflex foundations in animal and human research. Prog Brain Res 2006; 156: 3–29.
Adolphs R . The biology of fear. Curr Biol 2013; 23: R79–R93.
Perusini JN, Fanselow MS . Neurobehavioral perspectives on the distinction between fear and anxiety. Learn Mem 2015; 22: 417–425.
Fanselow MS . Neural organization of the defensive behavior system responsible for fear. Psychon Bull Rev 1994; 1: 429–438.
Panksepp J . Affective Neuroscience. Oxford University Press: New York, NY, USA, 1998.
Panksepp J, Fuchs T, Iacabucci P . The basic neuroscience of emotional experiences in mammals: the case of subcortical FEAR circuitry and implications for clinical anxiety. Appl Anim Behav Sci 2011; 129: 1–17.
Rau V, DeCola JP, Fanselow MS . Stress-induced enhancement of fear learning: an animal model of posttraumatic stress disorder. Neurosci Biobehav Rev 2005; 29: 1207–1223.
Milad MR, Quirk GJ . Fear extinction as a model for translational neuroscience: ten years of progress. Annu Rev Psychol 2012; 63: 129–151.
Marx MH . Intervening variable or hypothetical construct? Psychol Rev 1951; 58: 235–247.
LeDoux J . Rethinking the emotional brain. Neuron 2012; 73: 653–676.
LeDoux JE . Coming to terms with fear. Proc Natl Acad Sci USA 2014; 111: 2871–2878.
LeDoux JE . The slippery slope of fear. Trends Cogn Sci 2013; 17: 155–156.
Dolan RJ, Vuilleumier P . Amygdala automaticity in emotional processing. Ann N Y Acad Sci 2003; 985: 348–355.
Mineka S, Ohman A . Phobias and preparedness: the selective, automatic, and encapsulated nature of fear. Biol Psychiatry 2002; 52: 927–937.
Vuilleumier P, Pourtois G . Distributed and interactive brain mechanisms during emotion face perception: evidence from functional neuroimaging. Neuropsychologia 2007; 45: 174–194.
Whalen PJ, Kagan J, Cook RG, Davis FC, Kim H, Polis S et al. Human amygdala responsivity to masked fearful eye whites. Science 2004; 306: 2061.
Luo Q, Holroyd T, Majestic C, Cheng X, Schechter J, Blair RJ . Emotional automaticity is a matter of timing. J Neurosci 2010; 30: 5825–5829.
Lissek S, Levenson J, Biggs AL, Johnson LL, Ameli R, Pine DS et al. Elevated fear conditioning to socially relevant unconditioned stimuli in social anxiety disorder. Am J Psychiatry 2008; 165: 124–132.
Hariri AR, Tessitore A, Mattay VS, Fera F, Weinberger DR . The amygdala response to emotional stimuli: a comparison of faces and scenes. NeuroImage 2002; 17: 317–323.
Anderson AK, Phelps EA . Is the human amygdala critical for the subjective experience of emotion? Evidence of intact dispositional affect in patients with amygdala lesions. J Cogn Neurosci 2002; 14: 709–720.
Feinstein JS, Buzza C, Hurlemann R, Follmer RL, Dahdaleh NS, Coryell WH et al. Fear and panic in humans with bilateral amygdala damage. Nat Neurosci 2013; 16: 270–272.
Phelps EA . Emotion and cognition: insights from studies of the human amygdala. Annu Rev Psychol 2006; 57: 27–53.
Weiskrantz L . Consciousness Lost and Found: A Neuropsychological Exploration. Oxford University Press: New York, NY, USA, 1997.
Morris JS, DeGelder B, Weiskrantz L, Dolan RJ . Differential extrageniculostriate and amygdala responses to presentation of emotional faces in a cortically blind field. Brain 2001; 124 (Pt 6): 1241–1252.
Tamietto M, de Gelder B . Neural bases of the non-conscious perception of emotional signals. Nat Rev Neurosci 2010; 11: 697–709.
Bertini C, Cecere R, Ladavas E . I am blind, but I "see" fear. Cortex 2013; 49: 985–993.
LeDoux J, Pine DS . Using neuroscience to help understand fear and anxiety: a two-system framework. Am J Psychiatry; advance online publication, 9 September; appiajp201616030353 [e-pub ahead of print].
Downey G, Mougios V, Ayduk O, London BE, Shoda Y . Rejection sensitivity and the defensive motivational system: insights from the startle response to rejection cues. Psychol Sci 2004; 15: 668–673.
Lang PJ, Bradley MM . Appetitive and defensive motivation: goal-directed or goal-determined? Emot Rev 2013; 5: 230–234.
Dickinson A, Balleine B The role of learning in the operation of motivational systems. In: Gallistel CR (ed). Steven’s Handbook of Experimental Psychology: Learning, Motivation and Emotion 3rd edn vol. 3. John Wiley & Sons: New York, NY, USA, 2002 pp 497–534.
Church RM . Systematic effect of random error in the yoked control design. Psychol Bull 1964; 62: 122–131.
Church RM The yoked control design. In: Archer T, Nilsson L (eds). Aversion, Avoidance and Anxiety: Perspectives on Aversively Motivated Behavior. Erlbaum: Hillsdale, NJ, USA, 1989 pp 403–413.
Clarke RJ . Flight behaviour elicited by electrical stimulation of the hypothalamus and midbrain in rats: escape and avoidance properties. Master of Science thesis, University of British Columbia, Vancouver, Canada 1972.
Brown JL, Hunsperger RW, Rosvold HE . Defence, attack, and flight elicited by electrical stimulation of the hypothalamus of the cat. Exp Brain Res 1969; 8: 113–129.
Cardinal RN, Parkinson JA, Hall J, Everitt BJ . Emotion and motivation: the role of the amygdala, ventral striatum, and prefrontal cortex. Neurosci Biobehav Rev 2002; 26: 321–352.
Dickinson A . Action and habits: the development of behavioural autonomy. Philos Trans R Soc Lond B Biol Sci 1985; 308: 67–78.
Dezfouli A, Balleine BW . Habits, action sequences and reinforcement learning. Eur J Neurosci 2012; 35: 1036–1051.
Everitt BJ, Robbins TW . Neural systems of reinforcement for drug addiction: from actions to habits to compulsion. Nat Neurosci 2005; 8: 1481–1489.
Balleine BW, O'Doherty JP . Human and rodent homologies in action control: corticostriatal determinants of goal-directed and habitual action. Neuropsychopharmacology 2010; 35: 48–69.
Choi JS, Cain CK, LeDoux JE . The role of amygdala nuclei in the expression of auditory signaled two-way active avoidance in rats. Learn Mem 2010; 17: 139–147.
Lazaro-Munoz G, LeDoux JE, Cain CK . Sidman instrumental avoidance initially depends on lateral and Basal amygdala and is constrained by central amygdala-mediated Pavlovian processes. Biol Psychiatry 2010; 67: 1120–1127.
Mackintosh NJ . The Psychology of Animal Learning. Academic Press: New York, NY, USA, 1974.
Dickinson A Instrumental conditioning. In: Mackintosh NJ (ed). Animal Learning and Cognition. Academic Press: San Diego, CA, USA, 1994 pp 45–79.
Rescorla RA . Evidence for an association between the discriminative stimulus and the response-outcome association in instrumental learning. J Exp Psychol Anim Behav Process 1990; 16: 326–334.
Balleine B, Dickinson A . Instrumental performance following reinforcer devaluation depends upon incentive learning. Q J Exp Psychol B 1991; 43: 279–296.
Holland PC . Relations between Pavlovian-instrumental transfer and reinforcer devaluation. J Exp Psychol Anim Behav Process 2004; 30: 104–117.
Holland PC . Cognitive versus stimulus-response theories of learning. Learn Behav 2008; 36: 227–241.
Mackintosh NJ . Conditioning and Associative Learning. Oxford University Press: New York, NY, USA, 1983.
Dickinson A, Boakes RA (eds). Mechanisms of Learning and Motivation: A Memorial Volume to Jerzy Konorski. Erlbaum: Hillsdale, NJ, USA, 1979.
Cain CK, LeDoux JE . Escape from fear: a detailed behavioral analysis of two atypical responses reinforced by CS termination. J Exp Psychol Anim Behav Process 2007; 33: 451–463.
Amorapanth P, LeDoux JE, Nader K . Different lateral amygdala outputs mediate reactions and actions elicited by a fear-arousing stimulus. Nat Neurosci 2000; 3: 74–79.
Rescorla RA . Pavlovian conditioned fear in Sidman avoidance learning. J Comp Physiol Psychol 1968; 65: 55–60.
Morris RGM . Pavlovian conditioned inhibition of fear during shuttlebox avoidance behavior. Learn Motiv 1974; 5: 424–447.
De Houwer J, Crombez G, Baeyens F . Avoidance behavior can function as a negative occasion setter. J Exp Psychol Anim Behav Process 2005; 31: 101–106.
Moscarello JM, LeDoux JE . Active avoidance learning requires prefrontal suppression of amygdala-mediated defensive reactions. J Neurosci 2013; 33: 3815–3823.
LeDoux JE Emotion and the amygdala. In: Aggleton JP (ed). The Amygdala: Neurobiological Aspects of Emotion, Memory, and Mental Dysfunction. Wiley-Liss: New York, NY, USA, 1992 pp 339–351.
Davis M The role of the amygdala in conditioned fear. In: Aggleton JP (ed). The Amygdala: Neurobiological Aspects of Emotion, Memory, and Mental Dysfunction. Wiley-Liss: New York, NY, USA, 1992 pp 255–306.
Kapp BS, Whalen PJ, Supple WF, Pascoe JP Amygdaloid contributions to conditioned arousal and sensory information processing. In: Aggleton JP (ed). The Amygdala: Neurobiological Aspects of Emotion, Memory, and Mental Dysfunction. Wiley-Liss: New York, NY, USA, 1992 pp 229–254.
Maren S . Synaptic mechanisms of associative memory in the amygdala. Neuron 2005; 47: 783–786.
LeDoux JE, Ruggiero DA, Forest R, Stornetta R, Reis DJ . Topographic organization of convergent projections to the thalamus from the inferior colliculus and spinal cord in the rat. J Comp Neurol 1987; 264: 123–146.
Romanski LM, LeDoux JE, Clugnet MC, Bordi F . Somatosensory and auditory convergence in the lateral nucleus of the amygdala. Behav Neurosci 1993; 107: 444–450.
Bordi F, LeDoux JE . Response properties of single units in areas of rat auditory thalamus that project to the amygdala. II. Cells receiving convergent auditory and somatosensory inputs and cells antidromically activated by amygdala stimulation. Exp Brain Res 1994; 98: 275–286.
Rogan MT, LeDoux JE . LTP is accompanied by commensurate enhancement of auditory-evoked responses in a fear conditioning circuit. Neuron 1995; 15: 127–136.
Rogan MT, Staubli UV, LeDoux JE . Fear conditioning induces associative long-term potentiation in the amygdala. Nature 1997; 390: 604–607.
Quirk GJ, Armony JL, LeDoux JE . Fear conditioning enhances different temporal components of tone-evoked spike trains in auditory cortex and lateral amygdala. Neuron 1997; 19: 613–624.
Quirk GJ, Repa C, LeDoux JE . Fear conditioning enhances short-latency auditory responses of lateral amygdala neurons: parallel recordings in the freely behaving rat. Neuron 1995; 15: 1029–1039.
Repa JC, Muller J, Apergis J, Desrochers TM, Zhou Y, LeDoux JE . Two different lateral amygdala cell populations contribute to the initiation and storage of memory. Nat Neurosci 2001; 4: 724–731.
Maren S . Auditory fear conditioning increases CS-elicited spike firing in lateral amygdala neurons even after extensive overtraining. Eur J Neurosci 2000; 12: 4047–4054.
LeDoux JE, Cicchetti P, Xagoraris A, Romanski LM . The lateral amygdaloid nucleus: Sensory interface of the amygdala in fear conditioning. J Neurosci 1990; 10: 1062–1069.
Muller J, Corodimas KP, Fridel Z, LeDoux JE . Functional inactivation of the lateral and basal nuclei of the amygdala by muscimol infusion prevents fear conditioning to an explicit conditioned stimulus and to contextual stimuli. Behav Neurosci 1997; 111: 683–691.
Nader K, Majidishad P, Amorapanth P, LeDoux JE . Damage to the lateral and central, but not other, amygdaloid nuclei prevents the acquisition of auditory fear conditioning. Learn Mem 2001; 8: 156–163.
Grundemann J, Luthi A . Ensemble coding in amygdala circuits for associative learning. Curr Opin Neurobiol 2015; 35: 200–206.
Blair HT, Schafe GE, Bauer EP, Rodrigues SM, LeDoux JE . Synaptic plasticity in the lateral amygdala: a cellular hypothesis of fear conditioning. Learn Mem 2001; 8: 229–242.
Rodrigues SM, Schafe GE, LeDoux JE . Molecular mechanisms underlying emotional learning and memory in the lateral amygdala. Neuron 2004; 44: 75–91.
Pitkänen A, Savander V, LeDoux JE . Organization of intra-amygdaloid circuitries in the rat: an emerging framework for understanding functions of the amygdala. Trends Neurosci 1997; 20: 517–523.
Pare D, Smith Y . Intrinsic circuitry of the amygdaloid complex: common principles of organization in rats and cats. Trends Neurosci 1998; 21: 240–241.
Royer S, Martina M, Pare D . An inhibitory interface gates impulse traffic between the input and output stations of the amygdala. J Neurosci 1999; 19: 10575–10583.
Pascoe JP, Kapp BS . Electrophysiological characteristics of amygdaloid central nucleus neurons during Pavlovian fear conditioning in the rabbit. Behav Brain Res 1985; 16: 117–133.
Wilensky AE, Schafe GE, Kristensen MP, LeDoux JE . Rethinking the fear circuit: the central nucleus of the amygdala is required for the acquisition, consolidation, and expression of pavlovian fear conditioning. J Neurosci 2006; 26: 12387–12396.
Duvarci S, Popa D, Pare D . Central amygdala activity during fear conditioning. J Neurosci 2011; 31: 289–294.
Ciocchi S, Herry C, Grenier F, Wolff SB, Letzkus JJ, Vlachos I et al. Encoding of conditioned fear in central amygdala inhibitory circuits. Nature 2010; 468: 277–282.
Haubensak W, Kunwar PS, Cai H, Ciocchi S, Wall NR, Ponnusamy R et al. Genetic dissection of an amygdala microcircuit that gates conditioned fear. Nature 2010; 468: 270–276.
Li H, Penzo MA, Taniguchi H, Kopec CD, Huang ZJ, Li B . Experience-dependent modification of a central amygdala fear circuit. Nat Neurosci 2013; 16: 332–339.
LeDoux JE, Iwata J, Cicchetti P, Reis DJ . Different projections of the central amygdaloid nucleus mediate autonomic and behavioral correlates of conditioned fear. J Neurosci 1988; 8: 2517–2529.
Hitchcock JM, Davis M . Fear-potentiated startle using an auditory conditioned stimulus: effect of lesions of the amygdala. Physiol Behav 1987; 39: 403–408.
Kim JJ, Rison RA, Fanselow MS . Effects of amygdala, hippocampus, and periaqueductal gray lesions on short- and long-term contextual fear. Behav Neurosci 1993; 107: 1093–1098.
LaBar KS, LeDoux JE, Spencer DD, Phelps EA . Impaired fear conditioning following unilateral temporal lobectomy in humans. J Neurosci 1995; 15: 6846–6855.
Bechara A, Tranel D, Damasio H, Adolphs R, Rockland C, Damasio AR . Double dissociation of conditioning and declarative knowledge relative to the amygdala and hippocampus in humans. Science 1995; 269: 1115–1118.
LaBar KS, Gatenby JC, Gore JC, LeDoux JE, Phelps EA . Human amygdala activation during conditioned fear acquisition and extinction: a mixed-trial fMRI study. Neuron 1998; 20: 937–945.
Morris JS, Ohman A, Dolan RJ . A subcortical pathway to the right amygdala mediating "unseen" fear. Proc Natl Acad Sci USA 1999; 96: 1680–1685.
Morris JS, Ohman A, Dolan RJ . Conscious and unconscious emotional learning in the human amygdala. Nature 1998; 393: 467–470.
Mendez-Bertolo C, Moratti S, Toledano R, Lopez-Sosa F, Martinez-Alvarez R, Mah YH et al. A fast pathway for fear in human amygdala. Nat Neurosci 2016; 19: 1041–1049.
Cain CK, Choi JS, LeDoux JE Active avoidance and escape learning. In: Koob G, Thompson R, Le Moal M (eds). Encyclopedia of Behavioral Neuroscience. Elsevier: New York, NY, USA, 2010.
Moscarello JM, LeDoux J . Diverse effects of conditioned threat stimuli on behavior. Cold Spring Harb Symp Quant Biol 2014; 79: 11–19.
Maren S, Poremba A, Gabriel M . Basolateral amygdaloid multi-unit neuronal correlates of discriminative avoidance learning in rabbits. Brain Res 1991; 549: 311–316.
Poremba A, Gabriel M . Medial geniculate lesions block amygdalar and cingulothalamic learning-related neuronal activity. J Neurosci 1997; 17: 8645.
Gabriel M, Burhans L, Kashef A . Consideration of a unified model of amygdalar associative functions. Ann N Y Acad Sci 2003; 985: 206–217.
McDonald AJ . Topographical organization of amygdaloid projections to the caudatoputamen, nucleus accumbens, and related striatal-like areas of the rat brain. Neurosci 1991; 44: 15–33.
Groenewegen HJ, Wright CI, Beijer AV, Voorn P . Convergence and segregation of ventral striatal inputs and outputs. Ann N Y Acad Sci 1999; 877: 49–63.
Wright CI, Beijer AV, Groenewegen HJ . Basal amygdaloid complex afferents to the rat nucleus accumbens are compartmentally organized. J Neurosci 1996; 16: 1877–1893.
Blackburn JR, Pfaus JG, Phillips AG . Dopamine functions in appetitive and defensive behaviours. Prog Neurobiol 1992; 39: 247–279.
Carelli RM . Nucleus accumbens cell firing and rapid dopamine signaling during goal-directed behaviors in rats. Neuropharmacology 2004; 47 (Suppl 1): 180–189.
Kelley AE . Functional specificity of ventral striatal compartments in appetitive behaviors. Ann N Y Acad Sci 1999; 877: 71–90.
Everitt BJ, Parkinson JA, Olmstead MC, Arroyo M, Robledo P, Robbins TW . Associative processes in addiction and reward. The role of amygdala- ventral striatal subsystems. Ann N Y Acad Sci 1999; 877: 412–438.
Berridge KC . 'Liking' and 'wanting' food rewards: brain substrates and roles in eating disorders. Physiol Behav 2009; 97: 537–550.
Burhans LB, Gabriel M . Contextual modulation of conditioned responses: role of the ventral subiculum and nucleus accumbens. Behav Neurosci 2007; 121: 1243–1257.
Shiflett MW, Balleine BW . At the limbic-motor interface: disconnection of basolateral amygdala from nucleus accumbens core and shell reveals dissociable components of incentive motivation. Eur J Neurosci 2010; 32: 1735–1743.
Robbins TW, Cador M, Taylor JR, Everitt BJ . Limbic-striatal interactions in reward-related processes. Neurosci Biobehav Rev 1989; 13: 155–162.
Cador M, Robbins TW, Everitt BJ . Involvement of the amydala in stimulus-reward associations: interaction with the ventral striatum. Neurosci 1989; 30: 77–86.
Ramirez F, Moscarello JM, LeDoux JE, Sears RM . Active avoidance requires a serial Basal amygdala to nucleus accumbens shell circuit. J Neurosci 2015; 35: 3470–3477.
Wenzel JM, Rauscher NA, Cheer JF, Oleson EB . A role for phasic dopamine release within the nucleus accumbens in encoding aversion: a review of the neurochemical literature. ACS Chem Neurosci 2015; 6: 16–26.
Nakanishi S, Hikida T, Yawata S . Distinct dopaminergic control of the direct and indirect pathways in reward-based and avoidance learning behaviors. Neuroscience 2014; 282C: 49–59.
Lichtenberg NT, Kashtelyan V, Burton AC, Bissonette GB, Roesch MR . Nucleus accumbens core lesions enhance two-way active avoidance. Neuroscience 2014; 258: 340–346.
Oleson EB, Cheer JF . On the role of subsecond dopamine release in conditioned avoidance. Front Neurosci 2013; 7: 96.
Roitman MF, Stuber GD, Phillips PE, Wightman RM, Carelli RM . Dopamine operates as a subsecond modulator of food seeking. J Neurosci 2004; 24: 1265–1271.
Martinez RC, Gupta N, Lazaro-Munoz G, Sears RM, Kim S, Moscarello JM et al. Active vs. reactive threat responding is associated with differential c-Fos expression in specific regions of amygdala and prefrontal cortex. Learn Mem 2013; 20: 446–452.
Amat J, Baratta MV, Paul E, Bland ST, Watkins LR, Maier SF . Medial prefrontal cortex determines how stressor controllability affects behavior and dorsal raphe nucleus. Nat Neurosci 2005; 8: 365–371.
Poremba A, Gabriel M . Amygdala neurons mediate acquisition but not maintenance of instrumental avoidance behavior in rabbits. J Neurosci 1999; 19: 9635–9641.
Everitt BJ, Robbins TW . Drug addiction: updating actions to habits to compulsions ten years on. Annu Rev Psychol 2016; 67: 23–50.
Delgado MR, Jou RL, LeDoux JE, Phelps EA . Avoiding negative outcomes: tracking the mechanisms of avoidance learning in humans during fear conditioning. Front Behav Neurosci 2009; 3: 33.
Collins KA, Mendelsohn A, Cain CK, Schiller D . Taking action in the face of threat: neural synchronization predicts adaptive coping. J Neurosci 2014; 34: 14733–14738.
Schlund MW, Hudgins CD, Magee S, Dymond S . Neuroimaging the temporal dynamics of human avoidance to sustained threat. Behav Brain Res 2013; 257: 148–155.
Rachman S . The passing of the two-stage theory of fear and avoidance: fresh possibilities. Behav Res Ther 1976; 14: 125–131.
Bouton ME, Mineka S, Barlow DH . A modern learning theory perspective on the etiology of panic disorder. Psychol Rev 2001; 108: 4–32.
LeDoux JE, Gorman JM . A call to action: overcoming anxiety through active coping. Am J Psychiatry 2001; 158: 1953–1955.
van der Kolk B . The Body Keeps the Score: Brain, Mind, and Body in the Healing of Trauma. Viking: New York, NY, USA, 2014.
Raio CM, Phelps EA . The influence of acute stress on the regulation of conditioned fear. Neurobiol Stress 2015; 1: 134–146.
Maier SF, Watkins LR . Role of the medial prefrontal cortex in coping and resilience. Brain Res 2010; 1355: 52–60.
Gillan CM, Morein-Zamir S, Urcelay GP, Sule A, Voon V, Apergis-Schoute AM et al. Enhanced avoidance habits in obsessive-compulsive disorder. Biol Psychiatry 2014; 75: 631–638.
Cuthbert BN, Insel TR . Toward the future of psychiatric diagnosis: the seven pillars of RDoC. BMC Med 2013; 11: 126.
Poulton R, Menzies RG . Fears born and bred: toward a more inclusive theory of fear acquisition. Behav Res Ther 2002; 40: 197–208.
McNally RJ . On nonassociative fear emergence. Behav Res Ther 2002; 40: 169–172.
Acknowledgements
This work is supported by NIH Grants R56DA029053 and R01MH38874.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no conflict of interest.
Rights and permissions
This work is licensed under a Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-sa/4.0/
About this article

Cite this article
LeDoux, J., Moscarello, J., Sears, R. et al. The birth, death and resurrection of avoidance: a reconceptualization of a troubled paradigm. Mol Psychiatry 22, 24–36 (2017). https://doi.org/10.1038/mp.2016.166
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/mp.2016.166
This article is cited by
-
The genetic architecture of youth anxiety: a study protocol
BMC Psychiatry (2024)
-
Cyclin Y regulates spatial learning and memory flexibility through distinct control of the actin pathway
Molecular Psychiatry (2023)
-
A sex-specific role for the bed nucleus of the stria terminalis in proactive defensive behavior
Neuropsychopharmacology (2023)
-
Automated phenotyping of postoperative delirium-like behaviour in mice reveals the therapeutic efficacy of dexmedetomidine
Communications Biology (2023)
-
Generalization of beneficial exposure effects to untreated stimuli from another fear category
Translational Psychiatry (2023)
