

Abstract
RNA interference (RNAi) is a straightforward approach to study gene function from the in vitro cellular level to in vivo animal behavior. Although RNAi-mediated gene knockdown has become essentially routine in neuroscience over the past ten years, off-target effects of short hairpin RNAs (shRNAs) should be considered as the proper choice of control shRNA is critical in order to perform meaningful experiments. Luciferase shRNA (shLuc), targeting firefly luciferase, and scrambled shRNAs (shScrs) have been widely used as controls for vertebrate cell research. However, thorough validation of control shRNAs has not been made to date. Here, we performed thorough physiological and morphological studies against control shRNAs in mouse hippocampal CA1 pyramidal neurons. As expected, all control shRNAs exhibited normal basal synaptic transmission and dendritic morphology. However, to our surprise, shLuc exerted severe off-target effects on voltage-gated ion channel function, while the shScr had no detectable changes. These results indicate that thorough validation of shRNA is imperative and, in the absence of such validation, that shScr is the best available negative control for gene knockdown studies.
Significance Statement
RNA interference (RNAi), a process through which small RNAs induce sequence-specific post-transcriptional gene silencing, is widely recognized as one of the most ideal tools not only for functional genomics but also for therapeutic applications. Ensuring the specificity of small interfering RNAs (siRNAs) for specific messenger RNAs is critical, as off-target effects of siRNA can compromise the interpretation of data. Since the existence of off-target effects has been suggested in the past, it is critical to unequivocally establish that siRNAs do not present unintended effects on the biophysical properties of neurons. Here, we found that luciferase short hairpin RNA (shRNA), but not scrambled shRNA (shScr), exhibited off-target effects on voltage-gated ion channels, indicating that careful evaluation is required for studies using luciferase shRNA (shLuc) and siRNA in general.
Introduction
Since the finding of RNA interference (RNAi; Fire et al., 1998), RNAi-mediated gene knockdown has become one of the most straightforward and high throughput approaches to address gene functions from the cellular level to the animal level. Although this approach is technically straightforward, off-target effects have been repeatedly reported and represent a major concern when using RNAi gene knockdown technology. Three off-target mechanisms have been described: (1) microRNA-like regulation through sequence complementary to the small interfering RNA (siRNA) seed region (Jackson et al., 2003; Birmingham et al., 2006; Jackson et al., 2006a,b); (2) Toll-like receptor-mediated immune stimulation (Sledz et al., 2003; Kariko et al., 2004); (3) oversaturation of endogenous RNAi machinery by siRNA and short hairpin RNA (shRNA) transfection (Grimm et al., 2006; Khan et al., 2009). Therefore, it is important to perform multiple layers of confirmatory experiments to validate the effect(s) of RNAi, such as verifying phenotypes by independent multiple siRNAs, rescuing phenotypes by exogenous transgene expression and including nonspecific RNAi control(s) (Cullen, 2006).
shRNAs, the most widely used double-stranded RNAs, are processed to siRNAs by Dicer and silence target genes along the RISC-mediated RNAi pathway. shRNA directed against firefly luciferase (shLuc) has been widely used as a control in mammalian cells. To date, >70 publications used shLuc as a control. However, control shRNAs, including shLuc, have not been fully validated. Alvarez et al. (2006) reported the off-target effects of shLuc in hippocampal CA1 pyramidal neurons. Their findings indicate that transfection of shLuc caused dysregulation of spine density and dendritic complexity, and concomitant reduction of excitatory and inhibitory synaptic transmission. Despite these striking results, shLuc continues to be used as a control (Hoogenraad et al., 2010; Wakita et al., 2011; Chen et al., 2014a,b), indicating the need for a thorough validation of control shRNAs in cellular function.
Here, we have performed a detailed characterization of two control shRNAs on neuronal function and morphology, and demonstrate that shLuc has considerable off-target effects on voltage-gated ion channels without exhibiting any synaptic or morphologic defects. In contrast, nonsilencing scrambled shRNA (shScr) exhibited no abnormal neuronal functions and morphology. This study highlights the importance of thoroughly validating shRNAs and proposes shScr as a negative control appropriate for gene knockdown studies in mammalian cells.
Materials and Methods
Animals
All animal protocols were approved by the Institutional Animal Care and Use Committee of the University of Massachusetts Medical School. Male and female C57BL6 mice were used.
DNA constructs
The human H1 promoter-based pSuper Luciferase-RNAi construct has been previously described (Zhang and Macara, 2006) and targets the sequence 5´-CGTACGCGGAATACTTCGA-3´. pGIPZ-shScr (Dharmacon, #RHS4346) targets the sequence 5´-ATCTCGCTTGGGCGAGAGTAAG-3´. Dharmacon nontargeting scrambled sequence was designed using a proprietary algorithm to ensure the sequence will not target any annotated gene in human, mouse, or rat. EGFP (Clontech) gene was sub-cloned to pCAG vector.
Organotypic slice culture preparation and biolistic gene transfection
Mice hippocampal organotypic slice cultures were prepared from postnatal day 5–7 C57BL6 mice (both genders; Stoppini et al., 1991; Futai et al., 2007; Futai et al., 2013). Briefly, hippocampal slices (350-μm thickness) were prepared using a tissue chopper (Ted Pella, INC), and slices were cultured in a CO2 incubator at 35°C. Neurons were transfected at days in vitro 4–6 using a biolistic gene gun (Bio-Rad; Lo et al., 1994) with 1.6-μm gold particles (10 mg per ∼50 bullets) coated with cDNAs: shRNA vector, pCAG and pCAG-EGFP (45:45:10 μg), and were assayed 5 d after transfection, unless otherwise noted.
Electrophysiology
The recording chamber was filled with extracellular solution containing 119 mM NaCl, 2.5 mM KCl, 4 mM CaCl2, 4 mM MgCl2, 26 mM NaHCO3, 1 mM NaH2PO4, and 11 mM glucose, gassed with 5% CO2/95% O2, pH 7.4. For whole-cell recordings, thick-walled borosilicate glass pipettes (Warner Instruments) were pulled to a resistance of 2–4 MΩ. Na+ currents were measured in the presence of K+ and calcium (Ca2+) channel blockers, tetraethylammonium (TEA; 30 mM, Sigma), 4-amynopyridine (4-AP; 0.5 mM, Sigma), and CdCl2 (100 mM) in extracellular solution. To measure K+ currents, tetrodotoxin (TTX; 1 μM) and CdCl2 were applied to extracellular solution to block Na+ and Ca2+ channels.
Current-clamp recordings were performed with glass electrodes filled with internal solution containing the following: 115 mM potassium methanesulfonate, 20 mM KCl, 10 mM HEPES, 2.5 mM MgCl2, 4 mM adenosine triphosphate disodium salt, 0.4 mM guanosine triphosphate trisodium salt, 10 mM sodium phosphocreatine, and 0.6 mM EGTA, pH 7.25, with KOH. For voltage-clamp recordings, the potassium was replaced by cesium.
All experiments and the analysis of data were performed in a blind manner. Recordings were performed using a MultiClamp 700B amplifier and Digidata 1440, and data were acquired and analyzed using Clampex 10.3 and Clampfit 10.3 (Molecular Devices).
Imaging
The organotypic slices were fixed in 4% paraformaldehyde and 4% sucrose in PBS overnight. The slices were then cryoprotected in 30% sucrose in 0.1 M phosphate buffer (pH 7.4) for 2 h at room temperature followed by rapid freeze and thaw treatments. The slices were then stained with antibodies [γ-H2AX (Millipore, #05-636), GFP (MBL, #598), and secondary Alexa Fluor 647/488 dye-conjugated anti-mouse (Jackson ImmunoResearch) antibodies] in GDB buffer (0.1% gelatin, 0.5% TX-100, 450 mM NaCl, and 32% 0.1 M phosphate buffer, pH 7.4; McAllister, 2000). Two-photon and confocal microscopy were used for spine and γ-H2AX imaging, respectively. Two-photon images were captured using a LUMPlanN 60× (1.0 NA) objective. All secondary dendrites for each neuron were subjected to spine dimension and density analysis, and averaged values per dendritic segments were pooled. Scanimage software (Pologruto et al., 2003), allowed continuous acquisition of high-magnification images at 0.2-μm intervals at maximal resolution (512 × 512 pixels). Photomultiplier tube voltage settings were maintained at the same level for image collection for each cell. Registered projections of stacks collected were used to determine dendritic spine enumerations. Images of γ-H2AX in CA1 pyramidal neurons were obtained using confocal microscopy (Leica TCS SP8). Images were analyzed by MetaMorph and ImageJ software. All imaging and image analyses were performed in a blind manner.
Statistical analysis
Results are reported as mean ± SEM. The statistical significance was evaluated by one- and two-way ANOVA with post hoc Tukey for multiple comparison. Mann–Whitney U test and Student’s t test were used for two group comparison. Statistical significance was set at p < 0.05 (Table 2).
Results
Basal excitatory synaptic transmission in shRNA-transfected neurons
To examine the physiologic effects of shRNAs on neuronal function, we prepared organotypic slice cultures from mouse hippocampi and biolistically transfected GFP with pSuper empty vector (pSup), luciferase shRNA (shLuc) and shScr. To test whether control shRNAs display abnormal off-target effect(s) in excitatory synaptic function, simultaneous whole-cell recordings of untransfected and transfected CA1 pyramidal neurons were performed. AMPAR- and NMDAR-mediated evoked EPSCs were measured by stimulating Shaffer collateral inputs (Fig. 1A-C). As expected, transfection of pSuper empty vector, shLuc or shScr exhibited no off-target effects on the amplitude of AMPAR- and NMDAR-EPSCs, as well as the AMPAR/NMDAR ratio, confirming previous results (Hoogenraad et al., 2010; Chen et al., 2014b). Paired-pulse ratio (PPR) measured by the AMPAR-EPSC response obtained by the double stimulation of Shaffer collateral inputs with a 50-ms interval exhibited comparable levels of facilitation in all transfected neurons regardless of the plasmids used, indicating that the transfection of these genes do not affect presynaptic release probability (Fig. 1D).
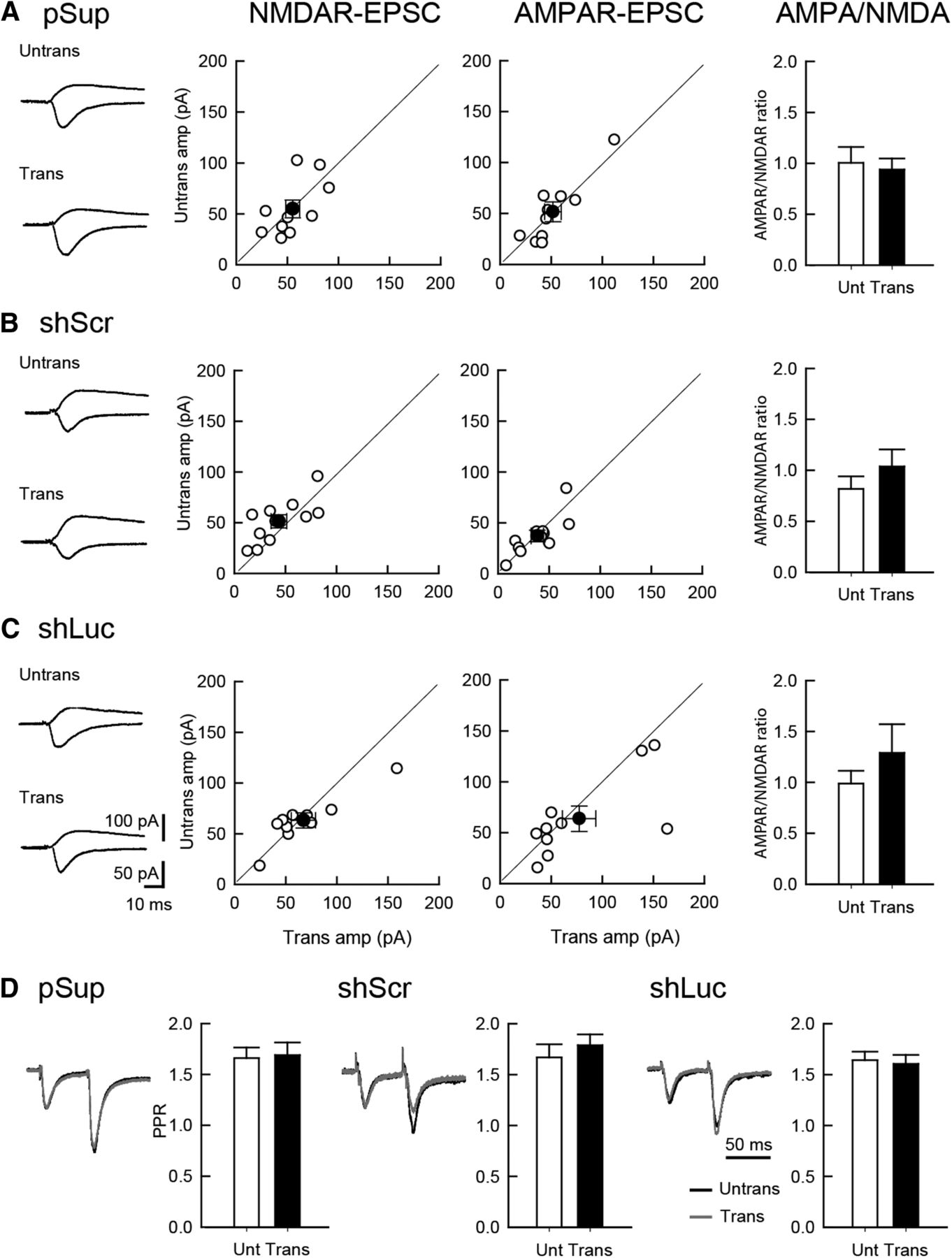
Comparable levels of excitatory synaptic transmission between shRNA-transfected and untransfected neurons. Effect of overexpression of three different plasmid transfections on excitatory synaptic transmission in hippocampal CA1 pyramidal cells. An empty vector (pSup; A), shScr (B), and shLuc (C) were transfected together with pCAG-EGFP. Recordings were conducted 5 d following transfection. Left column, Sample EPSC traces mediated by AMPARs (downward) and NMDARs (upward) from pairs of transfected neurons (Trans) and neighboring untransfected neurons (Untrans). Stimulus artifacts were truncated. Middle columns, Scattered plots of NMDAR- (left) and AMPAR- (right) EPSC amplitude (amp). Each pair of transfected and neighboring untransfected cells are presented as open symbols while filled symbols indicate the mean. Right column, Bar graphs of AMPAR/NMDAR ratios. D, PPR of AMPAR-EPSCs recorded from trans- and untransfected neurons, as indicated. Left, Sample traces. Normalized EPSCs to the first EPSC amplitude from trans- (gray) and untransfected neurons (black) are superimposed. Right, Summary graphs of PPR. The PPR was calculated by dividing the average amplitude of the second EPSC by that of the first EPSC. Number of cell pairs tested: pSup, 10 cells/6 mice; shLuc, 10/6; shScr, 11/6 (for AMPAR-EPSC, NMDAR-EPSC, and PPR, respectively).
Normal inhibitory synaptic transmission in shRNA-transfected neurons
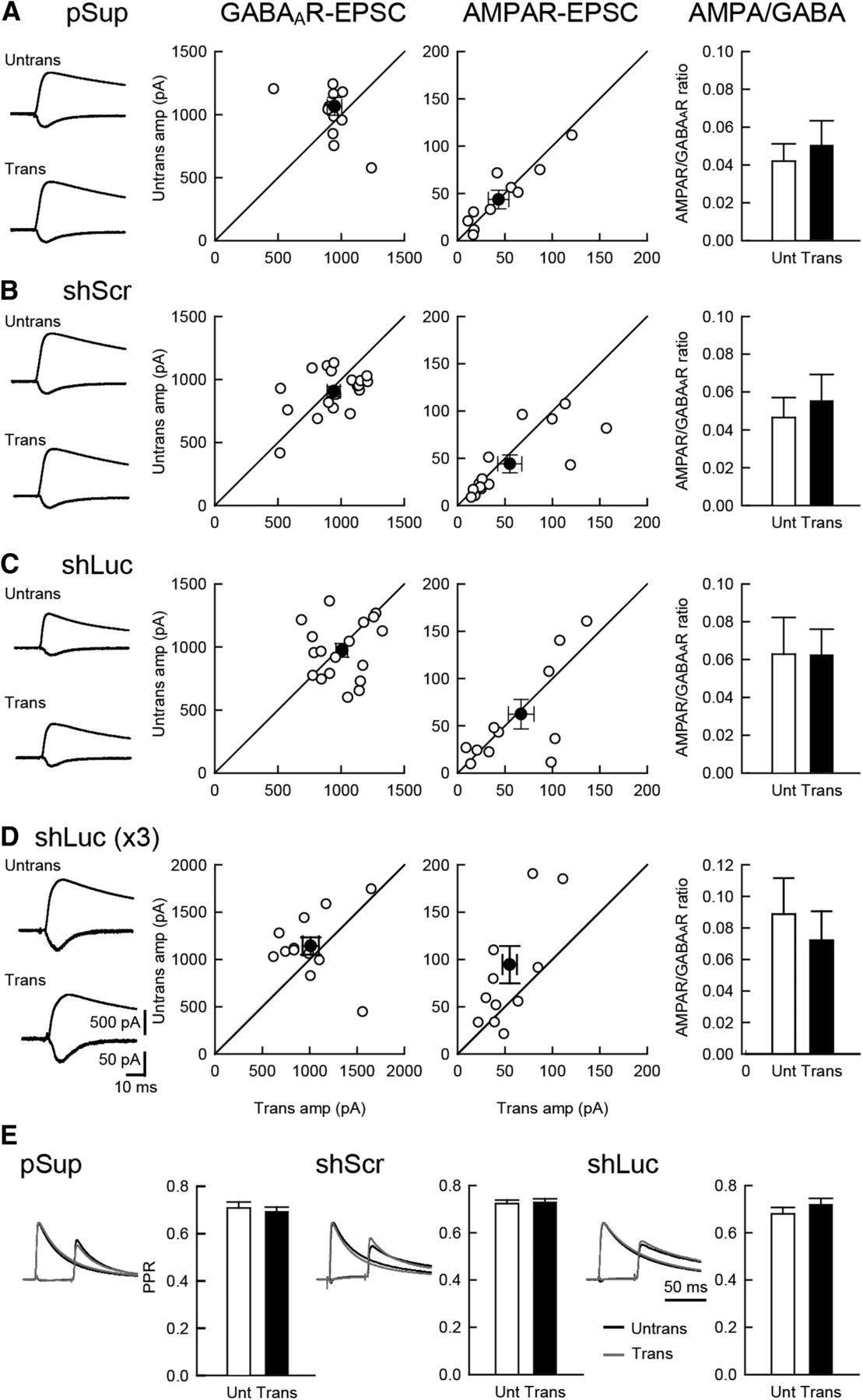
Next, we examined the effects of shRNAs on inhibitory synaptic transmission and the balance of excitatory and inhibitory transmission. Neither GABAAR-mediated IPSC nor the ratio of AMPAR and GABAAR responses displayed significant changes following transfection of any of the three plasmids (Fig. 2A–C). Paired-pulse stimulation of inhibitory inputs exhibited comparable levels of depression in all transfected neurons, suggesting that the transfection of these genes do not have any effects on presynaptic release probability (Fig. 2E).

Comparable level of inhibitory synaptic transmission between shRNA-transfected and untransfected neurons. Effect of overexpression of the three different plasmid on excitatory and inhibitory synaptic transmission in hippocampal CA1 pyramidal cells. An empty vector (pSup; A), shScr (B), shLuc (C) and shLuc x3 (triple the amount of shLuc) (D) were transfected together with pCAG-EGFP. Left column, Sample AMPAR-EPSC and GABAAR-IPSC traces mediated by AMPARs (downward) and GABAARs (upward) from pairs of transfected neurons (Trans) and neighboring untransfected neurons (Untrans). Stimulus artifacts were truncated. Middle columns, Scattered plots of GABAAR- (left) and AMPAR- (right) EPSC amplitude (amp). Each pair of transfected and neighboring untransfected cells are presented as open symbols while filled symbols indicate the mean. Right column, Bar graphs of AMPAR/GABAAR ratios. E, PPR of GABAAR-IPSCs recorded from trans- and untransfected neurons, as indicated. Left, Sample traces. Normalized IPSCs to the first IPSC amplitude from trans- (gray) and untransfected neurons (black) were superimposed. The First GABAAR-IPSC overlaps with the second IPSC. Therefore, the first EPSC was cancelled by subtracting the traces receiving a single pulse from those receiving a paired pulse, both normalized to the first response. Right, Summary graphs of PPR. The PPR was calculated by dividing the average amplitude of the second IPSC by that of the first IPSC. Number of cell pairs: pSup, 13, 11, and 13 cells/7 mice (13, 11, 13/7); shLuc (18, 13, 18/8); shScr (20, 15, 20/8); shLuc x3 (13, 12/6), for GABAAR-IPSC, AMPAR-EPSC, and PPR, respectively.
In contrast to our results, Alvarez et al. (2006) reported that shLuc displayed reduced inhibitory and excitatory synaptic transmission in organotypic hippocampal CA1 neurons. The dosage of shRNAs, reported to contribute to off-target effects (Jackson et al., 2006a), may account for this discrepancy as Alvarez et al. (2006) used a 1.3-fold larger amount of shRNA than used in the present study. To test this possibility, we increased the shLuc plasmid concentration three-fold and measured IPSCs and AMPAR-EPSCs (Fig. 2D). A three-fold increase of shLuc did not significantly reduce synaptic transmission, indicating that a potential dosage-dependent off-target effect of shLuc is small in synaptic function as measured here and most likely is not the primary reason for the differences in results between those reported here and by Alvarez et al. (2006).
Normal dendritic morphology in shRNA-transfected neurons
We next addressed the effects of shRNAs on neuronal morphology. We overexpressed shRNAs together with EGFP. The transfected neurons were immunostained against GFP followed by imaging using two-photon microscopy (Fig. 3). The dendritic morphology, including spine density and dimension, was found to be unchanged by overexpression of the two shRNAs. Overall, we did not identify off-target effects of control shRNAs in synaptic function and structure.
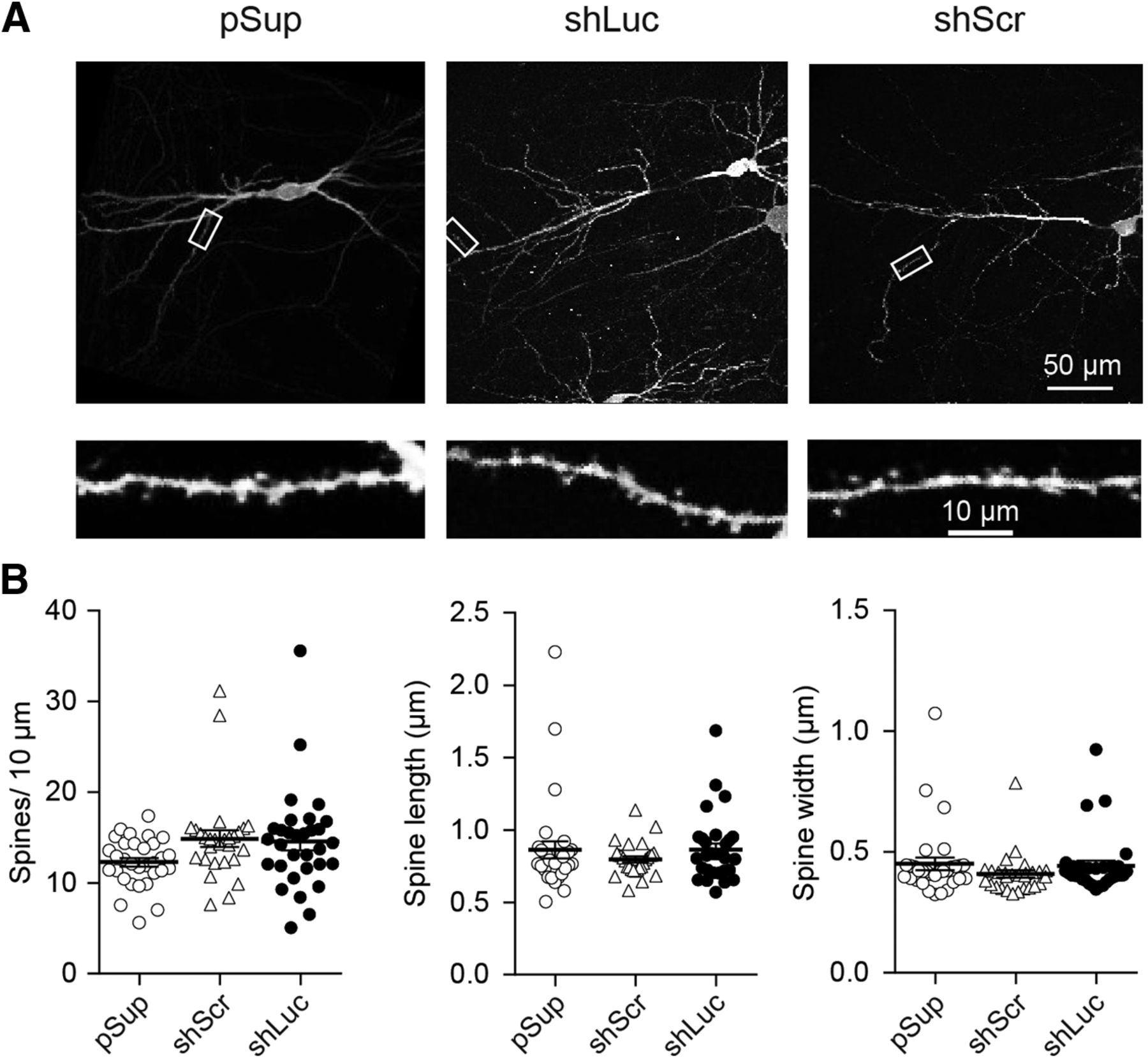
Comparable level of dendritic structure between different shRNA-transfected neurons. Effect of overexpression of three different plasmid transfections on dendritic morphology in hippocampal CA1 pyramidal neurons. A, Low (top) and high (bottom) magnification images obtained from empty vector, pSup (left), shLuc (middle), and shScr (right) transfected neurons. Each plasmid was cotransfected with pCAG-EGFP. Fixed slices were immunostained against GFP and neuronal images were obtained by two-photon microscopy. B, Scatter plots of spine length (left), width (middle), and density (right). Error bars indicate SEMs. Note that none of these parameters displayed statistical significance by gene transfection. Number of dendritic segments/cells/mice: pSup, 31/5/5; shLuc, 31/5/5; and shScr, 28/5/5.
Reduced membrane excitability and voltage-gated ion channel function in shLuc-transfected CA1 pyramidal neurons
To examine the effects of shRNAs on membrane properties, neurons transfected with shRNAs and EGFP were compared to untransfected neurons with respect to action potentials (APs) and basic membrane properties (Fig. 4; Table 1). To our great surprise, CA1 neurons transfected with shLuc exhibited a reduced number of APs compared to untransfected control neurons (Fig. 4A). The transfection of pSup did not cause any abnormal excitability, indicating that this vector backbone is not the cause of the reduced excitability in shLuc-transfected neurons.

Reduced membrane excitabilities in shLuc-transfected CA1 pyramidal neurons. A, Effect of shRNA overexpression on neuronal excitability. An empty vector (pSup), shLuc, or shScr was transfected together with pCAG-EGFP. Top, Sample traces from untransfected and transfected CA1 pyramidal neurons in organotypic hippocampal slice cultures. The superimposed traces were elicited by current injections of 0, 100, and 500 pA for 200 ms. Bottom, Summary graph of the frequency of APs in untransfected and transfected neurons. The input–output relationship [number of spikes elicited versus amount of current injection (200-ms duration)] was plotted for untransfected and transfected neurons. Neurons were held at resting membrane potentials (−56.8 mV ± 0.57, n = 100 cells, 9 mice). Number of cells tested: untrans, 45 cells from eight mice (45/8); pSup, 13/7; shScr, 20/5; and shLuc, 22/9. B, Effect of shRNA overexpression on AP kinetics. Top, Sample traces from CA1 pyramidal neurons untransfected and transfected with the three different plasmids. Single APs were induced by current injection (100 pA for 4 ms) and threshold (horizontal arrows) and half width of AP determined by vertical double arrow heads in trans- and untransfected neurons were measured. Bottom, Summary graph of the threshold (left) and half width (right) of single AP in untransfected and transfected neurons. Number of cells tested: untrans, 45 cells/8 mice (45/8); pSup, 13/7; shScr, 20/5; shLuc, 22/9. C, top, Sample traces of sodium currents recorded from untransfected and transfected CA1 pyramidal neurons. Note that these currents were completely blocked by TTX (right). Bottom, Summary graph of the sodium currents in untransfected and transfected neurons. Neurons were voltage-clamped at –80 mV and depolarized from –80 to 30 mV (10-ms duration). Number of cells: untrans, 16 cells/5 mice; pSup, 10/5; shScr, 10/5; shLuc, 14/6. D, top, Sample traces of potassium currents recorded from untransfected and transfected CA1 pyramidal neurons. Note that these upward currents were completely blocked by TEA and 4-AP (right). Bottom, Summary graph of the potassium currents in untransfected and transfected neurons. Neurons were voltage-clamped at –80 mV and depolarized from –80 to 30 mV (10-ms duration). Number of cells tested: untrans, 15 cells/4 mice; pSup, 13/3; shScr, 7/3; shLuc, 9/3.
The basic membrane properties of untransfected and gene transfected hippocampal CA1 neurons
Statistical table
To further test whether shLuc changed the threshold and kinetics of APs, we compared the onset, threshold, amplitude and half-width of APs in untransfected and transfected neurons (Fig. 4B; Table 1). We found that APs in shLuc-transfected neurons exhibited increased half-width (Fig. 4B, lower right) and decrease of threshold (Fig. 4B, lower left) without changing the amplitude of AP (Table 1). The basic membrane properties, including resting membrane potential and series and input resistance, remained unchanged, indicating an off-target effect of shLuc on voltage-gated ion channel function (Table 1). Taken together, our results strongly suggest that shLuc has off-target effect(s) on membrane excitability and AP kinetics in CA1 pyramidal neurons.
Given these striking results, we next addressed the underlying mechanism of the off-target effect of shLuc on membrane excitability and APs. Whole-cell voltage clamp recordings were performed in CA1 pyramidal neurons and the current−voltage (I-V) relationship of TTX-sensitive sodium (Na+) channel current was measured. Importantly, Na+ current maximum amplitude between −20 and +30 mV was markedly reduced in shLuc-transfected compared to untransfected neurons (Fig. 4C, closed circles). We then tested whether voltage-dependent potassium (K+) currents were altered by shLuc transfection. The potassium channel currents, sensitive to the broad potassium channel blockers, TEA, and 4-AP, were also markedly reduced in shLuc-transfected neurons compared with other neurons (Fig. 4D, closed circle). These results indicate that shLuc has off-target effects on voltage-gated sodium and potassium channels.
Normal H2A-X phosphorylation in shRNA-transfected neurons
Lastly, we addressed whether the reduced membrane excitability in shLuc-transfected neurons is due to potential cellular stress or damage to cellular functions. To test this hypothesis, we examined the level of the phosphorylation of histone H2A.X. H2A.X phosphorylation at serine 139 (γ-H2AX) is a routinely used biomarker to detect DNA damage and DNA replication stress (Mah et al., 2010). γ-H2AX is greatly increased in neurons by seizure insult (Crowe et al., 2011) and camptothecin, an inhibitor of the DNA enzyme topoisomerase I (Mah et al., 2010; Fig. 5A). Importantly, the level of γ-H2AX in shLuc-transfected neurons was comparable to that of pSup- and shScr-transfected neurons, suggesting that transfection of shLuc does not alter the overall health of the neurons (Fig. 5B). In summary, we conclude that shLuc exhibits off-target effects on membrane excitability without changing synaptic transmission, dendritic morphology or cellular health.
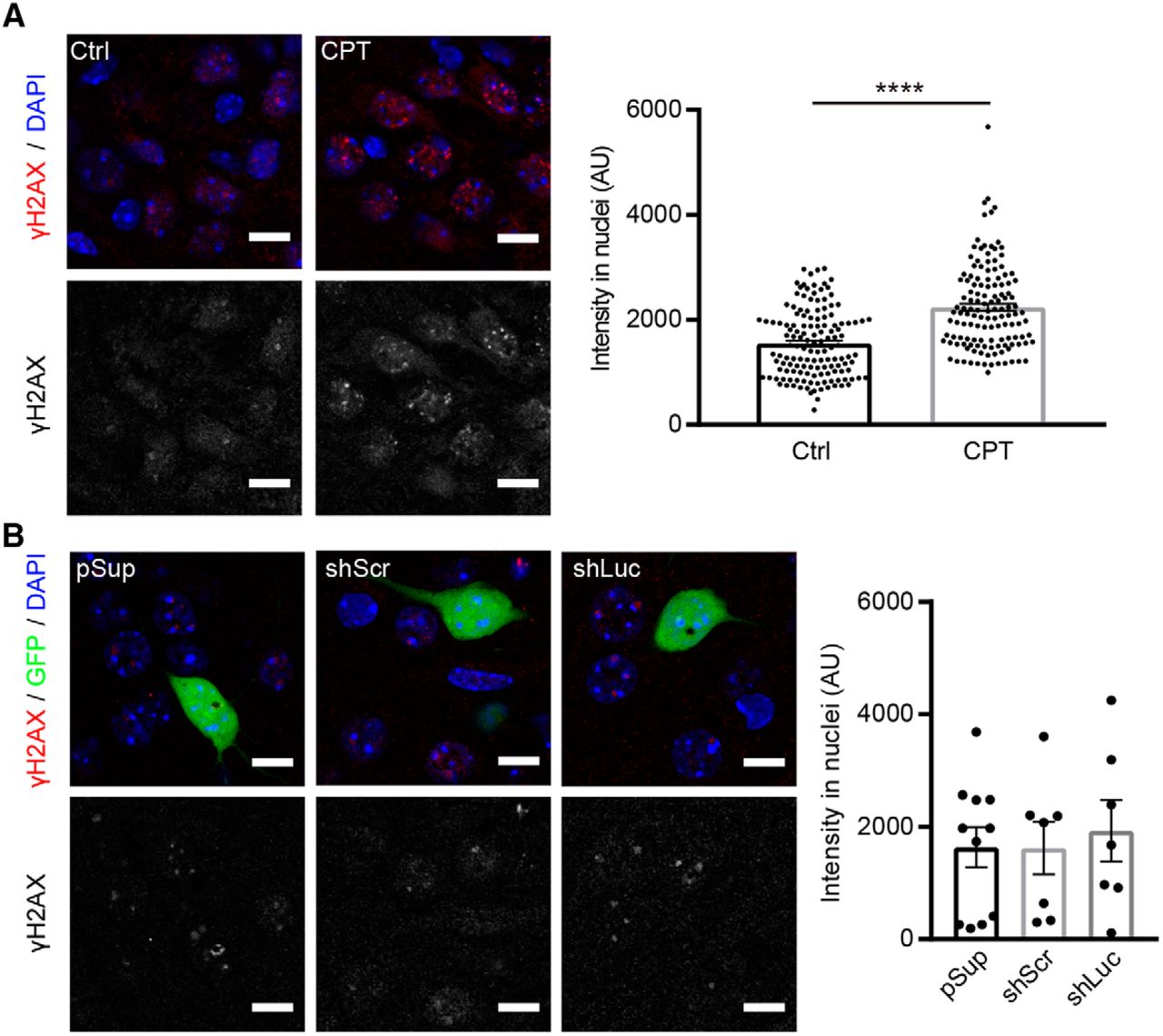
Comparable levels of H2A-X phosphorylation in neurons transfected with different shRNA vectors. γ-H2AX immunoreactivity in shRNA-transfected hippocampal CA1 neurons. A, left, Representative confocal images of immunofluorescence staining against γ-H2AX (red) and DAPI (blue) in CA1 pyramidal neurons. Right, Quantification of γ-H2AX signal intensity in nuclei. Treatment with 10 μM camptothecin (CPT) for 6 h increased the signal of γ-H2AX in nuclei of neurons in organotypic slice cultures, confirming the specificity of the anti-γ-H2AX antibody. AU, arbitrary units. Number of cells/mice: mock control (Ctrl), 140/3; CPT, 133/3. B, left, Representative confocal images of immunofluorescence staining against γ-H2AX (red), GFP epifluorescence (green) and DAPI (blue) in pSup-, shLuc-, and shScr-transfected neurons. Right, Quantification of γ-H2AX signal intensity in nuclei. All transfected neurons exhibited comparable levels of γ-H2AX signal. Number of cells/mice: pSup, 11/3; shScr, 7/4; shLuc, 7/3. Scale bars: 10 μm.
Discussion
RNAi is a powerful approach to validate the function of target genes both in vitro and in vivo. In light of repeated indications that shRNAs may have off-target effects (Cullen, 2006), their effects should be carefully studied. Here, we found that shLuc transfection increased AP threshold and half-width in addition to reducing the number of APs. shLuc also reduced TTX-sensitive sodium and TEA- and 4AP-sensitive potassium currents that contribute to shaping APs. In contrast to its effects on these currents, shLuc failed to alter basic membrane properties, suggesting that shLuc does not exhibit off-target effects on pumps (e.g., Na+/K+-ATPase) and other channels (e.g., leak channels) that contribute to maintaining resting membrane potential.
Is there any possibilities that shLuc targets the genes regulating the function of voltage-gated ion channels? A BLAST search against shLuc guide sequence (19 base-pairs: TCGAAGTATTCCGCGTACG), the strand incorporated into RNA-induced silencing complex, displayed seven genes with low homology (11–12 bp match, 58–63% homology), including Dnah11 (dynein axonemal heavy chain 11, XM_017314949.1), Plpp5 (phospholipid phosphatase 5, NM_001293703.1), Lpcat2 (lysophosphatidylcholine acyltransferase 2, XM_006531052.2), Asb7 (Ankyrin repeat and SOCS box-containing 7, XM_006540568.3), Ipo8 (importin 8, XR_001785142.1), Dis3 (DIS3 homolog exosome endoribonuclease and 3´->5´ exoribonuclease, XM_006519602.3), Fbcl20 (F-box and leucine-rich repeat protein 20, XM_006534299.3). Based on the homology, it is difficult to consider these genes as the potential target of shLuc and none of these have been known as the regulators of Na+ and K+ channels.
Alvarez et al. (2006) reported that shLuc caused a lack of dendritic spines and reduced excitatory and inhibitory synaptic transmission in hippocampal pyramidal neurons. However, we were unable to confirm their observations. As described in our results, neurons transfected with shLuc display normal synaptic transmission and dendritic spine structure compared with untransfected and other genes transfected neurons. Our results are also supported by other studies demonstrating normal excitatory synaptic transmission, long-term potentiation and dendritic morphology in shLuc-transfected neurons (Hoogenraad et al., 2010; Wakita et al., 2011; Chen et al., 2014a,b). Since Alvarez et al. (2006) used the same experimental approach as ours (Stoppini et al., 1991), this discrepancy may be due to differences in precise details of the experimental procedures, including the vector backbone and the methods of gene transfection in organotypic slice cultures. For example, Alvarez et al. (2006) used U6 promoter to synthesize shLuc but pSuper vector consists with H1 promoter. Both H1 and U6 promoters are member of the type III class of Pol III RNA promoters, but it is possible that these two different promoters cause the distinct off-target effects in neurons (Mäkinen et al., 2006). However, regardless of this diverging result, it is clear that shLuc exhibits off-target effects on ion channel or synaptic function, discouraging the usage of shLuc as a control shRNA, at least in hippocampal CA1 pyramidal cells.
While comprehensive guidelines for the RNAi approach have been described (2003), detailed studies of off-target effects of control shRNAs are very limited. Thus, our comparative study on shRNAs in pyramidal neurons is an important consideration for future RNAi experiments. Of note, gene expression is differentially regulated by cell-type, cellular activity and development. Therefore, careful evaluation is required for shRNA constructs, including nonsilencing controls. However, we consider that shScr is currently the best validated control for the RNAi approach in mammalian cells including neurons.
Acknowledgments
Acknowledgements: We thank Dr. Paul D. Gardner for valuable discussions and Mrs. Naoe Watanabe for skillful technical assistance.
Footnotes
The authors declare no competing financial interests.
This work was supported by grants from the Whitehall Foundation (K.F.), Japan Foundation for Pediatric Research (K.F.), and National Institutes of Health Grants R01NS085215 (to K.F.) and 1R01AA020501 (to G.E.M.).
This is an open-access article distributed under the terms of the Creative Commons Attribution 4.0 International license, which permits unrestricted use, distribution and reproduction in any medium provided that the original work is properly attributed.
References
In this issue

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}



